黄土高原典型草原草地根冠比的季节动态及其影响因素
2012-01-02李旭东张春平傅华
李旭东,张春平,傅华
(兰州大学草地农业科技学院 草地农业生态系统国家重点实验室,甘肃 兰州730020)
地上生物量(shoot)和地下生物量(root)是陆地生态系统碳储量的重要组成部分。有研究表明,CO2浓度升高会促进植物的光合产物向根系分配,从而提高陆地生态系统地下部分的碳素固定[1],这可能是全球“碳失汇”中的一个重要的汇[2]。生态学家已经认识到地下部分是陆地生态系统碳分配与碳过程的核心环节,而根系是联系地上和地下过程的主要纽带[3]。植物的干物质在地上、地下的分配是一个复杂的过程,它受到植物种类、年龄、气候条件等众多因素的影响[4]。相对于地上生物量而言,清楚地了解地下生物量及其所占比率还存在很大局限性[5],主要是由于地下生物量的测定尚存在方法上的缺陷,而且不同方法得出的结果有较大的差异[6,7]。为了更好地估算地下生物量,建立地上地下生物量的关系是一种常用的方法,而根冠比(即植物地下生物量和地上生物量的比值,R/S)则是最常用的一个参数[8],被众多陆地生态系统模型作为计算光合产物向植物地下部分分配的依据,如 HRBM[9],SLAVE[10],CASA[11],CARAIB[12],DEMETER[13]等模型。另外,利用根冠比,可以通过植物地上部分生物量估算景观、区域尺度上的植被碳储量。因此,确定根冠比的季节动态及其影响因素,可以更加精确地估算地下生物量和地下碳储量,对提高陆地生态系统模型的模拟精度以及生态系统碳储量估算的准确性具有十分重要的意义。
草地作为陆地生态系统的重要组成部分,约占陆地总面积的1/5,在陆地生态系统碳循环中占有十分重要的地位[4]。草地生态系统中净初级生产力的60%~90%分配于地下,地下生物量通常超过地上生物量2~5倍[14]。通过根系脱落物输入到土壤中的碳也远远超过了地上凋落物的碳输入量。所以,精确的计算草地地下生物量具有重要的生态意义。但草地生态系统植被碳储量的估算还存在很大的不确定性,其中一个重要的原因就在于对植物地下部分认识的不足[5]及其测定方法存在缺陷[15,16]。因此,本研究以黄土高原典型草原为研究对象,于2007-2009年连续3年进行了地上、地下生物量的测定,旨在探讨:1)黄土高原典型草原植被根冠比及其季节动态;2)放牧与围封对植被根冠比的影响;3)根冠比与降水量和气温的关系,从而为精确预测地下生物量提供依据,并且为陆地生态系统碳模型提供参考。
1 材料与方法
1.1 研究区概况
试验地设在兰州大学半干旱气候与环境观测站及周围区域。该站位于甘肃省榆中县北部夏官营镇,地理坐标为北纬35°57′,东经104°09′,海拔1 966m。地貌为黄土高原残塬梁峁沟壑,属大陆性半干旱气候,年平均气温6.7℃,年降水量382mm,蒸发量1 343mm,无霜期90~140d。年日照时数约2 600h。该区域塬面原为传统的耕作农田,20世纪80年代中期弃耕,已基本恢复到天然植被。植被类型为半干旱典型草原,草地植物主要有长芒草(Stipa bungeana)、赖草(Leymus secalinus)、铁杆蒿(Tripolium vulgare)和阿尔泰狗娃花(Heteropappus altaicus)等。土壤为灰钙土[17]。
1.2 研究方法
本研究设置围封与放牧2个处理。围封样地(ungrazed grassland,UG)在气候观测站内,2005年10月设立围栏,面积约为8hm2。围栏外为放牧样地(grazed grassland,GG),放牧强度为10-3月:1~2羊单位/hm2;4-9月:2~3.5羊单位/hm2。
在围封与放牧样地中,选择坡向和地势平缓一致的地段各设置3个50m×50m的重复样地。围封和放牧样地地上、地下生物量的测定于生长季(3-11月)各月中下旬进行。地上生物量采用收获法,每个重复样地中随机设置6个1m×1m的样方,将样方内植物齐地面刈割,带回实验室于65℃条件下烘干并称重(精确至0.01 g)。地下生物量采用根钻法测定(根钻直径为9.3cm)。在测定地上生物量后的18个样方内,用根钻分层取土至100cm土层,每10cm为一层。所取样品经干筛法(孔径0.5mm)获取根样后用清水洗净,于65℃烘干并称重(精确至0.01g),计算地下生物量。降水、气温等气象资料从兰州大学半干旱气候与环境观测站获取[17]。
1.3 数据处理方法
根冠比(R/S)=地下生物量(root)/地上现存生物量(shoot)
使用Excel 2003进行数据处理,应用统计分析软件SPSS 17.0的Correlate和Regression进行相关分析和回归分析。根冠比与气温和降水量进行相关性分析时,与各月的根冠比相对应的是上月的降水量(P)和当月气温的平均值(T)。
2 结果与分析
2.1 根冠比的季节动态
3年的试验结果表明,放牧与围封草地根冠比的季节变化均呈现反抛物线型(图1)。围封样地在3月牧草返青之前根冠比最大,随着植物的生长开始降低,到9月最小,之后开始上升,各月之间存在显著差异(P<0.05)。放牧样地最大值出现在11月,最小值出现在9月,各月之间也存在显著差异(P<0.05),放牧样地3年的根冠比平均值为3.42,围封样地为2.75。2个样地各个月份根冠比的年际间差异都不显著。
2.2 围封与放牧对根冠比的影响
方差分析结果表明,2007,2008和2009年放牧样地的根冠比在不同生长季都显著高于围封样地(P<0.05),其平均值放牧样地分别为3.13,3.36和3.77,围封样地分别为2.55,2.56和3.13(表1)。不同生长季根冠比的变异系数放牧样地3年分别是36.7%,32.3%,49.2%,均高于围封样地的28.2%,26.3%,26.1%(表1)。
2.3 气温和降水量对根冠比的影响
将各月的根冠比与上月的降水量(P)进行相关性分析。结果表明,根冠比与上月降水量之间存在极显著的负相关关系(n=27,P<0.001)(图2)。
将各月的根冠比与气温(T,当月气温平均值)进行相关性分析表明,根冠比与当月气温之间也存在着极显著的负相关关系(n=27,P<0.001)(图3)。
通过对根冠比与气温、降水量进行逐步回归分析,分别得出围封与放牧样地根冠比与气温、降水量的多元回归方程(表2)。对围封样地而言,降水量和气温的影响对根冠比的贡献率分别为55%和45%,而放牧样地则分别为38%和62%。
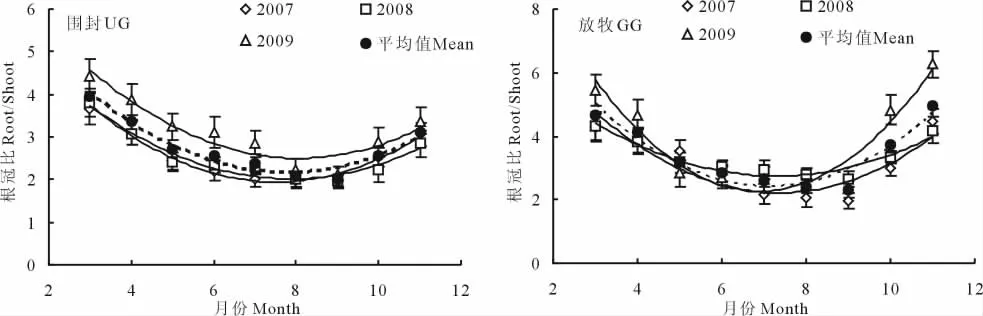
图1 围封与放牧草地根冠比(R/S)的季节变化Fig.1 The seasonal dynamics of the R/S in grazed grassland(GG)and ungrazed grassland(UG)

表1 围封与放牧草地根冠比的统计参数Table 1 Statistical parameters of the R/S in grazed grassland(GG)and ungrazed grassland(UG)

图2 围封、放牧草地R/S与降水量的关系Fig.2 The relationships between R/S and precipitation in grazed and ungrazed grassland
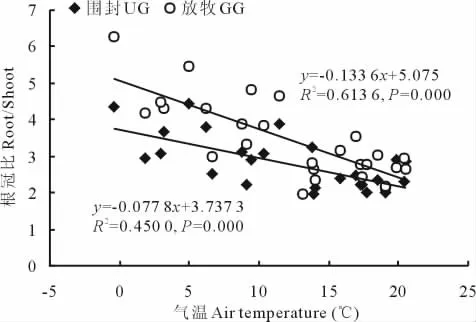
图3 围封、放牧草地R/S与气温的关系Fig.3 The relationships between R/S and air temperature in grazed and ungrazed grassland

表2 围封、放牧草地R/S与气温、降水量的回归方程Table 2 Regression equations between R/S and air temperature and precipitation
3 讨论
根冠比作为重要的参数之一,在全球陆地生态系统碳模型中具有重要的意义,其有助于精确估算全球生态系统的碳分配和碳固存[4]。
本研究结果显示,黄土高原典型草原植物的根冠比呈反抛物线型季节变化,生长季初和生长季末根冠比值较大。3月,草地的植物还没有返青,植物将大部分物质都储存在根系中,所以,3月份的根冠比较高。返青后,随着植物地上部分快速生长,光合产物向地上部分转移,导致根冠比下降,到9月份达到最低值。9-11月随着温度的降低,植物地上部分开始枯死,植物为了第2年的再生和适应环境的变化而将能量转运至根系中,导致根冠比上升。2007,2008和2009年围封样地根冠比变化范围分别为1.96~3.67,1.98~3.79和2.13~4.44;放牧样地分别为1.95~4.47,2.65~4.67和2.04~6.26。白永飞等[18]对内蒙古羊草(Leymus chinensis)群落研究表明,生长季内根冠比的变幅为3.26~210.62;耿浩林等[4]对恢复羊草草地的研究结果为22.3~41.5和10.4~39.7。与之相比,本研究连续3年的根冠比的变幅(1.95~6.26)均较小,但与几个黄土高原典型草地的根冠比结果(均值为1.99,2.33,2.86)基本一致[19]。张娜和梁一民[20]研究表明黄土丘陵区天然草地群落根冠比的季节变化也呈反抛物线型,本研究结果与此一致,但其根冠比的均值5.65要高于本研究结果,可能主要因为其测定年份极干旱,造成根冠比结果较高。
许多研究表明,植物在不同环境条件下的资源分配格局反映了植物生长对环境的响应规律和资源分配对策[21-25]。放牧能够改变牧草各器官之间固有的物质与能量分配模式。在本研究中,放牧样地的根冠比显著高于围封样地,这与李金花和李镇清[26]对放牧11年后的典型草地的研究结果一致。放牧样地由于家畜的采食,地上生物量显著低于围封样地,可能是造成放牧样地根冠比较高的主要原因;但与Jatimliansky等[27]放牧降低根冠比的结果不相同,原因是本研究在连续放牧草地进行,放牧草地地上部分不能获得充分生长的机会,降幅常常高于地下部分,造成根冠比增加。另外,放牧样地根冠比的变异也大于围封样地,原因是由于家畜的采食作用导致放牧样地的地上生物量的变异增大,进而使得根冠比的变异增大[26]。同时,放牧会抑制其他适口性好的植物的生长,从而改变草地的根冠比[28]。
在自然环境中,环境因子的胁迫作用往往限制了植物生物量分配格局,植物对光、养分、水分的竞争是决定生物量分配的重要因素[29,30]。植物生物量和水分的关系尤其密切,在干旱和半干旱区,水分是生物量形成的主要限制因子[31]。天然草地地上、地下生物量主要受降水量和温度的影响[32]。由于草地群落生物量相对于水热条件具有时滞效应,因此,一定时期的生物量必定与此前的水热条件有关[4]。为此,本研究分析了前期水热因素(包括上月降水量之和、当月平均气温)对根冠比的影响,结果显示根冠比与降水量和温度的相关性极显著。这与Coupland[33]对北美草原12个长期生态站资料的分析中发现根冠比与干旱的程度及温度有关的结论一致;另外,其他一些研究也表明根冠比随着降水量的增加而降低[34-37]。根冠比同降水量的相关性可以用以下假说解析:随着植物可获得水分的增加,植物生长对水分的要求降低,地上部分生长要强于地下部分,于是根冠比降低[35-37]。另外,气温的升高使得土壤温度升高,进而促进了植物根系对营养和水分的吸收,从而降低了植物对地下生物量的投入[38],导致根冠比降低。对围封样地而言,降水量对根冠比的影响更大,主要是由于半干旱地区水分是调控植物生物量分配的主要因子,降水对其地上和地下生物量分配的影响要大于气温。而放牧样地由于动物采食、践踏等其他因素的影响,水分和温度对根冠比的影响发生了变化,其综合的影响尚需进一步研究。
不同生态系统的地上-地下生物量分配格局主要受气候因素的影响[39]。因此,根冠比是可预测的[40]。本研究根据各月根冠比与上月降水量、当月平均气温之间建立了拟合度良好的回归方程,可以用来模拟不同季节根冠比的动态变化并估算地下生物量。
致谢:感谢文海燕、安卓、杨益和滕泽琴等在野外实验中所付出的辛勤劳动;感谢兰州大学半干旱气候与环境观测站提供气象资料。
[1]Hungate B A,Holland E A,Jackson R B,et al.The fate of carbon in grasslands under carbon dioxide enrichment[J].Nature,1997,388:576-579.
[2]Woodwel G M,Whittacker R H,Reiners W A,et al.The biota and the world carbon budget[J].Science,1978,199:141-146.
[3]贺金生,王政权,方精云.全球变化下的地下生态学:问题与展望[J].科学通报,2004,49(13):1226-1236.
[4]耿浩林,王玉辉,王风玉,等.恢复状态下羊草(Leymus chinensis)草原植被根冠比动态及影响因子[J].生态学报,2008,28(10):122-128.
[5]McNaughton S J,Banyikwa F F,McNaughton M M.Root biomass and productivity in a grazing ecosystem:the Seren-geti[J].Ecology,1998,79:587-592.
[6]Vogt K A,Vogt D J,Palmiotto P A,et al.Review of root dynamics in forest ecosystems grouped by climate,climatic forest type and species[J].Plant and Soil,1996,187:159-219.
[7]Titlyanova A A,Romanova I P,Kosykh N P,et al.Pattern and process in above-ground and below-ground components of grassland ecosystems[J].Journal of Vegetation Science,1999,10:307-320.
[8]Mokany K,Raison R J,Prokushkin A S.Critical analysis of root∶shoot ratios in terrestrial biomes[J].Global Change Biology,2006,12:84-96.
[9]Esser G.Sensitivity of global carbon pools and fluxes to human and potential climatic impacts[J].Tellus,1987,39:245-260.
[10]Friedlingstein P,Delire C,Muller J F,et al.The climate induced variation of the continental biosphere:a model simulation of the Last Glacial Maximum[J].Geophysical Research Letters,1992,19:897-900.
[11]Potter C S,Randerson J T,Field C B,et al.Terrestrial ecosystem production:process model based on global satellite and surface data[J].Global Biogeochemical Cycle,1993,7:811-841.
[12]Wamant P,Francois L,Strivay D,et al.CARAIB:global model of terrestrial biological productivity[J].Global Biogeochemical Cycles,1994,8:255-270.
[13]Foley J A.Net primary productivity of the terrestrial biosphere:The application of a global model[J].Journal of Geophysical Research,1994,99:20773-20783.
[14]Speidel B.Primary production and root activity of a golden oat meadow with different fertilizer treatments[J].Polish Ecological Studies,1976,2:77-89.
[15]朱桂林,韦文珊,张淑敏,等.植物地下生物量测定方法概述及新技术介绍[J].中国草地学报,2008,(3):93-99.
[16]Lauenroth W K.Methods of estimating belowground net primary production[A].In:Sala O E.Methods in Ecosystem Science[M].New York:Springer,2000:58-71.
[17]Li X D,Fu H,Li X D,et al.Effects of land-use regimes on carbon sequestration in the Loess Plateau,Northern China[J].New Zealand Journal of Agricultural Research,2008,51:45-52.
[18]白永飞,许志信,李德新.羊草草原群落生物量动态研究[J].中国草地,1994,3:1-5,9.
[19]中国科学院内蒙古草原生态系统定位站.草原生态系统研究-I[C].北京:科学出版社,1985.
[20]张娜,梁一民.黄土丘陵区天然草地地下/地上生物量的研究[J].草业学报,2002,11(2):72-78.
[21]Schmid B.Some ecological and evolutionary consequences of modular organization and clonal growth in plants[J].Evolutionary Trends in Plants,1990,4:25-34.
[22]Watson M A,Geber M A,Jones C S.Ontogenetic contingency and the expression of plant plasticity[J].Trends in Evolution and Ecology,1995,10:474-475.
[23]Geber M A,Watson M A,Kroon H de.Development and resource allocation in perennial plants:the significance of organ preformation[A].In:Bazzaz F A,Grace J.Plant Resource Allocation[M].New York:Academic Press,1997.
[24]Geber M A,Watson M A,Kroon H de.Morphological plasticity in clonal plants:the foraging concept reconsidered[J].Journal of Ecology,1997,83:143-152.
[25]Smith N S.Variation in response to defoliation between population of Bouteloua curtipendula var.caespitosa with different livestock grazing histories[J].American Journal of Botany,1998,85:1266-1272.
[26]李金花,李镇清.不同放牧强度下冷蒿、星毛委陵菜的形态可塑性及生物量分配格局[J].植物生态学报,2002,26(4):435-440.
[27]Jatimliansky J R,Gimenez D O,Bujan A.Herbage yield,tiller number and root system activity after defoliation of prairie grass(Bromus cathartics Vahl)[J].Grass and Forage Science,1997,52:52-62.
[28]汪诗平,李永宏.内蒙古典型草原退化机理的研究[J].应用生态学报,1999,10:437-441.
[29]Geiger D R,Servaites J C.Carbon allocation and response to stress[A].In:Mooney H A,Winner W E,Pell E J.Response of Plants to Multiple Stresses[M].CA,San Diego:Academic Press,1991:103-127.
[30]Mooney H A,Winner W E.Partitioning response of plants to stress[A].In:Mooney H A,Winner W E,Pell E J.Response of Plants to Multiple Stresses[M].CA,San Diego:Academic Press,1991:129-141.
[31]成文联,柳海鹰,王世东.生物量与其影响因素之间关系的研究[J].内蒙古大学学报(自然科学版),2000,31(3):285-289.
[32]董全民,赵新全,马玉寿,等.牦牛放牧率与小嵩草高寒草甸暖季草地地上、地下生物量相关分析[J].草业科学,2005,22(5):65-71.
[33]Coupland R T.Grassland Ecosystems of the World:Analysis of Grasslands and their Uses[M].Cambridge:Cambridge University Press,1979:62.
[34]Wang R Z,Gao Q,Chen Q S.Effects of climatic change on biomass and biomass allocation in Leymus chinensis(Poaceae)along the Northeast China Transect(NECT)[J].Journal of Arid Environments,2003,54:653-665.
[35]Gower S T,Vogt K A,Grier C C.Carbon dynamics of Rocky Mountain Douglas-fir:influence of water and nutrient avail-ability[J].Ecological Monographs,1992,62:43-65.
[36]Brand B M.Quantifying biomass and carbon sequestration of plantation blue gums in South West Western Australia[D].Curtin University of Technology,1999.
[37]Schenk H J,Jackson R B.Rooting depths,lateral root spreads and below-ground/above-ground allometries of plants in water-limited ecosystems[J].Journal of Ecology,2002,90:480-494.
[38]Garkoti S C,Singh S P.Variation in net primary productivity and biomass of forests in the high mountains of Central Himalaya[J].Journal of Vegetation Science,1995,6:23-28.
[39]Hui D,Jackson R B.Geographical and interannual variability in biomass partitioning in grassland ecosystems:A synthesis of field data[J].New Phytologist,2005,169(1):85-93.
[40]Friedlingstein P,Joel G,Field C B,et al.Toward an allocation scheme for global terrestrial carbon models[J].Global Change Biology,1999,5:755-770.
