钩虾和蜾蠃蜚群落在日本对虾养殖池塘中变动规律的研究*
2012-01-08钟源,马甡
钟 源,马 甡
(中国海洋大学海水养殖教育部重点实验室,山东青岛266003)
钩虾和蜾蠃蜚群落在日本对虾养殖池塘中变动规律的研究*
钟 源,马 甡**
(中国海洋大学海水养殖教育部重点实验室,山东青岛266003)
2009年4~7月,对移入日本对虾养殖池的钩虾和蜾蠃蜚群落进行了研究。共调查到2种钩虾,3种蜾蠃蜚。钩虾的优势种为中华原钩虾;蜾蠃蜚的优势种具有明显的交替性,放养后前53d,优势种为大蜾蠃蜚,78d后,优势种为隐居蜾蠃蜚。钩虾群落的物种多样性(H′)与均匀度(J)在放养当天达到最高,放养后第67天出现差异显著;蜾蠃蜚群落的物种多样性(H′)与均匀度(J)在放养后第64天最高,第90天出现差异显著。钩虾的生物量和丰度放养后第30天达到最大值,随后快速下降;蜾蠃蜚的生物量和丰度放养后第14天达到最大值,其丰度在第64天达到第二个高峰。同时,钩虾和蜾蠃蜚群落不同体长个体的丰度百分比差异明显,表现出一定的周期性变化规律。由此,初步分析了引起群落变化的两类原因,提出了钩虾和蜾蠃蜚群落结构的调控方案。
钩虾;蜾蠃蜚;群落;变动规律;虾池
钩虾、蜾蠃蜚隶属于甲壳纲,端足目,钩虾亚目[1],栖息于潮间带、河口地区的海藻和底泥中,营底栖生活,主要以藻类和有机碎屑等为食,我国海域均有分布[2]。钩虾和蜾蠃蜚含有丰富的蛋白质[3-4],并具有营养价值高、不污染水质等特点,是对虾优良的生物饵料[5-6]。
王克行[7]首次提出在虾池中移植和繁殖底栖饵料生物的设想。韩方训[8]、张志南[9]、周一兵[10]、邓锦松[11]向对虾养殖池中移植沙蚕,大量摄食虾池中的残饵、粪便和有机碎屑等,取得了良好的效果。在日本对虾养殖过程中移植入底栖生物钩虾和蜾蠃蜚,在我国北方已成为1种比较流行的模式[12]。迄今为止,对钩虾和蜾蠃蜚的调查多限于海域的底栖生物调查[13-15],池塘养殖系统中钩虾和蜾蠃蜚的群落变动鲜有报道。为此,本文探讨了钩虾和蜾蠃蜚群落在日本对虾养殖池中的变动规律,旨在为今后优化日本对虾生态养殖模式,提高钩虾和蜾蠃蜚等饵料生物的养殖效益,提供基础数据。
1 材料和方法
1.1 实验条件
本实验于2009年4~7月,在青岛市宝荣水产有限公司东区8#、9#和11#养殖池进行。上述养殖池为改造后土池,面积1.6hm2,属露天泥沙质池塘。在虾池进水前20d,用生石灰兑水对虾池彻底消毒,杀灭池中全部的野生杂鱼。养殖用水为地下咸水,盐度为23~26,经100目筛绢过滤进入养殖池塘,采用仅添水不排水的封闭式养殖模式。4月9日移植入钩虾每池50kg,4月12日移植入蜾蠃蜚每池25kg,4月15日移入以硬毛藻(Chaetomorpha sp.)为主的水草,供钩虾和蜾蠃蜚摄食,移入量为每池360kg。池中日本对虾的养殖分为2期进行,每期的起止时间与放苗情况如表1。

表1 日本对虾养殖情况Table 1 The state of cultivation of Marsupenaeus japonicus
1.2 实验方法
实验期间,每养殖池塘设5个采样点,进出水口各1个点,中央区3个点,用铁锹挖取底泥[16],每个采样点定量取3个样方,采样面积为0.017m2,将所采底泥倒入过滤网(网目0.037mm),洗净污泥后将网中剩余物倒入塑料瓶中,10%福尔马林固定后,带回实验室进行分类计数,体长测量,生物量(湿质量)测定和种类鉴定[1]等。标本的处理和分析均参照《海洋调查规范》和《全国海岸带和海涂资源综合调查简明规程》进行。
实验期间每隔15d左右检查日本对虾的生长情况,每池取对虾不少于40尾,收获时取样不少于80尾,测量其体长和体质量。
1.3 实验管理
实验开始前1个月池塘使用漂白粉消毒,3口池塘于2009年3月14日进水,实验期间每月换水1次,并保持水深(150±20)cm。一期养殖期间不投饵,二期放苗后,只投喂新鲜蓝蛤,每天投饵2次,分别为06:00和17:00。每天观察各池内残饵情况,并根据日本对虾摄食情况和水温天气情况等,及时调整日投饵量,各池的日投饵量见表2。
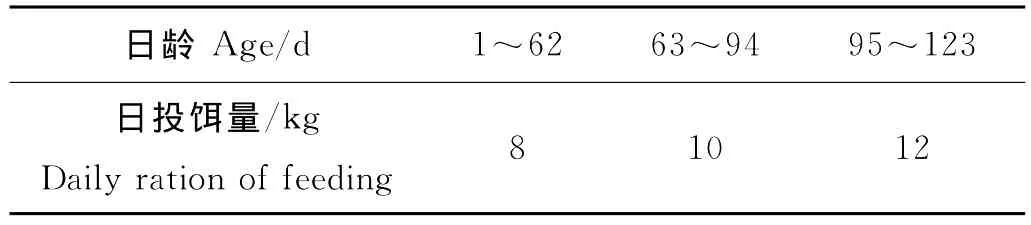
表2 日本对虾的日投饵量Table 2 The amount of daily diet of Marsupenaeus japonicus
1.4 物种多样性的计算
使用下列公式[17-18],进行分析。
Pielou均匀度指数J=H′/log2S;
Marglef种类丰富度指数R=(S-1)/log2N。
式中,Pi为种i的个体数占总个体数的比例;N为所有种的个体总数;S为总种数。
1.5 数据的统计分析
数据以平均值±标准差(MEAN±SD)表示,利用Excel 2003和SPSS 13.0软件对实验数据进行统计分析。采用单因子方差分析(One-way ANOVA)和Duncan法进行多重比较分析,取P<0.05为差异显著。
2 结果与分析
2.1 钩虾和蜾蠃蜚群落的组成
实验过程中,调查到2种钩虾,隶属2科2属,分别为中华原钩虾(Eogammarus sinensis Ren)和强壮藻钩虾(Ampithoe valida Smith)。调查到3种蜾蠃蜚,隶属1科1属,分别为大蜾蠃蜚(Corophium major Ren)、隐居蜾蠃蜚(Corophium insidiosumCrawford)和河蜾蠃蜚(Corophium acherusicumCosta)。
2.2 钩虾和蜾蠃蜚的生物量与丰度的变化
各池钩虾的生物量和丰度变化分别见图1和图2,钩虾生物量和丰度在放养后第30天达到峰值,分别为(4.54±1.05)g·m-2和(146.67±42.14)inds·m-2,随后快速下降。蜾蠃蜚的生物量和丰度变化分别见图3和图4,蜾蠃蜚生物量和丰度在放养14d时达到峰值,分别为(2.52±0.54)g·m-2和(219.17±66.14)inds·m-2;在64d时达到第2个高峰,分别为(0.79±0.25)g·m-2和(99.68±32.79)inds·m-2;第78天起又开始缓慢下降。
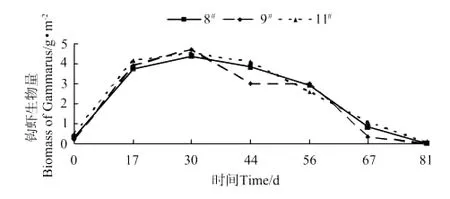
图1 日本对虾养殖池中钩虾生物量的变化Fig.1 The variations of biomass of gammarid amphipods in the farm ponds of Marsupenaeus japonicus
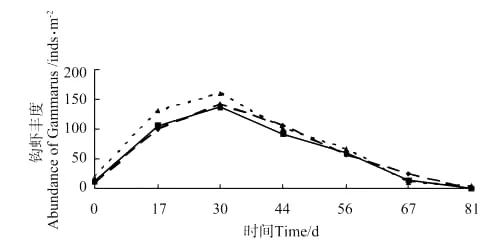
图2 日本对虾养殖池中钩虾丰度的变化Fig.2 The variations of abundance of gammarid amphipods in the farm ponds of Marsupenaeus japonicus
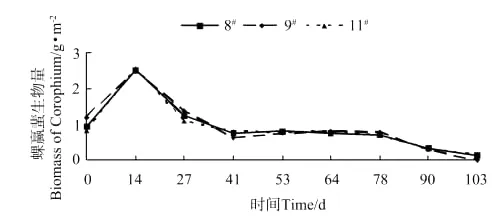
图3 日本对虾养殖池中蜾蠃蜚生物量的变化Fig.3 The variations of biomass of corophid amphipods in the farm ponds of Marsupenaeus japonicus
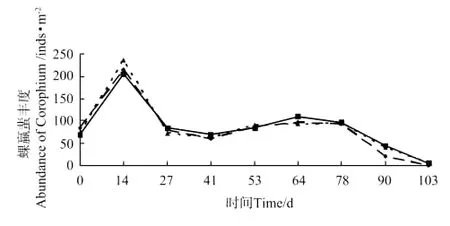
图4 日本对虾养殖池中蜾蠃蜚丰度的变化Fig.4 The variations of abundance of corophid amphipods in the farm ponds of Marsupenaeus japonicus
由图5可知,中华原钩虾(E.sinensis)为养殖池中钩虾群落的优势种,其丰度百分比在放养后第30天出现显著差异(P<0.05),第67天达到最大值。强壮藻钩虾(A.valida)丰度百分比在放养后第30天出现显著差异(P<0.05),第67天降到最低点。
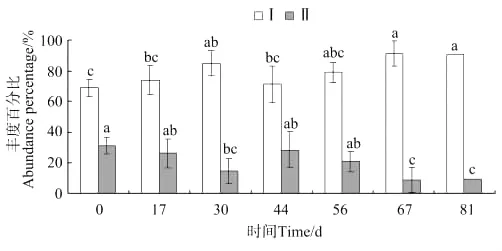
图5 钩虾各物种丰度百分比的变化Fig.5 The variations of abundance percentage of two gammarid ammphipodspecies
蜾蠃蜚群落中,其优势种出现明显的交替性(见图6)。放养53d前,优势种为大蜾蠃蜚(C.major);放养78d后,优势种为隐居蜾蠃蜚(C.insidiosum)。大蜾蠃蜚(C.major)在前期缓慢增加,放养第53天达到最大值后,出现快速下降。隐居蜾蠃蜚(C.insidiosum)所占的丰度百分比先降低后升高,第53天达到最低点,随后快速增加,至第103天达到最大值。河蜾蠃蜚(C.acherusicum)前期缓慢增加,至放养后第41天出现显著差异(P<0.05),随后逐渐下降,第103天降至最小值。
通过测量不同生长时期日本对虾的体质量,分别得到日本对虾体质量(Y)与钩虾生物量(BG)和蜾蠃蜚生物量(BC)的回归方程如表下:Y=-0.402 4 BG+4.154 3(R2=0.952 9);Y=0.93 Bc-0.1731(R2=0.173 1)。
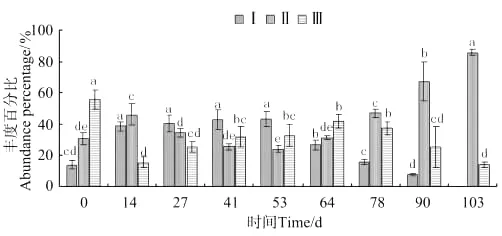
图6 蜾蠃蜚各物种丰度百分比的变化Fig.6 The variations of abundance percentage of three corophid amphipodspecies
2.3 钩虾和蜾蠃蜚群落结构分析
2.3.1 钩虾和蜾蠃蜚群落物种多样性的变化 从图7可以看出,养殖池中的钩虾群落,Shanno-Wiener指数(H′)、Pielou均匀度指数(J)和Marglef种类丰富度指数(R)均是放养当天最高,放养81d时最低。其中H′和J在放养后第67天出现显著差异(P<0.05)。
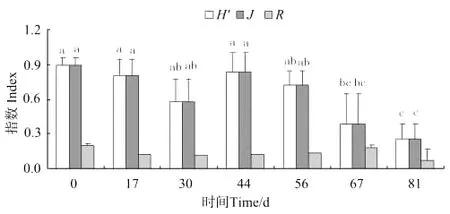
图7 钩虾群落物种多样性的变化Fig.7 The patterns of species diversity of gammarid amphipodcommunity
由图8可知,养殖池中的蜾蠃蜚群落,Shannon-Wiener指数(H′)、Pielou均匀度指数(J)在放养27d后达到最高,第103天时最低;在放养后第90天出现显著差异(P<0.05)。Marglef种类丰富度指数(R)则是在放养后第90天达到最大值。
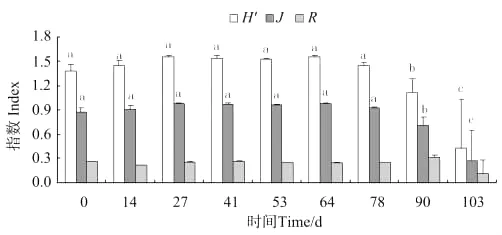
图8 蜾蠃蜚群落物种多样性的变化Fig.8 The patterns of species diversity of corophid amphipodcommunity
2.3.2 钩虾和蜾蠃蜚群落个体体长变化 钩虾群落个体体长变化如图9所示,体长0.038~0.2mm钩虾的丰度百分比,在放养第17天出现显著差异(P<0.05),第81天达到最大值。体长0.2~0.5mm钩虾,在放养当天达到最大值,第56天出现显著差异(P<0.05)。体长0.5mm以上的钩虾,放养第17天出现最大值,第67天出现显著差异(P<0.05),随后消失。
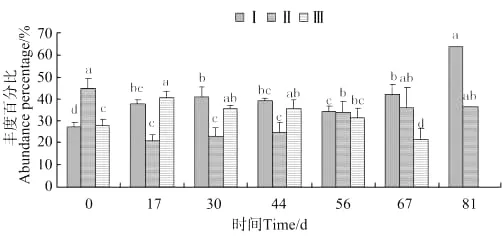
图9 钩虾个体体长丰度百分比的变化Fig.9 The variations of abundance percentage of individual body length of gammarid amphipods
中华原钩虾种群中个体体长变化如图10所示,体长0.038~0.2mm钩虾的丰度百分比,第30天起开始下降至第44天出现显著差异(P<0.05),第56天达到最低值,随后开始上升至第81天达到最高。体长0.2~0.5mm钩虾的丰度百分比放养当天最高,第17天出现显著差异(P<0.05)并达到最小值,至56天出现第二个丰度高峰后下降。体长0.5mm以上钩虾的丰度百分比自放入起缓慢上升,至第44天出现显著差异(P<0.05)并达到最大值,随后下降至第81天消失。
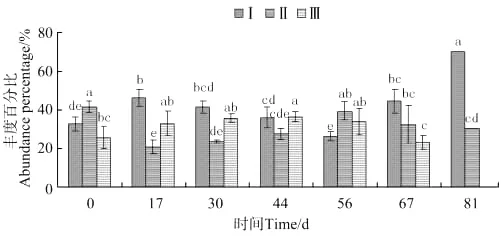
图10 中华原钩虾个体体长丰度百分比的变化Fig.10 The variations of abundance percentage of individual body length of E.sinensis
强壮藻钩虾种群中个体体长变化如图11所示,体长0.038~0.2mm钩虾的丰度百分比,第30天出现显著差异(P<0.05)并缓慢上升,至第56天达到最大值快速下降。体长0.2~0.5mm钩虾的丰度百分比放养后17天出现显著差异(P<0.05),17~56d内无显著性差异,并在第56天达到最小值,第67天出现显著差异(P<0.05)并达到最大值。体长0.5mm以上钩虾的丰度百分比放入第17天达到最大值,第30天出现显著差异(P<0.05),30~56d内无显著性差异,第56天达到最小值随后消失。
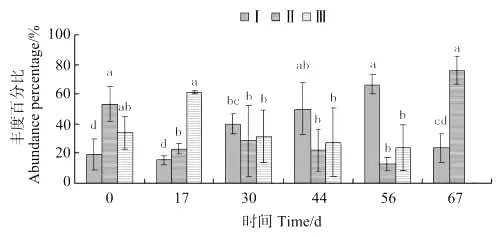
图11 强壮藻钩虾个体体长丰度百分比的变化Fig.11 The variations of abundance percentage of individual body length of A.valida
蜾蠃蜚群落个体体长变化如图12所示,体长0.038~0.2mm蜾蠃蜚的丰度百分比,在放养后第64天出现显著差异(P<0.05),第103天达到最小值。体长0.2~0.5mm蜾蠃蜚,第90天出现显著差异(P<0.05),第103天达到最大值。体长0.5mm以上的蜾蠃蜚,放养第14天出现最大值,第64天出现显著差异(P<0.05)。
大蜾蠃蜚种群中个体体长变化如图13所示,体长0.038~0.2mm蜾蠃蜚的丰度百分比,在放养90天前差异不显著,至第90天出现显著差异(P<0.05)并达到最大值。体长0.2~0.5mm蜾蠃蜚,在放养64天前差异不显著,至第64天出现显著差异(P<0.05)并达到最大值,第78天出现显著下降。体长0.5mm以上的蜾蠃蜚,在放养78d前差异不显著,至第78天出现显著差异(P<0.05)并达到最大值,随后消失。
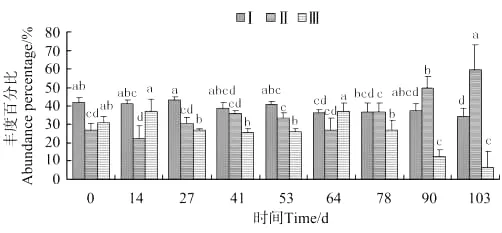
图12 蜾蠃蜚个体体长丰度百分比的变化Fig.12 The variations of abundance percentage of individual body length of corophid amphipods
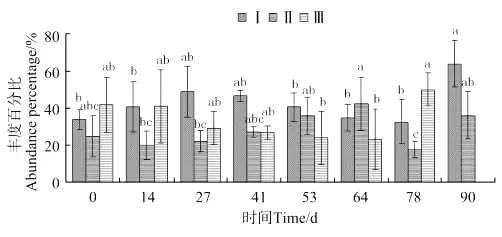
图13 大蜾蠃蜚个体体长丰度百分比的变化Fig.13 The variations of abundance percentage of individual body length of C.major
隐居蜾蠃蜚种群中个体体长变化如图14所示,体长0.038~0.2mm蜾蠃蜚的丰度百分比,第14天出现最小值,至第78天出现显著差异(P<0.05)并达到最大值。体长0.2~0.5mm蜾蠃蜚丰度百分比,前期缓慢上升至41d出现显著差异(P<0.05),随后快速下降至第64天出现最小值,然后再度上升至第90天出现显著差异(P<0.05),第103天达到最大值。体长0.5 mm以上的蜾蠃蜚,放养第14天达到最大值,随后快速下降至第41天出现显著差异(P<0.05)并达到最小值,其后丰度百分表现为先上升后下降,至第103天消失。
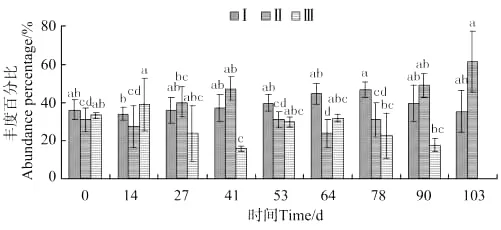
图14 隐居蜾蠃蜚个体体长丰度百分比的变化Fig.14 The variations of abundance percentage of individual body length of C.insidiosum
河蜾蠃蜚种群中个体体长变化如图15所示,体长0.038~0.2mm蜾蠃蜚的丰度百分比,第14天达到最大值后,27~90d内无显著性差异,第103天出现显著差异(P<0.05)并达到最小值。体长0.2~0.5mm蜾蠃蜚丰度百分比,第14天达到最小值后,至第78天出现显著差异(P<0.05),第90天达到最大值。体长0.5mm以上的蜾蠃蜚第64天达到最大值,随后快速下降至第90天出现显著差异(P<0.05)并达到最小值。

图15 河蜾蠃蜚个体体长丰度百分比的变化Fig.15 The variations of abundance percentage of individual body length of C.acherusicum
3 讨论
3.1 移植入日本对虾养殖池后钩虾和蜾蠃蜚群落结构的变动规律
日本对虾对钩虾和蜾蠃蜚的捕食量随着日本对虾体质量的增加而增大,而不同生长时期的日本对虾对饵料的摄食具有一定的选择性。Zupo等[19]研究表明,不同生长时期的日本对虾对饵料的选择性,主要是依据饵料生物的不同体长,而与饵料生物的营养成分无关。Reymond和Lagardère[20]对不同养殖阶段日本对虾的生长和摄食情况进行了研究,结果表明,日本对虾体长在2.3cm以下时,摄食的饵料主要为猛水蚤,其体长为0.05~0.80mm,约为此时日本对虾体长的1/20~1/30;当日本对虾体长在2.3~4.8cm时,摄食的饵料主要为摇蚊幼虫,其体长为1.0~6.2mm,约为此时日本对虾体长的1/23~1/8;当日本对虾生长至4.8cm以上时,摄食的饵料主要为大型底栖动物,体长在8.0mm以上,约为此时日本对虾体长的1/6。本实验中,第一期日本对虾生长至(1.60±0.25)cm时,体长0.2mm以下的钩虾和蜾蠃蜚丰度变化不明显,此时,日本对虾对钩虾和蜾蠃蜚的摄食量与其群落自身的繁殖量基本相等;待日本对虾生长至(2.70±0.43)cm时,体长0.2~0.5mm的钩虾和体长0.5mm以上的蜾蠃蜚丰度出现显著下降,此时钩虾和蜾蠃蜚的体长约为日本对虾体长的1/16~1/11;当日本对虾生长至(5.32±0.60)cm时,体长0.05mm以上的钩虾出现明显的下降,此时钩虾体长约为日本对虾体长的1/9。因此,处于不同生长时期的日本对虾所摄食的饵料体长比,随着日本对虾的生长而增大,本文进一步验证了Reymond和Lagardère的观点。
对于绝大多数钩虾亚目的种类,其繁殖模式可以概括为连续抱卵的多次生殖。郑新庆[21]报道了温度对强壮藻钩虾生长和繁殖的影响,15℃时强壮藻钩虾的繁殖节律最高(14.7±2.1)d。王超等[22-23]详细描述了温度对河蜾蠃生长繁殖的影响,实验室恒温培养河蜾蠃蜚F1代雌性个体可连续5次孵化幼体,雌体平均产幼体数量为22~32只,繁殖节律为12~14d。对于隐居蜾蠃蜚的性成熟时间,不同文献表述不一,初夏时为25d;水温30~34℃时,性成熟时间为8.5~20d。多篇文献对隐居蜾蠃蜚的胚胎发育时间进行了记载,Sheader[24]指出,15℃时,隐居蜾蠃蜚的胚胎发育时间仅为11d。Nair和Anger[25]指出,10、15和20℃条件下,隐居蜾蠃蜚的胚胎发育时间分别为13、9和8d。本实验研究表明,钩虾自移入养殖池,至第30天达到丰度高峰,由上文提供的数据计算,钩虾在此期间繁殖约2~3代;随后生物量和丰度均出现快速下降,此时日本对虾主要的捕食对象是体长0.5mm以上的钩虾个体,这部分钩虾无法大量繁殖便被摄食。蜾蠃蜚自移入养殖池,至第14天出现第1个丰度高峰,第64天出现第2个丰度高峰,由上文提供的数据计算,蜾蠃蜚约繁殖了3~4代。移入第14天时,日本对虾主要摄食体长0.2mm以下的蜾蠃蜚,此阶段成体蜾蠃蜚不断繁殖,雌性成体抱卵率可达43%,经笔者观察,隐居蜾蠃蜚雌性成体平均产幼体数量为18~45只,因此,此阶段蜾蠃蜚雌体的新生幼体个数应明显大于被摄食个数,产生第1个丰度高峰。当蜾蠃蜚移入64d时,蜾蠃蜚的生物量缓慢下降,丰度不断上升,这一现象表明,日本对虾此阶段主要捕食体长0.5mm以上的蜾蠃蜚个体,造成生物量下降;而这一部分蜾蠃蜚个体不断繁殖,并且繁殖出的新生个体数应大于被摄食的个体数,因此出现第2个丰度高峰。同时,通过对钩虾和蜾蠃蜚的丰度变化与其雌体抱卵率变化进行相关性分析可知,钩虾丰度与雌体抱卵率的相关系数为0.31,蜾蠃蜚的相关系数仅为0.25。由上述分析表明,钩虾和蜾蠃蜚的丰度高峰均与繁殖活动有关,但其丰度高峰个数低于笔者计算得到的繁殖代数,造成这一原因除了处于不同生长时期的日本对虾对钩虾和蜾蠃蜚的摄食压力不同外,还应与钩虾和蜾蠃蜚群落内雌性成体繁殖的不同步性有关。
3.2 移植入日本对虾养殖池钩虾和蜾蠃蜚群落结构的调控与养护
对钩虾和蜾蠃蜚群落在虾池内的调控,应主要包括以下3个方面:第一,确定日本对虾与饵料生物合理的密度比例,促进虾池养殖系统能流的良性循环;第二,确定调控措施,明确养护对象,设计合理的养护方案;第三,及时补充摄食饵料,为钩虾和蜾蠃蜚营造适宜的生存条件。
张士华等[12]报道,向养殖密度为10.5万尾/hm2的日本对虾养殖池移植入钩虾和蜾蠃蜚等饵料生物,合计160kg,取得了良好的养殖效果,按林德曼营养级定律[26]计算,上述饵料生物所提供直接的对虾产量可达255kg/hm2,并且待第一期对虾收获后至放养第137天,钩虾和蜾蠃蜚会达到第2个繁殖高峰期,但丰度均显著低于第1个峰值。本实验中,钩虾和蜾蠃蜚所提供的对虾产量可达106kg/hm2,并且蜾蠃蜚在放养第64天时,也出现第2个丰度高峰。上述2实验中,日本对虾与饵料生物的放养比例较为适宜,使饵料生物获得再次生长繁殖的机会,因此在该养殖条件下,日本对虾的放苗量宜控制在10.5~15万尾/hm2,钩虾和蜾蠃蜚的移入量宜控制在50~160kg/hm2。日本对虾放苗密度与饵料生物的移入量的比例应适宜,日本对虾的密度过大,可能使饵料生物还未充分繁殖,就被完全摄食,最终影响对虾的产量;饵料生物的密度过高,虽可以满足日本对虾的摄食需求,但对虾收获后,饵料生物剩余较多,造成浪费。
对钩虾和蜾蠃蜚群落的调控,应兼顾不同种的繁殖和生长周期,合理安排不同种个体引入的时间,从而达到理想的调控效果。本实验涉及2种钩虾和3种蜾蠃蜚,其中2种钩虾种间丰度差异明显,但繁殖时间有所不同,中华原钩虾在移入17~30d出现丰度高峰,而强壮藻钩虾在移入44~56d出现丰度高峰。对于体长0.5mm以上的钩虾个体,中华原钩虾在第44天达到最高,第67天出现显著差异;强壮藻钩虾在第17天达到最高,随后快速下降。因此,在本文涉及的养殖模式下,可以人为地调节两种钩虾种间的丰度组成,如在放养钩虾17d前,向养殖池中移入一定量已达到性成熟的中华原钩虾,使其充分繁殖,同时新生幼体不断生长,可以满足养殖67d后日本对虾对其大量摄食的需求。在放养钩虾44d前,移入一定量已达到性成熟的强壮藻钩虾,提高该种在钩虾群落中的比例,既有效地补充生物量,又有利于该种钩虾后期的繁殖。3种蜾蠃蜚自移入后不断繁殖,繁殖高峰出现的时间有所差异,河蜾蠃为第14天,隐居蜾蠃蜚为第78天,大蜾蠃蜚为第90天。对于体长0.5mm以上的蜾蠃蜚个体,河蜾蠃和大蜾蠃蜚变化趋势不明显,隐居蜾蠃蜚在第14天达到最高,第41天达到最低后,至第64天出现第二个高峰。因此,在日本对虾放养前,可以移入已达到性成熟的河蜾蠃成体,提高蜾蠃蜚群落中该种的丰度,使其充分繁殖。在第53天前,提高大蜾蠃蜚在蜾蠃蜚群落中的丰度,有利于其进行繁殖,同时,大蜾蠃蜚个体较大,经笔者测量,最大个体可达11.8mm,繁殖后可以供此阶段的日本对虾摄食。隐居蜾蠃蜚不同体长个体的变动规律性较强,可以在第41天前,移入一批已达到性成熟的隐居蜾蠃蜚,既保证其种群的生物量水平,又为后期其繁殖打下了基础。
在日本对虾养殖池中,主要的养护对象应为达到生物学最小型,并连续抱卵和繁殖的个体。郑新庆[21]报道,强壮藻钩虾的生物学最小型为8.01mm。Anger[27]表明,隐居蜾蠃蜚雌性达到性成熟的生物学最小型为1.87mm,抱卵雌体的生物学最小型为2.07mm。笔者通过室内实验观察,中华原钩虾的生物学最小型为6.94mm,大蜾蠃蜚的生物学最小型为2.53mm。本文涉及的钩虾和蜾蠃蜚0.5mm以上成体消失的顺序依次为,强壮藻钩虾67d,中华原钩虾81d,大蜾蠃蜚90d和隐居蜾蠃蜚103d。随着日本对虾对这些具有繁殖能力的钩虾和蜾蠃蜚成体的摄食,钩虾和蜾蠃蜚群落结构受到直接影响,群落无法维持生物量水平。因此,在日本对虾放苗前,可以采用网目为1.8~2.0 mm的网具,平铺于池底部,建立养护区。对虾无法进入养护区内捕食钩虾和蜾蠃蜚,而较小的钩虾和蜾蠃蜚则可自由游出,既满足对虾摄食,又可降低区内种群密度,待二期对虾放养后,且饵料生物达到一定生物量时,再撤除网具。
对于钩虾和蜾蠃蜚的摄食习性,国内外学者进行了一系列的研究。Zimmerman[28]以4种钩虾为实验对象,详细研究了其对大型海藻、藻类等附生植物、小颗粒碎屑等的选择性摄食,并指出附生藻类是对钩虾最佳的食物源。Pomeroy和Levings[29]对1种原钩虾(Eogammarus confervicolus)与河口藻类的关系进行了研究,实验表明,对该种原钩虾(E.confervicolus)生存和生长最好的藻类是缘管浒苔(Enteromorpha linza)。Cruz-Rivera和Hay[30]表明,藻钩虾(A.longimana)对13种海藻的摄食选择性与海藻的营养价值无关,藻钩虾明显更喜欢摄食网地藻和团扇藻。Nair和Anger[25]和Cunha等[31]对隐居蜾蠃蜚的生活史进行了系统的研究,结果表明,隐居蜾蠃蜚可以利用多种不同的食物源,对由轮虫、藻粉和碎屑等组成的混合食物有较好的摄食。Dahl[32]、Sheader[24]和Kevrekidis[33]指出,隐居蜾蠃蜚更偏好摄食碎屑、原生动物和底栖硅藻等。因此,在设置养护网具的同时,还应适时移入缘管浒苔、网地藻和团扇藻等大型藻类,以满足钩虾的摄食;着重培养底栖硅藻和原生动物等,以满足蜾蠃蜚的摄食需求,这将有利于钩虾和蜾蠃蜚的生长和繁殖,有效地延长其在虾池中存活时间。
综上所述,移植入虾池的钩虾和蜾蠃蜚,一方面为日本对虾提供良好的饵料,同时还可以摄食对虾的残饵,净化水质;另一方面,设置一定的养护器具,采用一定的调控措施,可以有效地维持其在日本对虾养殖池中的生物量水平,节约养殖成本。这必将在日本对虾土池养殖过程中,有着更为广泛的应用前景。
致谢:感谢中科院海洋所任先秋研究员在钩虾、蜾蠃蜚种类鉴定方面给予的悉心指导,感谢海洋生命学院于子山副教授在实验准备过程中给予的无私帮助。
[1] 任先秋.中国动物志,无脊椎动物,第四十一卷,端足目,钩虾亚目(一)[M].北京:科学出版社,2006:132-374.
[2] 黄宗国.中国海洋生物种类与分布[M].北京:海洋出版社,1994:532-533.
[3] 武云飞,吴翠珍,黄勇,等.一种新蛋白源——青海钩虾营养价值的初步研究[J].青岛海洋大学学报:自然科学版,1996,26(3):309-311.
[4] 郭焱,张人铭,蔡林钢,等.赛里木湖钩虾营养及渔业效应分析[J].淡水渔业,2002,32(4):52-54.
[5] 郑严,李茂堂,田凤琴.蜾蠃蜚在对虾养殖中的应用研究[J].海洋科学,1985(9):35.
[6] Birldund J.Biomass,growth and production of the amphipod Corophiurn insidiosum Crawford,and preliminary notes on Corophiurn volutator(Pallas)[J].Ophelia,1977,16:187-203.
[7] 王克行.解决大面积养虾饲料的一点设想[C].[s.l.]:全国海水养殖增殖发展途径学术会议论文报告汇编,1980:200-203.
[8] 韩方训,王道和,韩丰贵,等.沙蚕在对虾养殖生产中的应用[J].海洋科学,1990(3):4-6.
[9] 张志南,于子山,段榕琦,等.虾池纳潮期日本刺沙蚕幼虫数量及其沉降的研究[J].海洋与湖沼,1994,25(3):248-257.
[10] 周一兵.沙蚕移植在对虾养殖中的应用及生态效益[J].海洋学通报,1999,34(11):12-15.
[11] 邓锦松.投放双齿围沙蚕和毛蚶对虾池的生物修复作用[D].青岛:中国海洋大学水产学院,2006:13-32.
[12] 张士华,杨秀霞,林式柱,等.几种对虾在黄河三角洲地区生长及移植饵料生物效果的比较[J].青岛海洋大学学报:自然科学版,2002,32(4):543-550.
[13] 张庆文,孔杰,栾生,等.日本囊对虾亲虾人工繁育技术初步研究[J].海洋水产研究,2005,26(4):14-16.
[14] 刘文亮.长江河口大型底栖动物及其优势种探讨[D].上海:华东师范大学,2007:22-53.
[15] 闫启仑,韩明辅,陈红星,等.辽宁沿岸海洋底栖钩虾类的种类组成与分布[J].海洋环境科学,1998,17(1):26-29.
[16] 袁兴中,陆健健.围垦对长江口南岸底栖动物群落结构及多样性的影响[J].生态学报,2001,21(10):1643-1644.
[17] 钱迎倩,马克平.生物多样性的原理与方法[M].北京:中国科学技术出版社,2006:140-166.
[18] Pielou E C,Ecological diversity[M].New York:John wiley,1975.
[19] Zupoa V,Lumareb F,Bisignano V.Feeding of Penaeus japonicus Bate(Decapoda:Penaeidae)in pond cultures:size descriptors and food selection.[J].Journal of Natural History,1998,32(10-11):1813-1824
[20] Reymond H,Lagardère J P.Feeding rhythms and food of Penaeus japonicus Bate(Crustacea,Penaeidae)in salt marsh ponds:role of halophilic entomofauna[J].Aquaculture 1990,84(2):125-143.
[21] 郑新庆.端足类啃食作用对筼筜湖大型海藻群落影响的初步研究[D].厦门:厦门大学,2008:64-73.
[22] 王超,闫启仑,陈红星,等.端足类河蜾蠃蜚生活周期及其沉积物毒理敏感性研究[J].华东师范大学学报,2009,3:1-5.
[23] 王超,王睿睿,闫启仑,等.温度对底栖端足类河蜾蠃蜚(Corophium acherusicum)存活、生长和发育的影响[J].海洋环境科学,2009,28(2):138-141.
[24] Sheader M.Distribution and reproductive biology of Corophium insidiosum(Amphipoda)on the north-east coast of England[J].Mar Boil Ass UK,1978,58:585-596.
[25] Nair K K C,Anger K.Life cycle of Corophium insidiosum(Crustacea,Amphipoda)in laboratory culture[J].Helgoland Marine Research,1979,32(3):279-294.
[26] 孙儒泳.动物生态学原理[M].北京:北京师范大学出版社,2001:478-480.
[27] Anger K.Untersuchungen zum Lebenszyklus des Amphipoden Bathyporeia sarsi,Microdeutopus gryllotalpaund Corophium insidiosumin der Kieler Bucht[J].Mitteilungen ausdem Zoologischen Museum der Universita,1979,1:1-6.
[28] Zimmerman R,Gibson R,Harrington J.Herbivory and detritivory among gammaridean amphipods from a Florida seagrass community[J].Marine Biology,1979,54:41-47.
[29] Pomeroy W M,Levings C D.Association and Feeding Relationships Between Eogammarus confervicolus(Amphipoda,Gammaridae)and Benthic Algae on Sturgeon and Roberts Banks,Fraser River Estuary[J].Can J Fish,1980,37(1):1-10.
[30] Cruz-Rivera E,Hay M E.Can quantity replace quality?Food choice,compensatory feeding and fitness of marine mesograzers[J].Ecology,2000,81:201-219.
[31] Cunha M R Sorbe J C,Moreira M H.The amphipod Corophium multisetosumin Ria de Aveiro.I.Life history and aspects of reproductive biology[J].Marine Biology,2000,137:637-650.
[32] Dahl E.Ecological range of Baltic and North Sea species[J].Oikos(Suppl.),1973,15:85-90.
[33] Theodoros Kevrekidis.Population dynamics,growth and reproduction of Corophium insidiosum(Crustacea:Amphipoda)at low salinities in Monolimni lagoon(Evros Delta,North Aegean Sea)[J].Hydrobiologia,2004(522):117-132.
Studies on the Variations of Gammarid and Corophid(Amphipoda)Communities in Shrimp Marsupenaeus japonicus Farm Pond
ZHONG Yuan,MA Shen
(The Key Laboratory of Mariculture,Ministry of Education,Ocean University of China,Qingdao 266003,China)
During April 2009to July,the Gammarid and Corophid amphipods were transplanted into three shrimp Penaeus japonicus farm ponds and the variation of amphipod communities was investigated.Two gammarid species were identified as Eogammarus sinensis Ren and Ampithoe valida Smith,and three corophid species as Corophium major Ren,C.major Ren,and C.insidiosumCrowford.Among the gammarid species E.sinensis Ren was predominant and the dominant corophid species were alternating obviously.During 53days after stocking,Corophium major Ren was predominant,and after 78days,Corophium insidiosum Crawford was predominant.There was a significant difference in the Shannon-Weaver diversity index(H′)and evenness(J)at 67th day after stocking in gammrid community,with the maximum index at the beginning.There was a significant difference in the Shannon-Weaver diversity index(H′)and evenness(J)at 90th day after stocking in corophid community,with the maximum index at 64th day.The biomass and density of gammird community were highest at 30th day after stocking,and then declined rapidly.The biomass and density of corophid community were highest at 14th day after stocking,and there was a second peak of density at 64th day.Meanwhile,there were significant differences in abundance percentage of different body length between gammrid community and corophid community,and some cyclical variation was showed.Thus two types of reasons that caused community variable were analyzed,and the control scheme of gammrid and corophid community was proposed.
Eogammarus;Ampithoe Corophium;community;variation;shrimp farm pond;shrimp farm poad
S917
A
1672-5174(2012)1-2-059-08
公益性行业(农业)科研专项(201103034)资助
2011-02-09;
2011-04-20
钟 源(1985-),男,硕士生。E-mail:albert.128@163.com
**通迅作者:E-mail:mashen@ouc.edu.cn
责任编辑 王 莉
