黄河口春、秋季节无机碳的行为变化*
2012-01-08刘志媛张龙军张向上
刘志媛,张龙军**,张向上,2
(1.中国海洋大学海洋环境与生态教育部重点实验室,山东青岛266100;2.威海市海事局,山东威海264200)
黄河口春、秋季节无机碳的行为变化*
刘志媛1,张龙军1**,张向上1,2
(1.中国海洋大学海洋环境与生态教育部重点实验室,山东青岛266100;2.威海市海事局,山东威海264200)
根据2009年5、9月黄河口考察航次数据,结合2004年9月和2006年4月的调查结果,讨论了春季与秋季黄河河口过程中无机碳的行为、差异,以及调水调沙对黄河口无机碳输运过程的影响,并采用黑白瓶法对黄河口不同盐度水体进行现场培养,验证浮游生物活动对DIC的消耗。结果表明:春、秋季节黄河口淡咸水混合过程中无机碳的输运形态和量均发生了显著变化,pCO2在淡咸水混合过程中迅速下降,DIC在低盐区发生了亏损,淡水端DIC浓度越大亏损区间越大。夏季调水调沙后大量泥沙在口门附近沉积形成的拦门沙群使河口淡咸水混合过程在较短的距离内完成,也可导致秋季DIC的亏损区间小于春季。浮游生物活动对CO2的消耗是导致黄河口低盐度区发生DIC亏损的主要原因,但同时也伴随有碳酸钙沉降对DIC的清除作用。
淡咸水混合;无机碳;黄河口
河口具有天然“过滤器”效应,是理化参数变化最大、生物地球化学过程最活跃、沉积和再悬浮过程最显著的区域[1],剧烈的河口过程对陆地向海洋的物质输送产生显著影响,导致物质在海洋中的含量与分布发生变化。
全球河流每年向海洋输送约1Gt(1015g)碳,其中约60%为无机碳[2-3],但这一结论是依据河流入海口淡水端的观测数据获得的,并没有考虑河口过程的影响。对河口区溶解无机碳(DIC)的研究发现,许多河口都存在非保守现象。Cauwet et al 1985年就观测到黄河口低盐度区存在DIC亏损现象,认为可能是由于黄河水体中碳酸钙过饱和,在河口低盐度区发生沉降导致的[4];Abril et al发现法国的Loire河在河口低盐度区DIC迅速增加,认为是由于水体富营养化严重,异养呼吸产生CO2使pH降低导致碳酸钙发生溶解[5];Neal et al在研究欧洲Tweed,Great Ourse和Thames河流水体中碳酸钙饱和度的影响因素时,认为水体中浮游植物的光合作用较强,CO2含量下降,pH升高,使水体中的碳酸钙达到过饱和状态从而发生沉降[6];而珠江口DIC在盐度0~2.5之间的不保守增加则是由于河口西部高DIC含量的支流输入造成[7]。另外,河口一般具有比海洋高得多的pCO2,而毗邻河口的陆架边缘海pCO2较低,甚至表现为CO2的汇[8-9]。这都说明在河口淡咸水混合过程中无机碳的形态和量发生了急剧变化,仅依据淡水端观测数据估算无机碳入海通量将会导致其被高估或低估。因此厘清河口无机碳的行为变化有助于提高对近海碳格局的认识。
中纬度地区干旱半干旱的气候特征往往使得流经该区域的河流水体具有高浑浊度高无机碳含量。黄河流经黄土高原,是世界上泥沙含量最高的河流之一,流域内强烈的风化作用导致水体中无机碳含量很高,淡水端DIC浓度可达3.64mmol·L-1,不仅高于Amazon河0.67~0.83mmol·L-1[10],珠江1~1.25mmol·L-1[7],长江1.5~2.08mmol·L-1[11],也高于Mississippi河2.5~2.67mmol·L-1[12],印度Gatami河2.08~3mmol·L-1[13]等。另外,黄河水体颗粒无机碳(PIC)入海通量占总碳通量的71.4%,远远高于长江(17%)[4]。Cauwet et al虽指出黄河口低盐区DIC的亏损可能是由于碳酸钙沉降导致的,但限于相关参数的缺失,并未对其做出准确描述[4]。作者在“黄河口无机碳输运过程对pH异常增高现象的响应”一文中曾报道过,2004年秋季黄河口DIC在0<S<5的区域、2006年春季DIC在S<18的区域发生亏损现象,当时初步分析认为碳酸钙的沉降作用是其亏损原因之一[14]。
受自然及人为因素影响,近年来黄河入海水沙通量显著降低,河流系统发生了明显的变异[15-16]。2002年起黄河中下游连续3a进行了调水调沙原型试验,自2005年转为正式生产运作。自此每年6月下旬~7月初的调水调沙,使得大量径流和泥沙在短期内快速入海,对河口地形、地貌、生态环境及水沙条件等都产生重大影响,河口的物质输运过程发生变化[17]。本文根据2009年5、9月黄河口考察航次数据,并结合2004年9月和2006年4月的调查数据,讨论春季与秋季黄河河口过程中无机碳行为变化的差异,以及调水调沙对黄河口无机碳输运过程的影响。
1 实验部分
1.1 站位布设
2009年春、秋季调查区域及站位布设如图1(底图为2009年春季卫片)。黄河口浮桥为2航次走航测定的起点,在利津浮桥、垦利浮桥、建林浮桥、瞭望台等地设站作为淡水端站位,由于这些站位离河口较远,未在图中标出。2009年春、秋季采样期间利津水文站流量分别为270、380m3·s-1。文中还结合了本文作者在2004年9月、2006年4月对黄河口淡咸水混合区域的部分调查结果,其采样期间利津水文站流量分别为622、372m3·s-1。(流量数据来自中国水文信息网每日8时水情http://www.hydroinfo.gov.cn/)

图1 黄河口站位图Fig.1 Survey stations in the Yellow River Estuary
1.2 样品采集、处理及分析方法
表层水体温度(T)、盐度(S)、溶解氧(DO)及二氧化碳分压(pCO2)由走航式连续综合观测设备获得,其中,T和S由SBE 45MicroTSG(Sea-Bird Inc.,Bellevue,WA,USA)走航获得;pCO2是将表层海水经连续流动式水-气平衡器平衡并除湿后用Li-Cor 7000非色散红外分析仪测定;DO由YSI-5000溶氧仪(膜电极法)走航连续测定,温克勒法校正。
其它参数测定所需水样由上述走航式连续综合观测设备定点采集并现场过滤:
悬浮物含量(TSS):水样用孔径为0.45μm的醋酸纤维膜过滤,滤膜置于膜盒中冷冻保存,重量法测定。
叶绿素a(Chl a):用0.7μm GF/F Whatman玻璃纤维膜过滤水样,泵压<0.05Mpa,滤膜加饱和MgCO3溶液固定后在-20℃下避光保存。回实验室用10mL 9∶1丙酮溶液低温萃取24h后离心10min,取上清液用荧光分光光度计(F3010,Hitachi Co.,Tokyo,Japan)测定。
溶解无机碳(DIC):用0.45μm醋酸纤维膜过滤水样,滤液加饱和HgCl2溶液固定后于0~4℃环境中冷藏保存。用日本岛津公司TOC-VCPN分析仪测定。
颗粒无机碳(PIC):水样用0.7μm GF/F Whatman玻璃纤维膜过滤,滤膜冷冻保存。用日本岛津公司TOCVCPN分析仪及SSM-5000A(固体试样燃烧装置)测定。
总碱度(TA):用0.45μm醋酸纤维膜过滤水样,滤液加饱和HgCl2溶液固定,采用Gran作图-酸直接滴定法用AS-ALK2碱度滴定仪(Apollo SciTech)测定。
溶解有机碳(DOC):水样用预先450℃灼烧的0.7μm GF/F Whatman玻璃纤维膜过滤,滤液滴加饱和HgCl2溶液固定后0~4℃保存,采用TOC-VCPN总有机碳分析仪680℃高温燃烧催化氧化-非分散红外吸收法(NDIR)测定。
化学需氧量(CODMn):取样后24h内采用《海洋监测规范》(GB 17378.4-2007)中碱性高锰酸钾法测定。
2 结果与讨论
2.1 河口过程中连续走航观测参数(温度、盐度、DO、pCO2)的变化
图2为2009年春、秋季黄河口淡咸水混合过程中温度、盐度、DO、pCO2随航迹变化图,其它参数采样站位以●表示。该图以至黄河口浮桥(0km处)的距离为横坐标,走航开始较长一段距离内,水体盐度一直保持在0.5左右,温度、DO、pCO2也未见明显变化,因此该部分数据未绘入图中。
由图可见,2009年春、秋季黄河口盐度、DO、pCO2随航迹总体变化趋势一致:淡水端主要表现为黄河水体低盐、低DO、高pCO2的性质;随着淡咸水交汇,水体性质发生较大变化,盐度、DO、pCO2均在较短距离内出现剧烈波动,随着水体盐度逐步增大,pCO2不断降低,DO则与盐度呈良好的正相关性,在盐度20左右达到饱和;至海水端,黄河冲淡水作用减弱,淡咸水混合基本完成,盐度稳定在25以上,DO维持在过饱和状态,pCO2则下降到400μatm以下,高盐、高DO、低pCO2的海水性质显现出来。

图2 黄河口走航过程T,S,DO,pCO2沿程分布Fig.2 Distribution of continuously measured T,S,DO and pCO2in Yellow River Estuary
然而,由于季节、调水调沙等因素的影响,2个航次走航数据存在一定的差异,主要表现在以下两方面:首先,春季温度变化较大,且与盐度呈良好负相关(R2=0.667 8),而秋季水温明显高于春季,与盐度无明显关系,秋季整个走航过程温度变化很小,仅在距黄河口浮桥12~12.5km处升高1℃左右,原因可能是该区域处于拦门沙附近,水深较浅。第二,春、秋季黄河口淡咸水混合区域不同,春季发生在距离浮桥约8.5~14 km处,混合区域较长且混合过程较秋季更为剧烈。秋季淡咸水混合区域发生在距浮桥较远的口门附近,且在较短的距离内即完成混合,这可能是由于6月下旬~7月初的调水调沙,使大量泥沙在短期内快速入海,在口门处沉积形成拦门沙群[17],阻挡了淡咸水的交汇,河道内流速变缓,淡水较多,进而在口门附近又发生短距离内即完成混合的现象。另外,2个航次采样期间均处于涨潮阶段,潮汐作用对淡咸水混合过程的影响基本一致。
2.2 河口过程中pCO2的行为
图3是黄河口淡盐水混合过程4个调查航次pCO2随盐度变化图。从图中可看出,4个调查航次pCO2均表现出淡水端较高,随盐度的增加而降低,在低盐区迅速下降的趋势。但不同航次间pCO2的浓度及分布还是存在不少差异的,这与各航次间温度的差别,淡水端输入碳酸盐的浓度,以及有机物降解、浮游植物光合作用等有关。
为了考察温度对黄河口pCO2分布的影响,本文将秋季的pCO2校正到春季航次平均温度19.69℃(黄河口2个春季航次温度相差不大,2006年4月19.36℃,2009年5月20.03℃),2004年9月航次pCO2平均值由825μatm降低到721μatm,2009年9月pCO2平均值由582μatm降低到484μatm,说明温度是维持秋季高pCO2的1个重要因素。
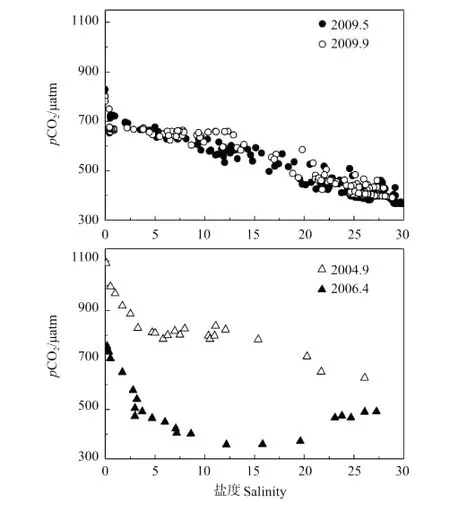
图3 河口过程pCO2与盐度ig.3 pCO2and salinity relationship during estuary dynamics
当去除温度影响后,2009年秋季pCO2(484 μatm)明显低于春季(553μatm),这可能是由于秋季黄河口水体中DIC、TSS、PIC含量明显低于春季(见2.3.1,图7~9),受碳酸盐体系平衡控制,相应的pCO2较低。另外,该航次的DOC也低于春季(见图4)。
2004年9月航次温度校正后的pCO2(721μatm)明显高于其它各航次,分析其原因,这个航次的DO是明显偏低的(见图5),自淡水端向海DO饱和度始终没有超过88%,可能是存在着显著的有机物降解过程,使DO偏低并产生了大量CO2,表现出具有较高的pCO2。通过查询黄河流域天气过程(中国气象科学数据共享服务网http://cdc.cma.gov.cn/)可知,在该航次采样之前的3天内黄河中、下游地区普遍降雨,雨水的地表冲刷导致大量植物碎屑及生活、工业排放的有机物进入黄河水体,而这些有机物不同于黄土土壤母质的有机碳,具有较强的易降解性。结合黄河口水体中CODMn的测定结果,2004年9月黄河口淡水端水体中CODMn平均值为5.62mg·L-1,远高于春季所测的2.49mg·L-1,说明了该航次黄河口DOC虽然低于春季(见图4),易降解有机物却明显较多。
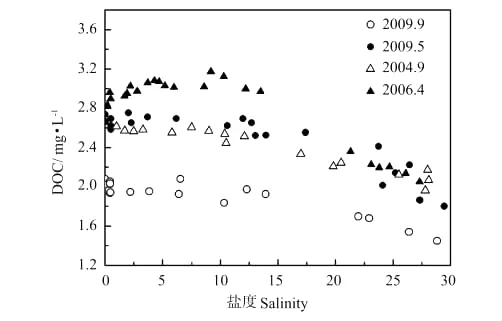
图4 河口过程DOC与盐度Fig.4 DOC and salinity relationship during estuary dynamic
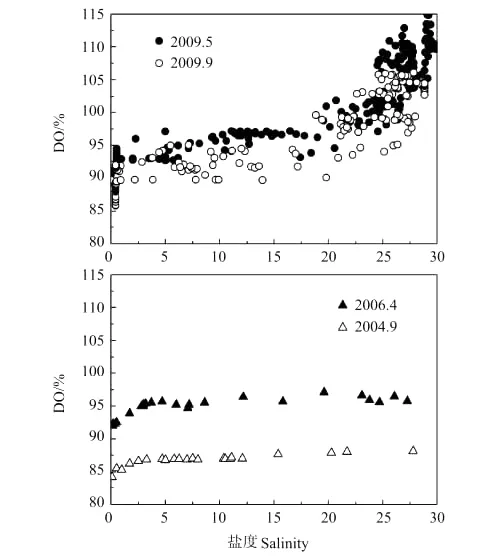
图5 河口过程DO与盐度Fig.5 DO and salinity relationship during estuary dynamics
另外,2006年4月pCO2在盐度5~20的水体明显偏低,甚至表现为CO2的汇(盐度15左右),可通过该航次盐度5~20附近区域Chla含量较高(见图6)来解释。
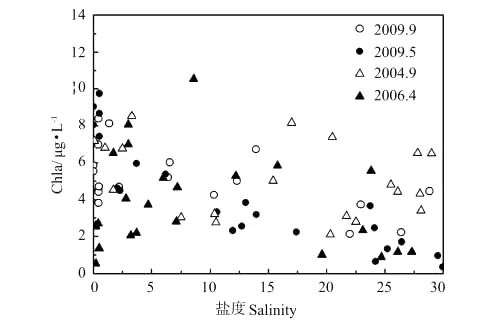
图6 河口过程Chl a与盐度Fig.6 Chlorophyll-a and salinity relationship during estuary dynamics
2.3 河口过程中DIC的行为
2.3.1 河口过程中DIC的分布 由图7可见,黄河口DIC总体表现为淡水端高于海水端的趋势,这是由于黄河流域内强烈的化学风化作用所致。各航次DIC均在低盐区发生亏损,即存在河口DIC的清除机制,这也是当前人们认识到的河口“过滤器”效应,淡水输入的DIC未能有效地输送到海洋。
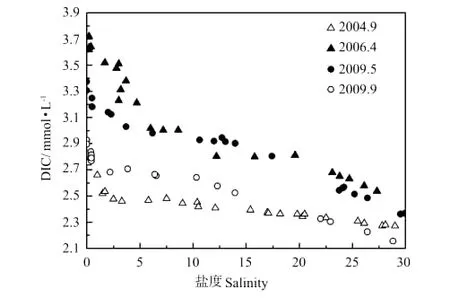
图7 河口过程DIC与盐度Fig.7 DIC and salinity relationship during estuary dynamics
黄河口不同调查航次间DIC的差别主要表现在:DIC亏损区间不同;秋季DIC浓度小于春季。
根据表1中Cauwet等及本实验室4个航次的DIC调查结果可以看出:黄河口淡水端DIC浓度越高,亏损区间越大;秋季DIC的亏损区间明显小于春季。对于秋季DIC亏损区间小于春季的原因,一是秋季DIC浓度小于春季,二是由于黄河夏季调水调沙,大量泥沙在口门附近沉积形成的拦门沙群阻挡了淡咸水的交汇,河道内水体流速变缓[17],使得秋季DIC亏损的河口过程在较短的距离内完成,正如走航图(见图2)所示,虽然秋季黄河流量大于春季(见表1),但其淡咸水混合区域却明显小于春季,从2009年黄河口站位布设(见图1)也可看出,春季航次要在较远的距离才能取到盐度大约为30的海水区域,而秋季航次则在河道口门之外很快就达到了盐度大约为30的海水区。
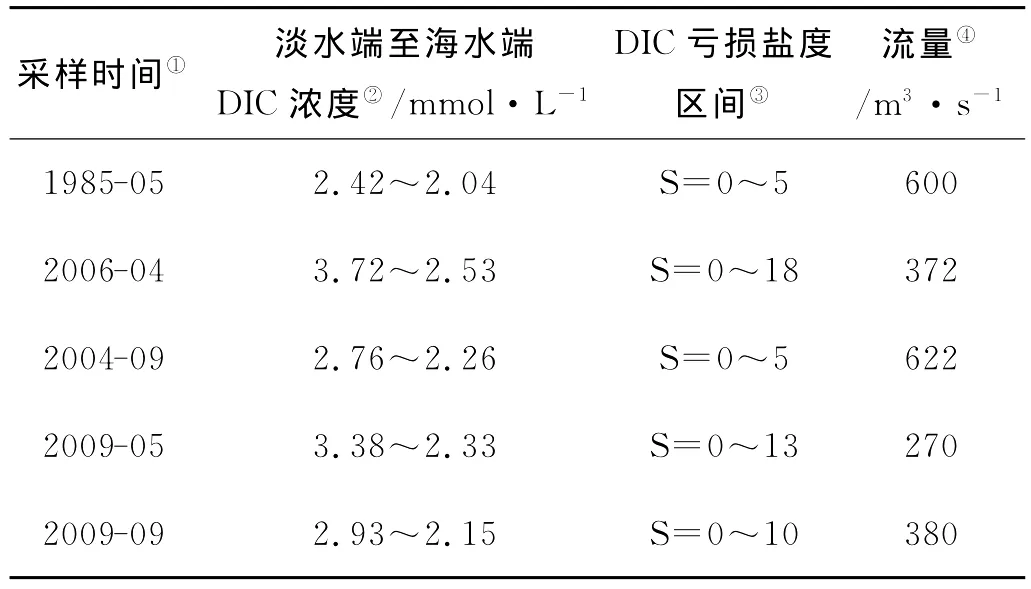
表1 不同航次DIC浓度及亏损区间Table 1 Concentration and dificit area of DIC in different surveys
秋季DIC浓度小于春季的原因,一是秋季黄河流量大,稀释作用导致秋季DIC浓度低;二是控制黄河及河口DIC浓度变化的另一参量是TSS,或者说是PIC的量。作者曾报道了黄河干流PIC=0.013 2TSS(R2=0.981 3)、DIC=0.797 3PIC(R2=0.865 9),富含碳酸盐的黄河悬浮颗粒物通过碳酸盐体系的固-液平衡控制着水体的DIC浓度[18]。由图8、9中TSS、PIC随盐度的变化可以看出,秋季TSS、PIC显著低于春季,这也是秋季DIC浓度低于春季的原因之一。而秋季黄河口TSS、PIC显著低于春季是由于拦门沙群的存在,河道内水流减缓、泥沙沉降速度加快。
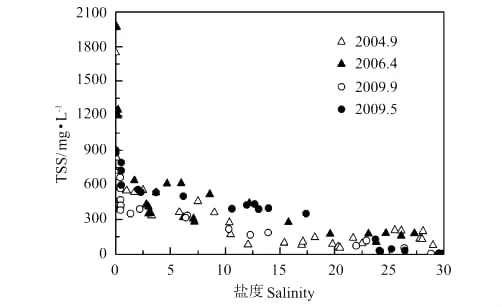
图8 河口过程TSS与盐度Fig.8 TSS and salinity relationship during estuary dynamics
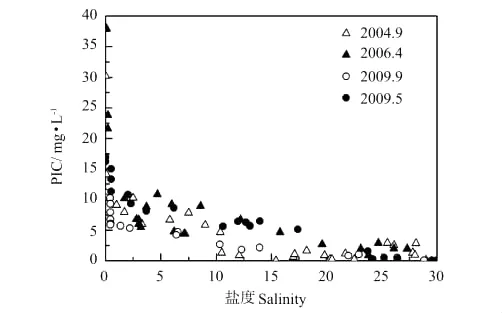
图9 河口过程PIC与盐度Fig.9 PIC and salinity relationship during estuary dynamics
2.3.2 DIC亏损原因探讨 河口区DIC与盐度不保守的原因主要有:①支流及外源输入,该方法引起ΔDIC和ΔTA呈1:1的比率变化;②好氧呼吸和光合作用能改变DIC的量,但几乎不引起TA变化;③碳酸钙的溶解和沉降分别可导致DIC的不保守增加和降低,该原因引起的ΔDIC和ΔTA的比率为1∶2;④沉积物及悬浮物中颗粒态的磷在淡咸水混合过程中向水体释放磷酸盐,磷酸盐为生物所利用并消耗水体中的硝酸盐、硅酸盐及DIC,造成DIC的不保守行为[7]。由于黄河下游是地上悬河,河口无外源DIC输入;黄河水体的磷酸盐含量普遍较低(4个航次数值在0~1.2 μmol·L-1之间);另外,黄河口DIC的不保守表现为低盐区亏损,未出现盈余。因此主要应从浮游生物活动对DIC的消耗,以及碳酸钙的沉降两方面讨论黄河口DIC的不保守原因。
为考察浮游生物活动对DIC的消耗,春季作者采用黑白瓶法分别对黄河口DIC亏损区域4种盐度水体进行培养,测定不同培养条件下的初级生产力和水体总耗氧速率,同时对白瓶中的DIC量进行了测定,以确定光合作用及耗氧呼吸对DIC的可能影响。
初级生产力及耗氧呼吸速率对应的氧收支可以根据下式计算[11,19-20]:
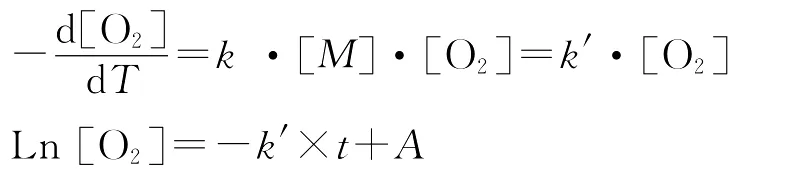
式中[O2]表示DO浓度;t代表培养时间;A为任意常数;k′(DO的自然对数对时间的斜率)在白瓶中表示光合作用净反应速率常数,与DO的乘积即净初级生产力对应的氧收入;在黑瓶中表示有机物降解准一级反应速率常数k′,与DO的乘积即总耗氧速率对应的氧支出;两者差值即为总初级生产力对应的氧收支。

表2 净初级生产力、耗氧呼吸速率及DIC消耗量Table 2 net primary productivity,respiration rate and consumption of DIC
由于光合作用及耗氧呼吸过程中O2与CO2变化量近似为1∶1的关系,因此可以通过DO的变化推算DIC变化量[21-22]。由表2可看出,耗氧呼吸对DIC的影响明显小于光合作用;白瓶中实测DIC消耗量与DO换算的DIC消耗量相比,两者在趋势上具有较好的一致性,在盐度为2.4时光合作用对DIC的消耗量最高,随着盐度的增加DIC的消耗量逐渐降低,由此推测,浮游植物光合作用吸收CO2导致的DIC净消耗在低盐度区更为显著,是低盐区DIC发生亏损的原因之一。
表2还显示:白瓶中实测DIC消耗量,与通过DO换算的DIC消耗量存在差异,特别是低盐度区DIC实测值大于DO的换算值。当然这里会存在2种获取数据的方法误差问题,但本文更倾向于黄河口低盐度区DIC的亏损除生物活动外还存在其他影响机制,如碳酸钙的沉降。
黄河口发生碳酸钙沉降的可能性是存在的,Gan et al.曾对黄河干流碳酸钙的饱和度进行了计算,指出黄河水体碳酸钙整体呈现过饱和状态[23]。在河口区,淡咸水混合使水体离子强度增大,Ca2+迅速增加,pCO2的降低使pH升高等,都有可能导致黄河口低盐度区过饱和的碳酸钙发生沉降。
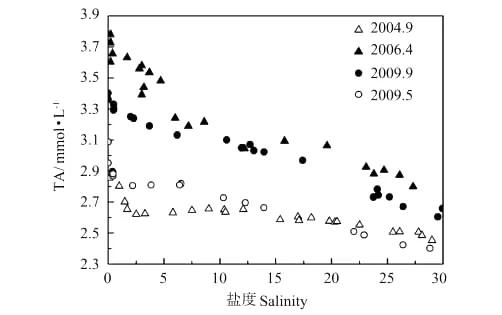
图10 河口过程TA与盐度Fig.10 TA and salinity relationship during estuary dynamics
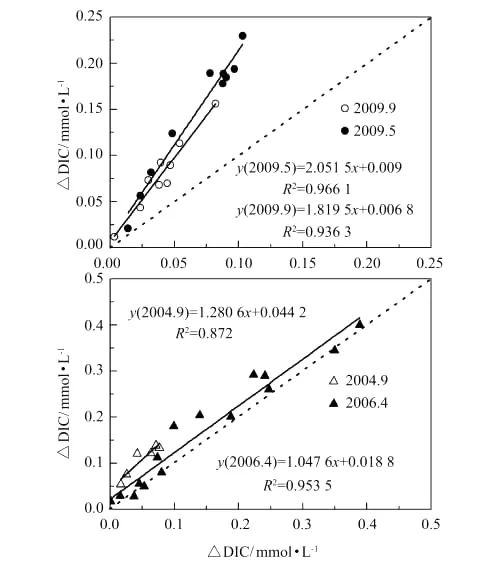
图11 ΔDIC与ΔTA相关性Fig.11 Correlation ofΔDIC andΔTA
由黄河河口过程中TA随盐度变化(见图10)可以看出,这4个航次在DIC发生亏损的区域,TA也同样发生了亏损。单一的光合作用对DIC的消耗,是不会引起TA变化的;而黄河口NH4-N含量低至不足以引起TA发生明显变化(2009年春季0.04~2.31μmol·L-1,2009年秋季1.48~5.52μmol·L-1);黄河口不缺氧,有机弱酸对TA的影响也很小。因此,TA的亏损可能是由于碳酸钙的沉降引起的。
图11是黄河口DIC亏损区域内ΔDIC与ΔTA的相关图。由图可见,各航次黄河口低盐度区ΔDIC/ΔTA均>1,即DIC的亏损量远远大于TA的亏损,说明浮游生物活动对CO2的消耗是导致DIC亏损的主要原因,但同时伴随有碳酸钙的沉降(TA亏损)对DIC的清除作用。
3 结论
(1)黄河口淡咸水混合过程中无机碳的输运形态和量发生了显著变化,pCO2在淡咸水混合过程中迅速下降,但由于输入淡水性质(如流域降雨)的差别和河口浮游生物光合作用的强弱,各航次pCO2的分布存在差异。
(2)DIC在低盐区发生了亏损,淡水端DIC浓度越大发生亏损的区间越大。夏季调水调沙大量泥沙在口门附近沉积形成的拦门沙群,使河口混合过程在较短的距离内完成,也可导致秋季DIC的亏损区间小于春季。
(3)通过现场生物培养实验,以及对DIC、TA的亏损程度(ΔDIC、ΔTA)的分析可知,浮游植物光合作用对CO2的消耗是导致黄河口低盐度区DIC发生亏损的主要原因,但同时也伴随有碳酸钙沉降对DIC的清除作用。
[1] Gattuso J P,Frankignoulle M,Wollast R.Carbon and carbonate metabolism in coastal aquatic ecosystems[J].Annual Review of Ecology and Systematics,1998,29:405-434.
[2] Hope D,Billet M F,Cresser M S.A review of the export of carbon in river water:fluxes and processes[J].Environmental Pollution,1994,84:301-324.
[3] Ludwig W,Probst J,Kempe S.Predicting the oceanic input of organic carbon by continental erosion[J].Global Biogeochem Cycles,1996,10(1):23-41.
[4] Cauwet G,Mackenzie F T.Carbon inputs and distribution in estuaries of turbid rivers:the Yang Tze and Yellow rivers(China)[J].Marine Chemistry,1993,43:235-246.
[5] Abril G,Etcheber H,Delille B,et al.Carbonate dissolution in the turbid and eutrophic Loire estuary[J].Marine Ecology Progress Series,2003,259:129-138.
[6] Neal C.Calcite saturation in eastern UK rivers[J].The Science of the Total Environment,2002,282:311-321.
[7] Cai W J,Dai M H,Wang Y C,et al.The biogeochemistry of inorganic carbon and nutrients in the Pearl River estuary and the adjacent Northern South China Sea[J].Continental Shelf Research,2004,24:1301-1319.
[8] Liu K K,Iseki K,Chao S Y.Continental marine carbon fluxes[M].∥Hanson R B,Ducklow H W,Field J G,eds.The Changing Ocean carbon cycle.Cambridge:Cambridge University Press,2000:187-239.
[9] Frankignoulle M,Borges A.European continental shelf as a significant sink for atmospheric carbon dioxide[J].Global Biogeochemical Cycles,2001,15:569-576.
[10] Ternon J F,Oudot C,Dessier A,et al.A seasonal tropical sink for atmospheric CO2in the Atlantic ocean:the role of the Amazon River discharge[J].Marine Chemistry,2000,68(3):183-201.
[11] Zhai W D,Dai M H,Guo X H.Carbonate system and CO2degassing fluxes in the inner estuary of Changjiang(Yangtze)River,China[J].Marine Chemistry,2007,107(3):342-356.
[12] Cai W J.Riverine inorganic carbon flux and rate of biological uptake in the Mississippi River plume[J].Geophysical Research Letter,2003,30(2):1-4.
[13] Bouillon S,Frankignoulle M,Dehairs R,et al.Inorganic and organic carbon biogeochemistry in the Gartami Godavari estuary(Andhra Pradesh,India)during pre-monsoon:The local impact of extensibe mangrove forests[J].Global Biogeochemical Cycles,2003,17(4):1-12.
[14] 张向上,张龙军.黄河口无机碳输运过程对PH异常增高现象的响应[J].环境科学,2007,28(6):1216-1222.
[15] Wang H J,Yang Z S,Saito Y,et al.Interannual and seasonal variation of the Huanghe(Yellow River)water discharge over the past 50years:Connections to impacts from ENSO events and dams[J].Global and Planetary Change,2006,50(3-4):212-225.
[16] Wang H J,Yang Z S,Saito Y,et al.Stepwise decreases of the Huanghe(Yellow River)sediment load(1950-2004):Impacts of climate changes and human activities[J].Global and Planetary Change,2007,57:331-354.
[17] 王厚杰,杨作升,毕乃双,等.2005年黄河调水调沙期间河口入海主流的快速摆动[J].科学通报,2005,50(23):2657-2664.
[18] 张龙军,徐雪梅,温志超.秋季黄河pCO2控制因素及水-气界面通量[J].水科学进展,2009,20(2):227-235.
[19] Zhai W D,Dai M H,Cai W J,et al.High partial pressure of CO2and its maintaining mechanism in a subtropical estuary:the Pearl River estuary,China[J].Marine Chemistry,2005,93(1):21-32.
[20] Dai M H,Guo X H,Zhai W D,et al.Oxygen depletion in the upper reach of the Pearl River estuary during a winter drought[J].Marine Chemistry,2006,102(1-2):159-169.
[21] Jiang L Q,Cai W J,Wang Y C,et al.Pelagic community respiration on the continental shelf off Georgia USA[J].Biogeochemistry,2009,98(1-3):101-113.
[22] Hopkinson C S.Shallow-water benthic and pelagic metabolism:evidence of heterotrophy in the nearshore Georgia Bight[J].Marine Biology,1985,87:19-32.
[23] Gan W B,Chen H M,Han Y F.Carbon transport by the Yangtze(at Nanjing)and Huanghe(at Jinan)rivers,People's Republic of China[R].∥Degens E T,Kempe S,Soliman S,eds.Transport of Carbon and Minerals in Major World Rivers,Part 2.Mitt Geol-Pal ontol Inst Univ Hamburg,SCOPE/UNEP Sonderd.Cambridge:Universit it Hamburg,1983,55:459-470.
The Changing Behavior of Inorganic Carbon in Yellow River Estuary Between Spring and Fall
LIU Zhi-Yuan1,ZHANG Long-Jun1,ZHANG Xiang-Shang1,2
(1.The Key Lab of Marine Environmental Science and Ecology,Ministry of Education,Ocean University of China,Qingdao 266100,China;2.Weihai Maritime Safety Administration,Weihai 264200,China)
Difference of changing behaviors of inorganic carbon and the influence of Water and Sand Regulate Scheme(WSRS)on the estuarine dynamics between spring and fall were discussed based on the data obtained from May and September,2009,combined with the data we discussed before(September,2004and April,2006).Furthermore,in-situ incubation of the different salinity water in the Yellow river estuary through the method of black-white bottles was carried out in spring 2009,which aims to identify the DIC consumption of plankton activities.The Results showed that:The mass and the form of the inorganic carbon had notably changed both in spring and fall during the estuarine mixing process,during which partial pressure of carbon dioxide(pCO2)decreased rapidly and deficit of dissolved inorganic carbon(DIC)occurred in the low-salinity area,The higher concentration of DIC would lead to a larger deficit area.A group of sandbars which was caused by a large amount of sediments depositing in the mouth of the estuary due to WSRS in summer made the estuarine mixing process accomplish in a short distance,so the deficit area of DIC in fall is smaller than spring.Phytoplankton activity which consumed CO2was the main reason which made deficit of DIC occur in the low-salinity area of Yellow River Estuary,however there still existed a scavenging process of DIC by calcium carbonate precipitation.
estuarine mixing process;inorganic carbon;Yellow River Estuarine
X143
A
1672-5174(2012)1-2-041-08
国家自然科学基金主任基金项目(40940019);海洋公益性专项(20080508006)资助
2011-03-21;
2011-04-15
刘志媛(1985-),女,硕士生。E-mail:lctuyuanzi123@163.com
**通讯作者:E-mail:longjunz@ouc.edu.cn
责任编辑 庞 旻
