成虫日龄与交配状态对棉褐环野螟繁殖力的影响
2012-02-28张清泉张雪丽
张清泉, 张雪丽, 陆 温
(1.广西壮族自治区植保总站,南宁 530022; 2.广西大学农学院,南宁 530004)
棉褐环野螟[Haritalodes der ogata (Fabricius)]属鳞翅目,草螟科(Crambidae)。别名棉大卷叶螟、棉大卷叶虫、包叶虫、棉野螟蛾、棉卷叶野螟[1]。这是绿篱植物——朱槿(Hibiscus rosa-sinensis)上的一种重要害虫,同时也是农作物棉花的主要害虫,在我国分布广泛。该虫以老熟幼虫在地上或草丛中结薄茧越冬,翌年4月下旬开始出现第1代幼虫,11月下旬部分老熟幼虫开始越冬,部分继续发育为下代。其以幼虫吐丝缀叶为害,盛发时整株布满虫苞,加害后成扫帚丝状,严重的吃光全部叶片,既影响植株生长,又不堪观赏[2]。
近年来棉褐环野螟除为害朱槿外,还为害木棉(Bombax mal abaricu m)、蜀葵(Althaea r osea),由于其生活史相对较短,产卵量大且繁殖快,部分地区绿化带已为害成灾,常规杀虫剂不仅防效差,而且还会造成棉褐环野螟再猖獗并引发出一系列的环境问题。因此,探索或开发棉褐环野螟的其他有效防治途径和方法,已成了控制棉褐环野螟成灾的关键所在。目前,在诸多的非化学农药防治途径中,应用性信息素防治鳞翅目害虫已取得了很大的进展[3-6],并有望于减少或取代化学杀虫剂的应用。搞清楚影响棉褐环野螟产卵量、孵化率及成虫寿命的因素,可对性信息素的研究开发提供重要的理论依据,为此,作者在室内对棉褐环野螟的繁殖进行了较为系统的研究,主要结果如下。
1 材料与方法
1.1 供试虫源
棉褐环野螟的初始虫源在广西大学校园朱槿绿化区采集。新孵幼虫用新采摘的朱槿叶片饲养,每天更换食料。到2龄后,将幼虫转移到饲养瓶内(直径10 c m,高12 c m)用新鲜朱槿叶片饲养,每2 d换一次新鲜叶片,每瓶2~3头,直至幼虫成熟。幼虫老熟后,让其在剩下的朱槿叶片中化蛹。将蛹分瓶饲养。成虫羽化后,将其转移到透明塑料饲养瓶(直径16 c m,高23 c m)中配对饲养。饲养瓶顶端用吸水纸和纱网封口,内放保鲜朱槿枝条供成虫产卵和栖息。成虫用5%的蜜糖水饲养并每天更换。饲养温度为(25±2)℃,光周期为L∥D=14 h∥10 h(光照时间为06:00-20:00)。供试昆虫及相关试验均是在该条件下获得和完成的。
1.2 雌虫交配持续时长对繁殖的影响
取羽化后2 d的处女雌蛾和雄蛾,分别设置交配时长20、40、80、120、180 min与成虫自然交配(不进行人为干预分开,不计交配时间长短)1次,共6个处理。将雌雄成虫各一头配对,如果不交配将在试验中被剔除,如果能进入正常交配,则分别在预定的交配时间内人为将其分开,中断交配,雌虫分别单独在大饲养笼中以5%蜜糖水饲养,放入盆栽植株供其产卵,每天更换植株并检查记录产卵量,已产卵植株放入饲养室待查并记录其孵化率;最后统计总产卵量和孵化率;每处理设20次重复。
1.3 雌虫交配次数对繁殖的影响
选取体重(蛹重)相当的处女雄蛾和处女雌蛾,分别设置雌虫不交配、雌虫交配1次、交配2次和长期配对等4种处理。不交配雌虫处理:自雌虫羽化之日起便单独饲养并放入盆栽朱槿植株供其产卵,观察记录产卵量、孵化率和成虫寿命;交配1次的处理:雌虫羽化后第2天用处女雄蛾与之自然交配1次;交配2次的处理:雌虫羽化后第2天与5头处女雄虫配对,观察其交配2次后取出雌虫;长期配对处理:选处女雌虫和雄虫配对,直至死亡;其余处理方法同前,每次交配间隔1 d;每处理观察20头。
1.4 雌雄不同年龄组配对成虫繁殖的影响
成虫的日龄设定:设羽化当晚的成虫日龄为0,第2天的成虫日龄为1,依此类推。处女雌虫推迟交配时间根据SAS软件设计的组合分12组,每组雌雄都有相对应的日龄(表1),每组处理配20对;把单头雌蛾引入直径16 c m玻璃瓶中,瓶底放一吸蜜糖水棉球,同时放入新鲜带叶片朱槿枝条,以便雌蛾在上面产卵。暗期前1 h每雌蛾引入1头对应日龄的处女雄蛾。在配对的当晚,每隔30 min用红光灯观察其交配行为。瓶中的雌雄蛾如果没有交配,则把它们去除,不再继续观察。如果能成功交配,每天定期查看叶片是否有野螟卵,并对枝条叶片进行保鲜,直到卵发育进入黑头期。同时记载雌蛾寿命、产卵期、产卵量、卵孵化率,直到雌蛾死亡。
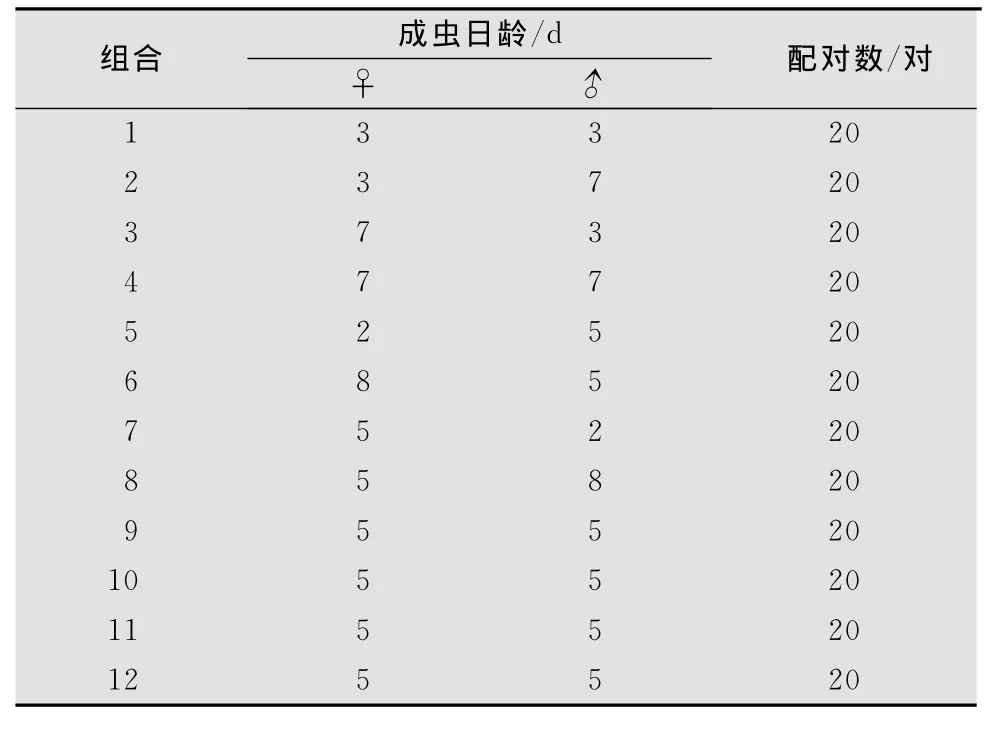
表1 不同的雌雄成虫年龄组配(SAS设计的组合)
1.5 资料的统计分析
所获数据用平均数±标准误(SE)来表示。数据经方差分析差异显著以后用Duncan多重比较法。统计程序为DPS v6.55版,图表制作在 Micr osoft Excel 2003版完成。
2 结果分析
2.1 交配持续时长对繁殖的影响
雌虫不同交配时长对其产卵前期有显著影响(F=2.971,p<0.05,df=5)(表2)。交配20 min的成虫产卵前期最长,平均为3.15 d,交配80 min的产卵前期最短,平均为1.71 d;交配20 min与交配40 min的产卵前期差异不显著,与交配80、120、180 min、自然交配1次的雌虫间,其产卵前期差异显著;而交配80、120、180 min与交配1次的雌虫的产卵前期无显著差异。
不同交配时长对雌虫的产卵期有显著影响(F=6.26,p<0.05,d f=5)(表2)。交配20、40 min的产卵期与交配80、120、180 min、交配1次之间差异显著,而交配20、40 min之间产卵期无显著差异,以交配20 min产卵期最短,平均为(2.31±0.58)d,交配80 min产卵期最长,平均为(6.58±0.82)d;而交配80、120、180 min、自然交配1次之间,其产卵期差异不显著。
不同交配时长对棉褐环野螟雌虫的产卵量有显著影响(F=10.191,p<0.05,df=5)(表2)。以交配80 min的雌虫产卵量最高,平均达218.50粒/雌,但与交配120、180 min、自然交配1次之间,其产卵量无明显差异;而交配20、40 min的处理,其产卵量较少,两处理间无显著差异。
雌虫每次的交配时间长短对卵孵化率有显著影响(F=7.072,p<0.05,d f=5)(表2)。雌虫交配时长短于20 min时,卵的孵化率仅为3.91%,交配40 min后其孵化率略有提高,但与交配20 min孵化率相比无显著差异;当交配时长达80 min以后时,其卵的孵化率迅速提高,平均达73.06%,以交配180 min的雌虫卵孵化率最高,达83.26%,但交配80、120、180 min、自然交配1次之间,其卵的孵化率无明显差异。由此可见,雌虫的交配时间至少应保持在80 min以上,其卵的孵化率才达到稳定。

表2 棉褐环野螟不同交配时长对雌虫产卵的影响1)
2.2 交配次数对繁殖的影响
雌虫不交配与交配1次、交配2次以及长期配对相比,其产卵期有显著性差异(F=10.53,p<0.05,df=3)(表3)。但交配1次、交配2次以及长期配对之间,其产卵期长短无显著性差异,表明雌虫一生接受一次交配即可满足其产卵需要,交配次数的多少对产卵期长短无影响,但不交配对产卵期有影响。
雌虫不交配对其产卵量有显著影响(F=8.92,p<0.05,df=3),与交配1次、交配2次以及长期配对的处理间差异显著;交配1次、交配2次以及长期配对的处理间差异不显著(表3)。其中以交配1次的雌虫产卵量最高,平均达(186.50±24.09)粒/雌,不交配的雌虫产卵量最低,平均仅(34.00±10.43)粒/雌,这说明雌虫不交配对其产卵量有影响,但雌虫交配次数的多少对其产卵量并无影响。
雌虫不同交配次数对卵的孵化率影响显著(F=11.18,p<0.01,df=3)(表3)。不经交配的雌虫所产下的卵均为无效卵,不能孵化;交配1次所产下的卵与交配2次以及长期配对的卵孵化率间无显著差异。
雌虫是否交配与一生中交配次数对雌虫的寿命会产生显著的影响(F=4.028,p<0.05,df=3)(表3)。以交配2次的雌虫寿命最长,平均达(14.23±0.48)d,明显长于未交配及交配1次的雌虫寿命,但与长期配对的处理相比,其雌虫寿命无显著差异。

表3 棉褐环野螟不同交配次数对雌虫产卵和寿命的影响1)
2.3 雌雄成虫不同年龄组配对繁殖的影响
雌雄成虫不同年龄组配对其产卵期会产生显著影响(F=4.04,p<0.05,df=11)(表4)。以♀3×♂7产卵期最长,平均达(5.87±0.58)d,明显长于♀5×♂2、♀5×♂8、♀5×♂5处理下的产卵期,而与♀3×♂3、♀7×♂3、♀7×♂7相比差异不显著;♀5×♂8产卵期最短(1.43±0.20)d。♀2×♂5与♀5×♂5相比在雄虫日龄一致而雌虫日龄不同的情况下,产卵期长短也存在差异,说明雌虫日龄对产卵期长短也有影响。♀5×♂2与♀5×♂5相比在雌虫日龄一致雄虫日龄不同的情况下,产卵期长短存在差异,由此说明雄虫日龄对产卵期的长短有一定的影响(表4)。
雌雄成虫不同年龄组配对其产卵量会产生显著影响(F=3.36,p<0.05,d f=11)(表4)。以♀3×♂7产卵量最高,平均达(189±17.17)粒/头,♀5×♂8产卵量最低,平均(41±5.53)粒/头。♀5×♂2与♀5×♂8相比在雌虫日龄一致雄虫日龄不同的情况下,产卵量存在一定的差异;♀2×♂5与♀8×♂5相比在雄虫日龄一致而雌虫日龄不同的情况下,产卵量也存在差异。雌雄成虫在7日龄之前(含7日龄),日龄的高低对产卵量有一定的影响,但影响不是很显著;当雌雄成虫达到8日龄之后对产卵量有显著影响,即当雌雄成虫达到一定日龄时对产卵量会产生显著影响。由此说明,延迟交配对其产卵量产生显著影响(表4)。
对卵孵化率的影响:雌雄成虫不同年龄组配♀3×♂7、♀7×♂3、♀7×♂7、♀2×♂5、♀8×♂5、♀5×♂2、♀5×♂8、♀5×♂5几种处理情况下卵的孵化率无显著差异(F=1.11,p>0.05),表明雌雄成虫日龄的大小对其孵化率的高低无影响。
♀3×♂7雌虫寿命最长,平均达(15.40±0.62)d,♀5×♂5与♀8×♂5相比雄虫日龄一致,雌虫日龄不一致的情况下,雌虫寿命无显著差异(F=2.88,p>0.05,d f=11)(表4)。由此说明,雌虫延迟交配对其寿命产生影响不显著。

表4 棉褐环野螟成虫日龄对雌虫产卵和寿命的影响
3 讨论
本研究发现,当棉褐环野螟成虫交配时间在80 min以上,其产卵量和卵的孵化率达到稳定,其中以交配时长180 min的卵孵化率最高,这可能是交配时间越长,雌虫所接受的精子量越多,其卵受精率和孵化率也随之增高,还需进一步研究证实。
试验结果还表明,交配和不交配处理间产卵期、产卵量、孵化率和寿命差异显著,但交配1次、交配2次以及长期配对之间,其产卵期、产卵量和幼虫孵化率无显著性差异,对寿命影响不大。不交配雌虫的产卵量最低,并且产下的是不育卵,均不能孵化。这说明该虫交配1次即可满足其产卵对精子的需要。这与前人关于鳞翅目其他昆虫的资料报道,即多次交配对于雌蛾的产卵和卵子的受精是必需的结论相反[7-8],而与焦晓国[9]对二化螟的研究结果一致。
不同年龄组配对繁殖适度最为显著的影响是,雌蛾的产卵量随着雌蛾交配日龄的增加而下降,与雌蛾相比,雄蛾交配日龄对雌蛾的生殖力影响较小。试验结果可以表明,雌蛾延迟交配2 d以上,其产卵量明显下降,这一现象在昆虫普遍存在[10-11]。在果园中进行新西兰云卷蛾(Cnephasia j actatana)干扰延迟交配试验表明,在该虫性成熟后延迟2~3 d,产卵量可降低27%,延迟4 d产卵量可降低88%。这在用性信息素防治害虫实践中无疑具有重要的意义。
不同年龄组配对其繁殖适度的影响机制尚不明确,有待进一步深入研究[13-14]。根据棉褐环野螟交配行为及能力和棉褐环野螟繁殖适度的研究结果,可进行其生物防治方面的研究。
[1] 吴时英.城市森林病虫害图鉴[M].上海:上海科学技术出版社,2005:53.
[2] 狄佳春,陈旭升,吴巧娟,等.棉大卷叶螟危害损失的初步研究[J].江苏农业科学,2007(5):82-83.
[3] Aplin R T,Birch M C.Identification of odor ous co mpounds from male Lepidoptera[J].Experientia,1970,26:1193.
[4] 杜家纬.昆虫信息素及其应用[M].北京:中国林业出版社,1988:1,9-40,133-180.
[5] Raina A K,Jaffe H,Kempe T G,et al.Identification of a neur opeptide hor mone t hat regulates sex pher o mone production in fe male mot hs[J].Science,1989,244:796-797.
[6] Ar n H,Tot h M,Priesner E.Listof sex pher o mone of Lepidoptera and related attractants[M].Paris:OILB-SROP,1992,75:16-20.
[7] Rogers C E,Marti O G.Beet ar my wor m(Lepidoptera:Noctuidae):effects of age at mating on repr oduction potential[J].Florida Ento mology,1996,79:343-352.
[8] Rogers C E,Marti O G.Once-mated beet ar my wor m (Lepidoptera:Noctuidae)effectof age at mating on fecundit y,fertility and longevity[J].Envir on mental Ento mology,1997,26:585-590.
[9] 焦晓国.性信息素在东北水稻二化螟种群预测及调控中的应用基础研究[D].北京:中国科学院,2004:43.
[10]焦晓国,宣维健,盛承发.水稻二化螟的交配行为[J].生态学报,2006,26(4):1110-1115.
[11]Torres-Vila L M,Rodríguez-Molina M C,Stockel J.Delayed mating reduces reproductive outputof female European grapevine mot h,Lobesia botr ana[J].Bulletin of Ento mological Research,2002,92:241-249.
[12]Wang Q,Shi G L.Mating frequency,duration,and circadian mating r hyt h m of New Zealand wheat bug Nysius huttoni White(Heter optera:Lygaeidae)[J].New Zealand Ento mologist,2004,27:113-117.
[13]Ar nqvist G,Danielsson I.Post mating sexual selection:the effects of male body size and recover y period on pater nity and egg production rate in a water strider[J].Behavioral Ecology,1999,10:358-365.
[14]刘芳,杨益众,陆宴辉,等.2005.转Bt基因棉对棉大卷叶螟种群动态的影响[J].昆虫知识,2005,42(3):275-277.
