松嫩平原不同生境芦苇生殖分株的异速生长分析
2012-01-02刘秀香杨允菲
刘秀香,杨允菲
(东北师范大学草地科学研究所 植被生态科学教育部重点实验室,吉林 长春130024)
异速生长是生物体各部分器官的不均匀和不成比例的生长,用于研究生长中的整体与部分,或部分与部分之间的关系问题,动植物异速生长的定量关系大多数都可以用幂函数较好的表达。异速生长是生物体生长的一种特性,受遗传物质所决定,同时也受外界条件的影响[1]。目前,关于异速生长问题的研究已有很多[2-8]。
芦苇(Phragmites australis),根茎型无性系禾草,营养繁殖能力强,天然种群主要以根茎繁殖补充更新,其生态适应性较广,是世界广布种,生长于池沼、河岸、河口、湖滨、荒漠、盐碱区等地,在适宜环境下常形成单优种群落[9,10]。在松嫩平原上,芦苇常以单优种群落或主要伴生种的形式存在[11]。在长期适应不同生态环境的过程中,芦苇形成了不同的生态型,具有较大的生态可塑性[12]。芦苇具有较高的经济和生态价值,近几年来,许多专家学者围绕着芦苇的分类、资源分布、生态功能、生理学、形态学和芦苇湿地的生态学效应等方面做了大量研究[13-18],并取得了丰硕成果。本研究根据实地调查,对松嫩平原季节性积水的单优种芦苇群落和旱地生境羊草(Leymus chinensis)与芦苇混生群落2种生境芦苇种群的生殖分株数量特征进行定量分析,以揭示芦苇种群生殖分株的生长和物质分配策略,为进一步探讨芦苇的生长对策和植物资源的有效利用提供一定的理论依据。
1 材料与方法
1.1 研究区自然概况
研究区位于松嫩平原南部、吉林省长岭县种马场、东北师范大学草地生态研究站(44°38′N,123°41′E)。该区属温带半湿润、半干旱季风型气候,平均海拔为141.5m,春季干旱多风,夏季炎热多雨,平均降水量约为449.5 mm,主要集中在6-9月,年蒸发量为年降水量的3~4倍。年平均气温4.6~6.4℃,年积温3 000~3 500℃。松嫩平原主要植被类型为羊草草甸草原,主要伴生种有芦苇、星星草(Puccinellia tenuiflora)和蒿属(Artemisia)类植物,土壤基本为碱化草甸土[19-21]。
1.2 取样方法
本研究选择2种旱地生境作为研究对象。生境1(H1):单优种芦苇群落,土壤为碱化草甸土,每年7-8月份常有积水。生境2(H2):羊草+芦苇群落,土壤为碱化草甸土,很少积水或积水时间很短。
2010年9月末对7月底至8月末积水20d左右和只有很少积水的H1、H2两生境芦苇生殖分株分别取样,设定3个10~15m2的样地,每个样地随机抽取20个分株左右,齐地面剪下,取样时注意保持各分株的完整性,将样本带回实验室,将3个样地所取样品充分混合后再随机抽取30株,测量各生殖分株的株高和花序长,并将花序、茎、叶各部分分置,在烘箱内80℃左右烘干至恒重,以各构件为单位分株称重。
1.3 数据处理
根据测量的数据,对各生境数量指标做统计分析,最大值和最小值反映样本的大小范围,平均数(M)代表样本各性状的整体水平,标准差(SD)反映绝对变异度,变异系数(CV)反映相对变异度。用花序长度占生殖分株高度的百分比作为生殖生长比率的数量指标,用花序生物量占分株总生物量的百分比作为生殖分配的数量指标,用叶生物量占分株总生物量的百分比作为生产分配的数量指标,用茎生物量占分株总生物量的百分比作为茎生物量分配的数量指标,计算公式分别为:

对各性状间的关系建立定量化描述模型,所有定量关系均采用经典的幂函数Y=βXα异速曲线进行描述,将上述方程经对数转换,得到如下方程:lgY=lgβ+αlgX,式中,β为标准化常数,α为异速指数[22,23],分析转换后的线性相关关系。

表1 2种生境芦苇种群生殖分株的数量特征(N=30)Table 1 Quantitative character in reproductive ramets of P.australis populations under two different habitats
2 结果与分析
2.1 构件的数量特征
单优种芦苇群落(H1)和羊草+芦苇群落(H2)除花序长和花序生物量间无显著差异外(表1),其他性状间均达到显著水平,具有较大的生态可塑性。绝对数量性状中,2种生境变异系数以花序生物量的变异系数为最高,达57.73%和49.87%,各性状变异系数总体上为生境1>生境2,单优种芦苇群落各性状变异系数大于羊草与芦苇混生群落,表明2种异质生境中,在构件水平上,花序生物量的变异度高于其他性状的变异度;整体水平上,单优种芦苇群落的变异度高于羊草与芦苇混生群落的变异度。2种生境的生物量分配在叶、茎和花序上均达到显著水平,其中叶和花序达到极显著水平。叶是植物的营养器官,可以产生有机质供个体生长发育之需,2种生境芦苇生殖分株生物量都是优先分配叶器官,把较多的物质分配给叶的建造上,以保证有机体供应,适应不利的生态环境。各相对数量性状的变异系数以生殖分配为最高,为36.17%和39.44%,表明不同生境下芦苇生殖分株各构件生物量分配有一定差异,但在整体水平上具有一定的相似性,在各组分中,花序的生态可塑性高于其他构件的生态可塑性。既体现了芦苇生殖分株对环境的适应能力,也蕴含了其生殖分配策略[11]。
2.2 生殖生长与株高的关系
生殖分株花序的伸长属于生殖生长,其生殖生长量可用绝对数量指标花序长和相对数量指标生殖生长比率来衡量。经相关分析,2种生境生殖株花序长对数和株高对数呈极显著线性正相关,即花序长和株高呈极显著幂函数正相关,而生殖生长比率与株高呈显著的幂函数负相关关系(图1),表明生殖株花序长度是随着植株高度的增加而增加,而生殖分株生殖生长比率的降低表明分株将更多的能量用于茎的伸长以提高较差环境条件下的现实竞争力[24]。反映了异质生境芦苇自我调节的生殖分配策略及对异质生境较强的适应能力[11]。
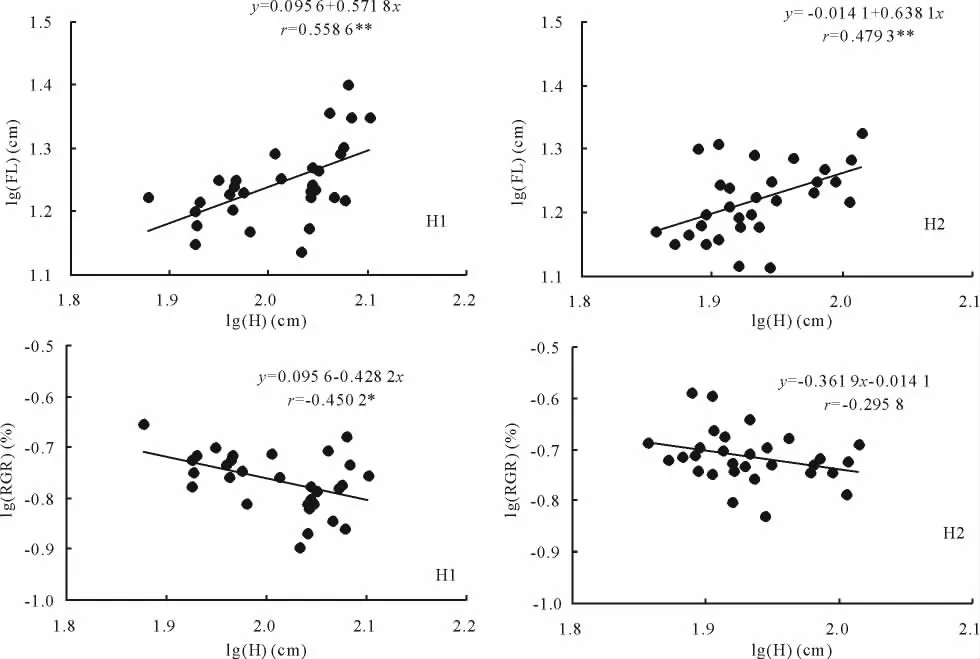
图1 不同生境芦苇种群生殖分株花序长对数[lg(FL)]/生殖生长比率对数[lg(RGR)]与株高对数[lg(H)]的关系Fig.1 Relationship between logarithm of flower length[lg(FL)]/logarithm of ratio of reproductive growth[lg(RGR)]and logarithm of height[lg(H)]of reproductive ramets in P.australis populations under different habitats
2.3 构件生物量分配之间的关系
经统计分析,2种生境向花序的生殖分配随向叶片的生产分配的增加呈极显著的幂函数负相关关系,其异速指数α均大于1;生产分配随茎生物量分配的增加呈极显著的幂函数负相关关系,其异速指数α约为1(图2),表明随生产分配的增加,生殖分配减少的速度加快,茎生物量分配增加的速率与生产分配降低的速率几乎相等。由此反映出芦苇种群生殖分株的生长与分配对策,在植物的生长发育过程中,对每一株植物来说首先要在激烈的竞争中生存下来,生殖处于第2位[25]。在外界条件较恶劣的环境中,个体首先将更多的能量用于植株生长,以获取更多的空间和资源,提高现实竞争力,再将能量用于个体的繁殖[26]。生殖分配的增加是以减少其他构件的生物量分配为代价的。构件间的这种生物量分配关系,蕴含了芦苇生殖分株生长与生殖策略。
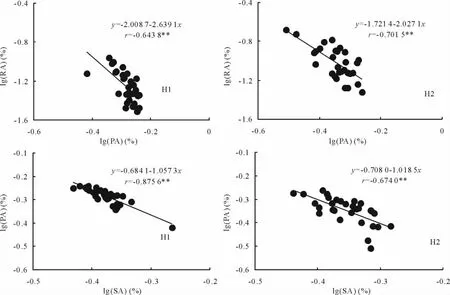
图2 不同生境芦苇种群生殖分株的生殖分配对数[lg(RA)]与生产分配对数[lg(PA)]和生产分配对数[lg(PA)]与茎生物量分配对数[lg(SA)]之间的异速生长关系Fig.2 Allometry relationships on the logarithm of reproductive allocation[lg(RA)]and logarithm of productive allocation[lg(PA)]/logarithm of productive allocation[lg(PA)]and logarithm of stem biomass allocation[lg(SA)]of reproductive ramets in P.australis populations under different habitats
3 结论
2种生境芦苇种群生殖分株各数量性状既具有较大的差异性,也具有相同的分配与变异特征。其中,分株高、分株生物量、生殖生长比率和构件生物量分配均以雨季积水时间较长的单优种芦苇群落显著高于几乎无积水发生的羊草+芦苇群落;在2种异质生境分株生物量分配中均以叶的生长分配最大,以及各数量性状均以花序生物量的变异系数最大,表现出花序生物量的可塑性最大。
2种生境芦苇种群生殖分株的花序生长与株高之间具有相同的异速生长规律。其中,花序长与株高之间呈幂函数正相关关系,生殖生长比率与株高之间呈幂函数负相关关系,表现出随株高的增加花序生长相对减少。
2种生境芦苇种群生殖分株各构件生物量分配之间具有相同的异速生长规律。其中,生殖分配与生产分配、生产分配与茎生物量分配之间均呈幂函数负相关关系,前者的异速指数均α>1,后者的异速指数均约α=1。
[1]李博.生态学[M].北京:高等教育出版社,2000:64-87.
[2]周婵,杨允菲.松嫩平原两个生态型羊草叶构件异速生长规律[J].草业学报,2006,15(5):76-81.
[3]李红,杨允菲,卢欣石.松嫩平原羊草种群生殖分蘖株的数量特征及定量分析[J].草业学报,2004,13(4):50-56.
[4]王俊炜,李海燕,杨允菲.温带地区4种园林灌木叶片的生长规律[J].东北师范大学学报(自然科学版),2005,37(1):95-98.
[5]杨允菲,张宝田.松嫩平原贝加尔针茅无性系构件的结构及生长规律[J].应用生态学报,2005,16(12):2311-2315.
[6]宋金枝,杨允菲.松嫩平原碱化草甸朝鲜碱茅无性系冬眠构件的结构及生长分析[J].生态学杂志,2006,25(7):743-746.
[7]杨允菲,张宝田,张春华.松嫩平原赖草无性系构件的形成与空间扩展实验[J].应用生态学报,2007,18(5):977-982.
[8]郑红梅,杨允菲.松嫩平原天然肥披碱草种群生殖器官的数量性状分析[J].草原与草坪,2007,(4):21-24.
[9]刘玉,王国祥,潘国权.地下水位深度对芦苇生长和叶绿素荧光参数的影响[J].南京师范大学学报(自然科学版),2008,31(3):91-95.
[10]杨允菲,郎惠卿.不同生态条件下芦苇无性系种群调节分析[J].草业学报,1998,7(2):1-9.
[11]杨允菲,李建东.松嫩平原不同生境芦苇种群分株的生物量分配与生长分析[J].应用生态学报,2003,14(1):30-34.
[12]林文芳,陈林姣,朱学艺.用分子标记技术分析不同生态型芦苇的遗传多样性[J].植物生理与分子生物学报,2007,33(1):77-84.
[13]李卫红,周洪华,杨晓明,等.干旱荒漠区草地植物群落地上生物量时空分布对地下水的响应[J].草业学报,2010,19(5):186-195.
[14]李博,刘存岐,闫蓬勃,等.白洋淀湿地芦苇生长参数的线性分析[J].湖北农业科学,2010,49(2):316-318.
[15]李素清,武冬梅,王涛,等.山西长治湿地草本植物优势种群和群落的空间格局分析[J].草业学报,2011,20(3):43-50.
[16]杨允菲,张宝田,田尚衣.松嫩平原旱地生境芦苇种群不同龄级根茎的干物质储藏及水溶糖含量[J].应用生态学报,2008,19(9):1905-1910.
[17]朱学艺,王锁民,张承烈.河西走廊不同生态型芦苇对干旱和盐渍胁迫的响应调节[J].植物生理学通讯,2003,39(4):371-376.
[18]宁祖林,陈慧娟,王珠娜,等.几种高大禾草热值和灰分动态变化研究[J].草业学报,2010,19(2):241-247.
[19]郑慧莹,李建东.松嫩平原盐碱植物群落形成过程的探讨[J].植物生态学报,1995,19(1):1-12.
[20]杨允菲,祝玲,李建东.东北草原豆科、莎草科习见植物籽实千粒重的多样性分析[J].东北师范大学学报,1996,(3):97-102.
[21]郑慧莹,李建东.松嫩平原盐生植物与盐碱化草地的恢复[M].北京:科学出版社,1995:12.
[22]黄迎新,赵学勇,张洪轩,等.沙米表型可塑性对土壤养分、水分和种群密度变化的响应[J].应用生态学报,2008,19(12):2593-2598.
[23]程栋梁.异速生长关系在生物学中的应用[J].沈阳大学学报,2009,21(6):12-15.
[24]张春华,杨允菲.松嫩平原光稃茅香种群生殖分蘖株数量特征分析[J].草业学报,2001,10(3):1-7.
[25]郭力华,杨允菲,李建东.松嫩平原不同生境寸草苔种群生殖分株的数量特征及生殖分配动态[J].草业学报,2005,14(2):63-68.
[26]韩忠明,韩梅,吴劲松,等.不同生境下刺五加种群构件生物量结构与生长规律[J].应用生态学报,2006,17(7):1164-1168.
