AtCS25基因过表达载体的构建和拟南芥的遗传转化
2012-06-14刘博宇
刘博宇,阮 颖
(湖南农业大学生物科学技术学院,长沙410128)
拟南芥作为经典的模式植物,是第一个完成全基因组测序的高等植物,随着研究的不断深入,越来越多的基因的功能被人们所了解,但是仍然有大量的功能未知的基因等待人们去探究。随着生物信息学、反向遗传学和现代分子生物学技术的不断进步,这些功能未知基因的定位,以及对于它们的表达与调控的研究,引起了人们极大的兴趣。
在拟南芥大量功能未知的基因里,有很多与花粉和花粉管的发育相关。在花粉发育过程中,涉及近万种特异表达的基因,但是迄今所发现的特异基因也不过几十种。花粉发育过程中的绝大多数基因的表达具有时间和空间的特异性,早期基因一般在减数分裂后被激活;而晚期基因一般在小孢子有丝分裂后表达,另外还有一些基因在植物生长发育过程中持续表达。近几年来,人们在拟南芥中分离了一些花粉发育特异基因,如花药绒毡层特异表达基因 A9,AtMLH1,AtDMC1,SYN1,DIF1,在花粉管的伸长期表达的AFH1,在小孢子有丝分裂前表达的AMS和在成熟花粉中特异表达的AVP1,以及一些在花粉发育晚期表达的基因[1]。
在与花粉发育相关的基因中,有一类编码花粉过敏原蛋白(Pollen allergen)。该蛋白不仅与植物花粉的发育有关,在植物整个生长发育过程中也起着重要的作用。花粉过敏原的分子大小从8 kD至50 kD不等,他们的特点表现为可溶,并且较稳定。到今天为止,已经有报道的花粉过敏原家族有10余类[2]。AtCS25属于花粉过敏原家族。
Profilin,BetvⅠ和BetvⅢ是白桦花粉的3种主要过敏原。Profilin的转录产物为800 nt,推测蛋白为266 aa,该蛋白不仅在花粉中有表达,同时在植物其他器官中也有微量的表达。BetvⅠ编码一个长度为160 aa蛋白质,而BetvⅢ包含203个氨基酸,它们都属于Ca2+结合蛋白,特异表达于成熟花粉中,并参与花粉管的萌发和生长[3,4]。豚草花粉的主要过敏原是AmbaI,其分子量大小为38 kD,在其N端存在一个疏水区信号肽序列,是一种可溶性蛋白。AmbaI基因家族的同源性非常高,转录的mRNA都在1 500 nt左右[3]。研究人员从黑麦草(ryegrass)花粉cDNA文库中筛选得到了两个糖蛋白过敏原基因,分别为LolpIa和LolpIb。LolpIa基因在花粉细胞中表达,编码的蛋白质大小为35 kD,其N端23个氨基酸为信号肽。LolpIb所编码的蛋白质大小为31 kD,其主要存在于淀粉粒中,信息学分析表明其N端25个氨基酸与叶绿体转运肽相似[3]。
Ole_e_1是第一个从橄榄树花粉中分离并纯化的花粉过敏原蛋白[5]。该蛋白由一条含有145个氨基酸的单肽链构成,它与番茄中LAT52基因和玉米花粉中Zmc13基因编码的蛋白具有相似性[6]。Ole结构域包含6个保守的半胱氨酸,这可能与二硫键的形成有关。同时该结构域还包含有一致的序列E/Q/T-G-X-V-Y-C-D-T/N/P-C-R(X代表任何氨基酸)[2]。有一些富含脯氨酸并且含有Ole结构域的蛋白编码类伸展蛋白[7]。至今已从橄榄树花粉中分离并证实了10种含有Ole结构域的蛋白。人们在水稻中的研究表明,编码含有Ole结构域的蛋白的基因在水稻的各个部位都有表达,说明花粉过敏原基因不仅在植物生殖生长中起作用,在植物的营养生长中也起到了很重要的作用[2]。
尽管对花粉过敏原蛋白的研究已取得一定的进展,但是在拟南芥中对该类蛋白的了解还很少,尤其是它们的功能。本文对拟南芥AtCS25基因做了生物信息学分析,以及过表达分析,以为继续深入探究该基因的功能奠定基础。
1 材料与方法
1.1 植物材料
拟南芥(Arabidopsis thaliana)哥伦比亚生态型Col,由湖南农业大学植物发育与表观遗传调控实验室保存。
1.2 菌株和质粒
大肠杆菌 (Escherichia coli)DH5α,农杆菌GV3101和pBI121过表达载体由湖南农业大学植物发育与表观遗传调控实验室保存;pMD-19购自TAKARA公司。
1.3 方法
1.3.1 生物信息学分析
在TAIR上下载AtCS25基因的全长CDS序列,利用DNA Star软件将其翻译成蛋白序列,然后将翻译后的蛋白序列输入到NCBI上进行保守结构域分析。利用在线跨膜结构分析软件TMHMM Server v 2.0分析该蛋白是否存在跨膜结构,再通过在线亚细胞定位分析软件TargetP 1.1 serve来推测该蛋白在细胞内的定位。
1.3.2 引物设计以及AtCS25基因全长CDS片段的克隆
由于园林花卉相对于人工培育花卉具有更强的生命力,因此在我国的园林绿化中应用非常广泛。园林绿化使用花卉时,要考虑各方面因素,选择适宜的花卉种类,并采取科学合理的栽培方法,加强后期养护管理,提高花卉成活率。
根据网站 TAIR上公布的序列,采用 Primer Premier 5引物设计软件设计了一对引物AtCS25-F/R,用来扩增AtCS25基因的全长CDS。并根据载体pBI121的特点,在引物的5'端分别加上Xbal和BamH1位点,引物序列为:
AtCS25-F:5'-TCTAGA ATGGCATCCACAG GCGCCG -3'(Xbal),
AtCS25-R:5'-GGATCCCTAGTAAACGGGT TTGGGAGCC-3'(BamH1)
扩增产物大小为567 bp。以拟南芥Col生态型为材料,采用Trizol法提取拟南芥叶片总RNA。根据Fermentas公司的RevertAidTMFirst Strand cDNA Synthesis Kit试剂盒,将得到的 RNA反转录为cDNA。以拟南芥Col生态型叶片cDNA为模板,用AtCS25-F/R引物扩增AtCS25基因的全长CDS。PCR反应采用NEB公司的LongAmp Taq高保真酶进行扩增,退火温度为60℃,循环28次。将PCR产物连接到 pMD19-T载体上,构成 pMD19-T-AtCS25载体,通过热激法将此载体转化入大肠杆菌DH5α中,挑选阳性克隆送博尚生物公司测序。
1.3.3 AtCS25基因表达载体的构建
用Xbal和BamH1限制性内切酶分别对pMD19-T-AtCS25和pBI121载体进行双酶切,通过胶回收试剂盒将目的片段和载体片段分别回收,采用 Fermentas公司的快速 T4连接试剂盒,将AtCS25基因全长CDS连接至pBI121载体相对应的位置,构建成pBI121-AtCS25过表达载体。
通过冻融法将构建好的AtCS25基因过表达载体pBI121-AtCS25转化到根癌农杆菌GV3101中,挑取阳性克隆后进行PCR验证。采用浸花法将pBI121-AtCS25载体转化拟南芥Col生态型,并获得转基因植株。
1.3.5 拟南芥转化子的筛选和鉴定
将收获的T0代种子铺在含有卡那霉素的MS培养基上,卡那霉素浓度为40 mg/mL,待植株生长15 d后,将具有抗性的植株从MS培养基上移栽至土壤中。为了检测目的片段是否已经整合到转化子的基因组上,根据pBI121-AtCS25载体序列信息,设计检测引物121JC-F:5'-TTCGCAAGACCCTTC CTCTATATAA -3',121JC -R:5'-AGCTCGACCAG GATGGGCAC-3'。提取抗性植株的总 DNA,用该对引物进行PCR检测,鉴定转化子。
2 结果与分析
2.1 生物信息学分析结果
将AtCS25氨基酸序列输入到NCBI网站中分析,结果表明,在其第57至151位氨基酸构成一个Pollen_Ole_e_1(Ole)结构域(图1)。

图1 结构域分析Fig.1 Domain analysis
用在线分析软件TMHMM 2.0分析AtCS25氨基酸序列,结果显示在第7至26位氨基酸处存在一个跨膜结构(图2)。

图2 跨膜结构分析Fig.2 Transmembrane structure analysis
使用亚细胞定位分析软件TargetP1.1对AtCS25氨基酸序列进行分析表明,该蛋白很有可能是一种分泌蛋白,可能定位在内质网上(图3)。

图3 亚细胞定位Fig.3 Subcellular localization
2.2 目的片段的克隆与分析
通过梯度PCR确定了引物AtCS25-F/R最适退火温度为 61℃。PCR扩增 AtCS25基因全长CDS,得到了约567 bp大小的条带(图4),其大小与预期的结果一致。将得到的目的片段切胶回收,然后连接到pMD19-T载体上,通过热激法将连接后的载体转化到大肠杆菌DH5α中,选取阳性克隆送公司测序,将测序结果与网站上公布的序列进行比对,其同源性为100%。表明克隆结果正确,可以用于下一步实验。

图4 AtCS25基因的克隆Fig.4 Clone of AtCS25 gene
2.3 重组质粒的酶切验证
挑取转化菌落,提取重组质粒 pBI121-AtCS25,用 Xbal和 BamH1双酶切 pBI121-AtCS25,得到一条约567 bp的条带(图5),表明pBI121-AtCS25载体已经构建成功。

图5 重组质粒的酶切验证Fig.5 Identification of recombinant vector by restriction enzyme digestion
2.4 转基因拟南芥的获得和鉴定
一共从含有卡那霉素的MS培养基上筛选出抗性苗5株,用CTAB法提取转基因植株和未经转化的Col野生型叶片的总 DNA,用载体检测引物121JC-F/121JC-R对其进行扩增,结果如图6。电泳结果显示,Col野生型没有出现条带,1~5号泳道条带大小与7号泳道质粒对照扩增出的片段一致,证明获得了5株转基因苗。

图6 抗性植株的PCR检测Fig.6 PCR identification of transgenic plants with kanamycin-resistance1~5.抗性苗;6.阴性对照;7.阳性对照
2.5 转基因植株过表达分析
提取Col野生型和抗性植株的总RNA,反转录后得到 cDNA,用 AtCS25基因的表达检测扩增cDNA,结果如图7所示。AtCS25基因在Col野生型中的表达量明显低于其他5株转基因植株,表明AtCS25基因在CaMV35S强启动子的驱动下得到了过表达。转基因植株在个体上比野生型略微大一些,茎秆稍粗,花在形态上也比野生型略大,说明该基因不仅与花粉的发育相关,还可能在植株整个发育过程中起作用。
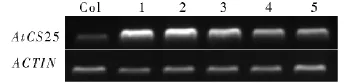
图7 AtCS25基因表达水平的RT-PCR分析Fig.7 RT-PCR analysis of AtCS25 overexpression
3 小结与讨论
对编码包含Ole结构域的基因的研究最初是在番茄和玉米中进行的[9,10],而在橄榄树花粉中分离得到的一些花粉过敏原蛋白,被认为是主要的过敏原物质。到目前为止,在不同的植物中共鉴定出数百个编码Ole结构域的基因。通过生物信息学分析,以及分子生物学的方法,研究人员在水稻中鉴定了114个花粉过敏原基因[2],这说明借助生物信息学以及公共的数据库资源来推测并鉴定新基因的方式变得尤为重要。尽管如此,只有少数基因的功能得到了验证,包括花粉特异表达基因Zmc13和在雌蕊中特异表达的基因 LAT52[7]。
本研究通过生物信息学分析,以及基因过表达的方法,对拟南芥中包含Ole结构域的功能未知基因进行了初步的探究。结果表明,该基因编码的蛋白属于花粉过敏原蛋白家族,具有跨膜结构,并很可能为一种分泌蛋白,推测该基因不仅与植物花粉的发育有关,也可能在植物营养生长过程中起着重要的作用。过表达植株与野生型植株表型相似,该基因的过量表达对植株生长发育具体有哪些影响有待进一步的研究。
[1]刘乐承,张 弢,曹家树.植物花粉发育的分子生物学研究进展[J].长江大学学报,2006,3(3):174-177.
[2]Jiang SY,Jasmin XH,Ting YY,et al.Genome- wide identication and molecular characterization of Ole_e_I,Allerg_1 and Allerg_2 domain-containing pollen-allergen- like genesin Oryza sativa[J].DNA Research,2005,12:167 –179.
[3]Mascarenhas JP.Pollen gene expression:molecular evidence[J].International Review of Cytology,1992,140:3-17.
[4]Susanne Seiberler,Otto Scheiner,Dietrich Kraft,et al.Characterization of a birch pollen allergen,Bet v Ill,representing a novel class of Ca2+binding proteins;specific expression in mature pollen and dependence of patients’IgE binding on protein - bound Ca2+[J].EMBO J,1994,13:3481- 3486.
[5]Marsh DG,Goodfriend L,King TP,et al.Allergen nomenclature[J].Bull World Health Organ,1986,64:767–774.
[6]De Dios Alche J,M’rani-Alaoui M,Castro AJ,et al.Ole e 1,the major allergen from Olive(Olea europaea L.)pollen,increases its expression and is released to the culture medium during in vitro germination[J].Plant Cell Physiol,2004,45:1149 –1157.
[7]Goldman MH,Pezzotti M,Seurinck J,et al.Developmental expression of tobacco pistil-specific genes encoding novel extensin - like proteins[J].Plant Cell,1992,4:1041–1051.
[8]宋江华,张立新.植物花粉发育的基因工程研究进展[J].农业生物技术科学,2008,24(12):23–26.
[9]Hanson DD,Hamilton DA,Travis JL,et al.Characterization of a pollen-specific cDNA clone from Zea mays and its expression[J].Plant Cell,1989,1:173 –179.
[10]Muschietti J,Dircks L,Vancanneyt G,et al.LAT52 protein is essential for tomato pollen development:pollen expressing antisense LAT52 RNA hydrates and germinates abnormally and cannot achieve fertilization[J].Plant J,1994,6:321–338.
