耐力、力量和混合运动对成年C57BL/6小鼠骨骼肌卫星细胞激活信号通路的影响
2011-12-29陈彩珍
陈 涛,陈彩珍,卢 健
耐力、力量和混合运动对成年C57BL/6小鼠骨骼肌卫星细胞激活信号通路的影响
陈 涛,陈彩珍,卢 健
目的:研究耐力、力量、混合运动3种训练方式激活静息状态卫星细胞的能力。方法:将32只3月龄C57BL/6小鼠随机分成4组:安静组(C,n=8),耐力运动训练组(E,n=8),力量运动训练组(S,n=8),混合运动训练组(M,n=8)。经过28周运动干涉后,采用实时荧光定量PCR法检测胫骨前肌卫星细胞激活信号通路中相关基因(nNOS,MMP-2,HGF,c-met)mRNA的转录水平。结果:1)与安静组相比,3组运动训练组C57BL/6小鼠胫骨前肌与体重的比值增加;2)耐力训练组nNOS,HGF,c-met mRNA转录水平显著增加,MMP-2 mRNA无显著性差异;3)力量训练组nNOS,MMP-2,HGF,c-met mRNA转录水平显著增加;4)混合训练组nNOS,MMP-2,c-met mRNA转录水平无显著性变化,HGF mRNA转录水平显著增加。结论:力量运动对骨骼肌中卫星细胞的激活具有明显的促进作用,在维持骨骼肌卫星细胞池的数量和促进骨骼肌肥大中具有显著意义;耐力运动能促进卫星细胞的激活,在防止骨骼肌中卫星细胞数量减少,在维持骨骼肌质量和力量上,也有其可取之处;混合训练组在促进卫星细胞激活的过程中,混合运动的效果并不是预期所设想的耐力和力量运动的效果叠加,提示在维持卫星细胞数量和促进骨骼肌肥大上应考虑运动强度、时间和频率,合理安排耐力和力量运动。
卫星细胞;耐力;力量;混合运动训练;神经元型一氧化氮合酶(nNOS);基质金属酶蛋白2(MMP-2);肝细胞生长因子(HGF);鼠;动物实验
随着年龄的增加,骨骼肌质量(II肌纤维为主)减少,力量下降,即肌肉衰减征(sarcopenia)已为多数研究证实,这种肌肉衰减征严重影响了骨骼肌的机能和表现。在肌细胞水平上,肌肉衰减征主要表现为肌纤维数量的减少,尤其以II型肌纤维为主[6]。作为有丝分裂后的细胞,肌纤维的再生依赖于未分化的肌源性祖细胞-骨骼肌卫星细胞[22]。出生后,卫星细胞增殖、分化,为发育的骨骼肌提供所需的肌细胞核。成熟的骨骼肌中,卫星细胞处于静息状态。当肌纤维受到损伤时,卫星细胞被激活,随后增殖、分化,融合成新的肌纤维更换损伤的肌纤维或肌细胞核[13,22]。骨骼肌正常的活动中,细微的肌纤维损伤是习以为常的,特别是随着年龄的增长[8],因此,贯穿整个生命活动,需要不断供给骨骼肌卫星细胞以修复和维持骨骼肌纤维的正常。目前的研究结果表明,老年个体骨骼肌Ⅱ型肌纤维萎缩伴随着Ⅱ型肌纤维内卫星细胞数量的减少[32]。因此,随着年龄的增长,卫星细胞数量减少可能在肌肉衰减征的发生过程中起到重要作用。目前的研究证实了HGF和NO在卫星细胞激活信号通路上的关键作用[29]。这些研究主要通过细胞培养技术,发现机械牵拉卫星细胞或骨骼肌,HGF快速从其储存部位细胞外基质释放并与卫星细胞表面受体c-met结合,从而激活卫星细胞[2,30]。此后的研究发现,HGF的释放依赖卫星细胞或肌纤维上的nNOS生成的NO,NO通过上调MMPs活性介导了HGF的释放[38]。最近的研究证实,运动训练诱导的骨骼肌质量和肌细胞核的增加伴随着卫星细胞数量和激活状态的增加[19,25]。因此,本研究拟对耐力运动、力量运动和耐力、力量混合型运动干预后骨骼肌中卫星细胞激活信号通路中的关键基因进行检测,以期了解运动诱导卫星细胞激活过程中的关键因素,并进一步探讨不同训练方式激活静息状态卫星细胞的能力,从而有利于我们设计出更好的骨骼肌发育和修复方案,以应用到运动科学(提高运动员机能表现)和健康科学(特别是肌肉萎缩和肌肉衰减征的防治方法)上。
1 材料和方案
1.1 实验动物及动物分组
购买3月龄雄性C57BL/6小鼠32只。分笼饲养,每笼1只,每日自由饮水进食,喂食标准饲料,自然光照,饲养环境温度21℃±2℃。所有C57BL/6小鼠经过1周适应性饲养并进行3天适应性训练,随后进行随机分组。安静组、耐力训练组、力量训练组和混合训练组各8只。
1.2 运动方案
耐力训练组使用由跑台改装成的动物跑台进行无负重跑台训练,1~10周速度为0.8~0.9km/h,11~20周速度为0.9~1.0km/h,21~28周为1.0km/h(表1);以小鼠尾部负重爬梯方式建立力量运动模型,起始负重量为10%BW,随后逐渐增加并摸索适宜的负重重量、爬梯重复次数及每组训练的时间间隔(表2),爬梯高1m,台阶间隔为2cm,与地面成85°倾斜;混合训练组为耐力与力量混合的训练模式(表3)。
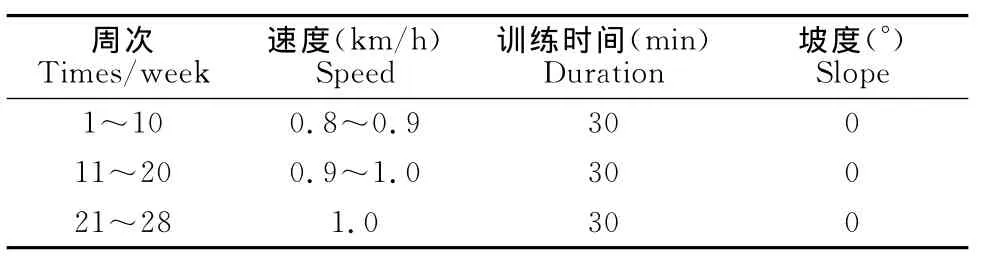
表1 本研究实验动物耐力组训练方案一览表Table 1 Proposal of Endurance Training
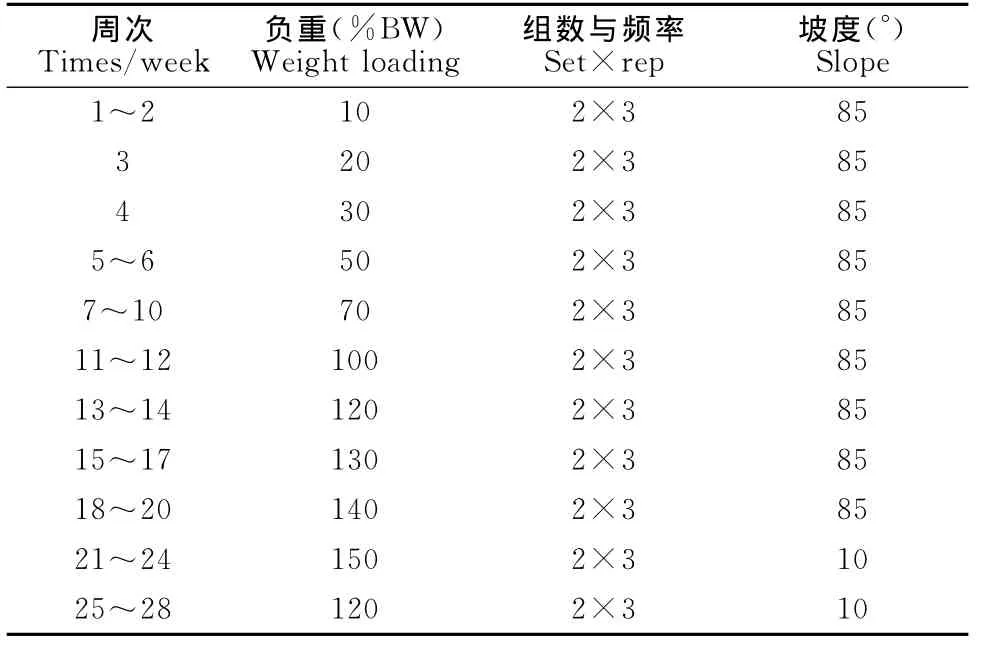
表2 本研究实验动物力量组训练方案一览表Table 2 Proposal of Strength Training

表3 本研究实验动物混合组训练方案一览表Table 3 Proposal of Mixing Type Training
1.3 取材
末次训练后禁食,常规饮水,24h后,给C57BL/6小鼠称重后断颈处死。迅速取完整右侧胫骨前肌,用滤纸吸取血液后,置于称量纸上,称量肌肉质量。PBS缓冲液冲洗后,放入标记好的1.5ml离心管中,放入液氮冻存0.5h后,转移至-80℃超低温冰箱,待检测。
1.4 实时荧光定量PCR检测nNOS、HGF、c-met及MMP-2的基因转录水平
1.4.1 总RNA提取和鉴定
取完整胫骨前肌,按Trizol(Invitrogen)说明书抽提RNA。使用紫外分光光度计检测总RNA的OD260/OD280比值,DEPC水做空白调零。取RNA样品于石英比色皿中,加入DEPC水进行稀释,混匀后放于比色室,记录样品在260nm和280nm波长处的吸收值,并计算OD260/OD280比值。
1.4.2 RNA逆转录反应
取RNA5μl,按照逆转录试剂盒(TaKaRa)配制RT反应液(总体积为10μl),逆转录后得cDNA模板。
1.4.3 实时荧光定量PCR(Real-time PCR)
Realtime PCR反应体系为20μl,其中,SYBR green PCR Master Mix(TOYOBO)10μl,前向、反向引物各1μl,dH2O 6μl,cDNA模板2μl。反应条件,Step 1:预变性(95℃,60s);Step 2:(95℃,15s;55℃/56℃,30s;72℃,35s)× 40个循环;Step 3:建立PCR产物的熔解曲线,再变性(95℃,30s),退火(55℃/56℃,35s),然后,从55℃/56℃缓慢加热到(95℃,15s);每1℃收集荧光一次。经仪器自动分析,各基因融解曲线(Melt Curve)均为单峰,表明扩增产物特异性较高。反应结束后,PCR仪给出各反应孔的Ct值,以GAPDH基因为内参,根据公式2-ΔCt(△CT=CT-target-CTGAPDH)计算各样品目的基因的相对表达量。

表4 本研究各基因引物序列一览表Table 4 Sequences and Details of Target Genes
1.5 数据处理
数据的表示方法为。实验数据由SPSS 18.0统计软件和Excel 2003处理,使用单因素方差分析不同运动训练组与安静组差异显著性,使用独立样本t检验分析不同运动组间差异显著性,P<0.05为显著性差异标准,P<0.01为极显著差异标准。
2 实验结果
2.1 体重以及骨骼肌湿重变化
2.1.1 C57BL/6小鼠体重
3月龄小鼠初始体重为24.4±0.827g,各组无显著性差异。各组小鼠体重随年龄均有所增加,与实验前相比有极显著差异(P<0.01,图1)。处死前,C组小鼠体重显著高于E、S和M组,且具极显著性差异(P<0.01,表5)。

图1 本研究各组小鼠体重随时间变化趋势示意图Figure 1. Weight of Mice in Each Group
2.1.2 骨骼肌湿重与体重的比值
M组胫骨前肌质量与体重比值非常显著高于C组(P<0.01),E组和S组胫骨前肌质量与体重比值显著高于C组(P<0.05,表6)。

表5 本研究各组小鼠处死前体重统计一览表Table 5 Weight of Mice in Each Group
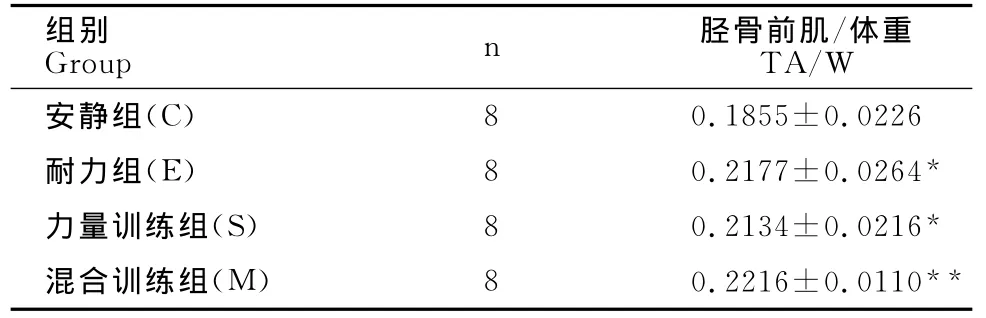
表6 本研究各组小鼠胫骨前肌(TA)与体重比值比较一览表Table 6 Ratio of Tibialis Anterior to Weight in Each Group
注:*表示与C组比较具有显著性差异(P<0.05);**表示与C组比较具有极显著性差异(P<0.01)。
2.2 各组骨骼肌中卫星细胞激活信号通路中各基因荧光定量PCR检测结果
表7显示,E组、S组与C组相比,nNOS mRNA表达增加,具有极显著性差异(P<0.01);M组与C组相比,nNOS mRNA表达无显著差异(P>0.05);与C组相比,S组MMP-2mRNA表达增加,有极显著性差异(P<0.01);E组、M组与C组相比,MMP-2mRNA表达无显著性差异(P>0.05);S组与M组相比,MMP-2mRNA表达增加,且具有显著性差异(P<0.05);E组与C组相比,胫骨前肌HGF mRNA相对表达量有所增加,具有极显著性差异(P<0.01);S组与C组相比,HGF mRNA相对表达量亦增加,且具极显著性差异(P<0.01);M组与C组相比,HGF mRNA相对表达量也增加,并具有显著性差异(P< 0.05);S组与M组相比,HGF mRNA相对表达量增加,且具有显著性差异(P<0.05);E组c-met mRNA表达显著高于C组(P<0.05),S组c-met mRNA非常显著高于C组(P<0.01);M组与C组相比,c-met mRNA表达无显著性差异(P>0.05);S组c-met mRNA显著高于M组(P<0.05)。

表7 本研究各组小鼠胫骨前肌中nNOS、MMP-2、HGF、c-met mRNA转录水平一览表Table 7 nNOS,MMP-2,HGF,c-met mRNA Transcription in Each Group
3 分析与讨论
3.1 不同训练方式对C57BL/6小鼠体重和胫骨前肌湿重的影响
Somerville等研究结果显示,C57BL/6小鼠在其6月龄时,体重和胫骨达到最大值[28]。在本实验的研究中,3月龄C57BL/6小鼠体重随年龄的增长而增加(图1),安静组在实验进行的第25周时体重达到其最大值;3个训练组体重变化趋于一致,21周后趋于较稳定,体重增加幅度均比安静组小。处死前,3组运动训练干预的C57BL/6小鼠体重接近,无显著差异,安静组平均体重最大,与3组运动干预C57BL/6小鼠有极显著差异。胫骨前肌湿重与处死前体重的比值数据显示:混合训练组比值最大,耐力训练组比值次之,其后是力量训练组,最后为安静组,且运动干预的3组数值均比安静组大,且具显著性差异。本实验的结果可以证实,运动训练可以明显地控制体重的增加,这可能因运动训练增强机体脂质代谢有关。
目前的研究都表明,力量运动是一种行之有效的促进骨骼肌肥大、增强骨骼肌质量的方法,而耐力运动则通过减少体脂对去脂体重产生较大的影响。本实验的研究结果表明,运动干预组小鼠骨骼肌质量与体重比明显高于安静组,均具有显著性差异。这种结果可能是由于运动干预组胫骨前肌适应性肥大或去脂体重增加,安静组由于缺乏运动体内脂肪积累过多,两个原因导致该结果的产生。
3.2 不同训练方式对C57BL/6小鼠胫骨前肌nNOS mRNA转录的影响
NO是一个非常小的分子,可以通过细胞膜自由扩散,在分子水平上发挥其生理作用,如调控血压、免疫调节、抑制血小板凝聚、传递神经信息等。骨骼肌、心肌、上皮细胞、内皮细胞、巨噬细胞、成纤维细胞核肝细胞均具NOS,可以生产NO[9]。目前已确定,3种NOS同工酶,骨骼肌中主要表达nNOS并与α1-syntrophin(抗肌萎缩蛋白-糖蛋白复合体),定位在肌膜表面[15]。NO通过HGF激活静息状态下的卫星细胞的信号作用已在许多研究中得到证实。
正常情况下,骨骼肌中的NO一般处于非常低的水平,卫星细胞也处于安静状态,在运动训练、负重、损伤、剪切应力或机械牵拉的刺激下,NOS的表达或活性增高[31]。Balon等让大鼠每天跑台90min,每10min进行1min的冲刺跑,8周后,发现大鼠趾长伸肌和比目鱼肌nNOS蛋白表达增多。Wook Song等人的研究发现,Fischer-344大鼠腓肠肌中,nNOS蛋白表达随着年龄的增加呈下降的趋势,但经过12周的跑台运动,Fischer-344大鼠腓肠肌和比目鱼肌中nNOS蛋白表达水平具有显著性提高[35]。McConell[23]等研究发现,优秀的耐力运动员(˙VO2peak>58ml· kg/min)股外侧肌中,nNOS蛋白表达水平与普通个体相比较多,且具有显著性差异,但mRNA水平并没有太大差异,推测可能因研究对象数目较少;普通个体骑功率自行车进行进行为期10天耐力运动,每天45~90min(75% ˙VO2)其间交替进行间歇训练(90%VO~100%˙VO2),最后一次训练30min后取样,运动训练前股外侧肌中,I、Ⅱa、Ⅱx肌纤维中nNOS蛋白表达水平没什么差异,运动训练后在3种肌纤维中nNOS蛋白表达水平显著升高。这些研究结果证实,耐力运动训练持续时间够达到一定程度或运动强度较大,骨骼肌中nNOS蛋白表达水平就会增多。力量运动训练对nNOS蛋白表达水平的研究或文献较少。在本实验的研究中,经过29周耐力训练,C57BL/6小鼠胫骨前肌中nNOS mRNA表达水平较安静组有所增加,且具有极显著性差异(P<0.01),与其他学者研究耐力运动对骨骼肌中nNOS基因表达影响的结果相符;力量训练组与安静组相比也有所增加,并具有极显著性差异(P<0.01);混合训练组与安静组相比虽有增加的趋势,但没有显著性差异,说明力量和耐力性训练可以显著提高nNOS mRNA表达水平。nNOS mRNA水平的增多可能会导致nNOS蛋白表达的增多从而提高骨骼肌中nNOS酶活性,增加NO生成量。
Leiter[16]等研究发现,雌性C57BL/6小鼠随着年龄的增长(6周龄,6、8、10、18月龄),骨骼肌中卫星细胞对机械牵拉或NO刺激的反应应答逐渐减少,机械牵拉不同年龄阶段小鼠趾长伸肌发现,6周龄和6月龄小鼠趾长伸肌中卫星细胞激活的数量最多,相同强度机械牵拉刺激对8月龄小鼠趾长伸肌中的卫星细胞激活数量影响也没达到显著性水平,但机械牵拉同时,添加NOS底物或NO供体刺激8月龄小鼠趾长伸肌卫星细胞,激活数量显著增加,而18月龄小鼠趾长伸肌中的卫星细胞对这种牵拉刺激或添加NO供体或同时使用两种刺激显得无动于衷。已经在人体和啮齿类动物实验的研究中证实,随着年龄的增加,骨骼肌质量(Ⅱ肌纤维为主)减少,力量下降,即肌肉衰竭症(sarcopenia)[27],骨骼肌中,卫星细胞随着年龄的增长更加趋于有丝分裂安静期。作为卫星细胞激活信号通路的起始信号,NO在骨骼肌中的水平升高可能会促进卫星细胞的激活,本实验研究耐力或力量运动训练均可增加nNOS mRNA水平,因为没有做蛋白水平的测试,只能猜测nNOS蛋白表达或酶活性可能因此而增加,提高骨骼肌中NO含量,从而促进卫星细胞的激活。
3.3 不同训练方式对C57BL/6小鼠胫骨前肌MMP-2 mRNA转录的影响
骨骼肌机能和形态学上的变化是对一些病理或生理刺激的应答。大部分但不是全部,这些应答反应都需要细胞外基质(ECM)的重新塑建(降解和再生)。事实上,骨骼肌对运动适应产生的诸多基因的上调都与细胞外基质有关联。基质金属酶蛋白(MMPs)是一个锌依赖的肽链内切酶大家族,目前,了解较清晰的已有25个家族成员,均有一个前肽区和一个包含锌离子的催化区域。作为肽链内切酶,其家族成员可以降解数种细胞外基质蛋白,如胶原蛋白、弹力蛋白、纤维结合蛋白、蛋白聚糖等[12]。不同的MMPs在骨骼肌中均有表达,在骨骼肌再生、发育及骨骼肌的血管发生的过程中发挥重要作用[4],并证实大鼠骨骼肌中表达MMP-2,机械牵拉可使其转化成52kDa的激活结构[4]。
运动训练导致的局部缺血或肌肉牵拉,可以影响MMP-2和MMP-9的表达水平[4]。NO可以直接通过其亚硝基作用MMP,调节其活性。此外,有相关研究发现,NO可以上调MMP-2和MMP-9的蛋白表达水平和酶活性。Rullman[26]等通过人类实验,观察10天和5周的恒定负荷伸膝练习(45min/天,60次/min),10天和5周的运动训练与训练前相比,股外侧肌中MMP2mRNA表达水平明显升高,MMP-2蛋白活性在训练后10天达到最高水平,5周后MMP-2蛋白活性略有下降。免疫组织化学染色发现MMP-2蛋白主要表达在骨骼肌纤维周围,肌纤维内也有表达,用激光分割肌纤维也观察到MMP-2的mRNA表达,进一步证实了MMP-2在骨骼肌中的表达。Koskinen等人研究发现,大鼠急性下坡跑恢复过程中,MMP-2mRNA水平明显升高[20]。Carmeli[10]等人则发现,不同的运动强度对SD大鼠骨骼肌中MMP-2表达水平影响不同。两周的高强度跑台运动训练(32m/min)后腓肠肌中MMP-2 mRNA和蛋白表达水平显著升高,而低强度的跑台运动训练(18m/min)与安静组相比,MMP-2mRNA和蛋白表达水平并没有发生显著性变化。这些研究均证实,在非病理的情况下,运动训练可以激活骨骼肌中的MMPs。这也就是说,急性运动或持续性运动诱导的肌肉损伤或重建过程中,通过调节MMP-2的表达水平完成修复或重塑。在本实验的研究中发现,耐力和混合组胫骨前肌MMP-2mRNA相对表达量有所增加,与安静组相比,无显著性差异,力量训练组和安静组相比,MMP-2mRNA相对表达量增加,且具有极显著性差异,可能是因为力量运动训练产生的牵拉刺激强烈,力量运动训练通过重复提升负重,对骨骼肌小肌群进行反复刺激,与耐力性运动训练相比对骨骼肌肥大影响显著从而增加骨骼肌力量,对骨骼肌的重塑和再建作用较大,从而促进MMP-2的表达增多。本实验中,耐力运动训练属于低强度的有氧训练,运动过程中,小鼠骨骼肌损伤的可能性较小,因此,骨骼肌中MMP-2表达变化不多,这也与Carmeli等的研究结果相符。
MMPs在运动诱导的骨骼肌重塑发挥许多作用,除了已经证实的血管发生作用,相关体外实验研究表明细胞外基质中MMPs裂解分子和连接蛋白的作用大大提高了诸多生长因子的生物利用率[12]。Rullman等发现,只需几分钟的运动就可增加血浆和细胞外液中速率限制的生血管因子VEGF(血管内皮生长因子)[26]。这些都说明,相关生长因子在生理情况下储存在细胞外基质中,其释放也不需事先转录和翻译。因此可以推测,运动诱导的蛋白水解作用介导了储存在细胞外基质生长因子的释放。Yamada等人对离体培养的大鼠骨骼肌卫星细胞的研究发现,NO通过MMPs把HGF从细胞外基质中释放出来,NO可能通过激活MMPs的酶内切活性,裂解与HGF结合的蛋白聚糖的核心蛋白或氨基葡聚糖链,从而将HGF从细胞外基质中释放出来[38]。虽然目前仍不是很清楚是何种MMPs负责HGF的释放,前期的研究证实骨骼肌中表达MMP-2、MMP-3、MMP-7、MMP-9,其中,MMP-2、MMP-3、MMP-9可以裂解硫酸乙酰肝素蛋白聚糖的核心蛋白[11]。因此,在机械牵拉激活卫星细胞的级联反应中,MMP-2可能参与HGF的释放,并依赖NO。此外有研究发现,HGF一般和蛋白聚糖的胞外区域形成的复合体从细胞外基质释放,一起结合到c-met受体上[1]。事实上,HGF与部分硫酸乙酰肝素形成的复合体和单独的HGF相比对c-met有更好的亲和力[24]。
综上所述,MMPs家族在骨骼肌正常生理状态下即有表达,在本实验的研究中,力量运动训练组与安静组相比,MMP-2mRNA表达水平显著升高,蛋白水平可能也随之升高,力量运动对骨骼肌重塑和在建的作用较大,并可以因此通过上调MMP-2的基因表达,增加HGF生物利用率,促进卫星细胞的激活。
3.4 不同训练方式对C57BL/6小鼠胫骨前肌HGF mRNA转录的影响
早期的实验研究已经证实,IGF-1和FGFs等生长因子虽然能够促进卫星细胞的增殖,但前提是卫星细胞已经处于激活状态[24]。TGF-β(转化生长因子β)和EGF(内皮细胞生长因子)等也不能促进安静状态下的卫星细胞进入细胞周期[17]。截止到目前为止,在对所有的生长因子的研究中,HGF是惟一一个经过证实能激活安静状态下卫星细胞的生长因子。未损伤的骨骼肌中,HGF与细胞外基质主要组成部分蛋白聚糖连接在一起,储存在肌细胞外基质中,当骨骼肌损伤时,HGF能快速从细胞外基质释放出来并与卫星细胞膜上c-met结合,激活卫星细胞[3]。目前,主要是通过细胞培养技术,研究HGF对骨骼肌卫星细胞激活作用,而有关运动训练对HGF在活体骨骼肌中卫星细胞激活作用的相关文献或报道几乎没有。在本实验的研究中发现,3种不同的运动训练方式使C57BL/6小鼠胫骨前肌HGF mRNA转录水平均比安静组高,且耐力训练组和力量训练组与安静组相比具有极显著性差异,混合组和安静组相比具有显著性差异。HGF mRNA水平的上升,可能会导致HGF蛋白表达的增多,使得细胞外基质中储存的HGF水平升高,在NO信号刺激的作用下,HGF释放增多,从而促进激活的卫星数量增加。
虽然众多的研究证实,HGF作为惟一一个能激活卫星细胞的生长因子,但体外培养实验发现,过高的HGF水平减少了机械牵拉诱导的卫星细胞激活。Wozniak[36]等发现,机械牵拉肌原细胞,同时,在其培养基中添加高浓度的HGF,机械牵拉虽然能够诱导内源性HGF从肌纤维外基质中释放并上调卫星细胞表面c-met表达水平,激活的卫星细胞数量却减少了。Wozniak等推测可能原因是:1)由于高水平的HGF,导致受体c-met脱敏作用;2)细胞外基质pH或HGF与c-met结合位点发生构象改变。Yamada[39]等使用细胞培养技术发现,骨骼肌再生和运动诱导的骨骼肌肥大依赖卫星细胞的激活和安静状态,机械牵拉诱导HGF释放引发卫星细胞激活,随后myostatin的表达增多可能作为一种信号介导了激活卫星细胞返回其安静状态。Yamada[39]研究小组证实,卫星细胞激活增殖的过程中,HGF在激活的卫星细胞返回其安静状态可能通过其浓度依赖性负反馈调节机制,即高水平表达HGF可能上调myostatin mRNA水平,增加myostatin蛋白的分泌和表达水平,从而使激活的卫星细胞恢复其安静状态。该研究提示,HGF可以通过其浓度依赖性负反馈调节机制,对卫星细胞的状态进行调节。这些研究结果说明,骨骼肌纤维损伤或运动刺激下可以通过释放细胞外基质中的HGF激活卫星细胞,修复损伤肌纤维或产生适应性变化,也可通过调节HGF的表达或释放水平,使得尚未分化融合的卫星细胞恢复其安静状态,以维持卫星细胞池自身的数量。在本实验的研究中,3种运动训练方式均导致HGF mRNA表达水平的增多,作为对运动训练产生的适应性变化,骨骼肌纤维外基质储存的HGF蛋白可能也会同步增多,从而提高激活骨骼肌中卫星细胞的潜能。
3.5 不同训练方式对C57BL/6小鼠胫骨前肌c-met mRNA转录的影响
c-met作为HGF受体,是一个广泛表达的异源二聚体受体络氨酸激酶,在骨骼肌中以跨膜蛋白的形式结合在卫星细胞表面[7]。HGF结合到c-met上使其形成αβ异源二聚体,络氨酸自磷酸化形成募集多种信号转导中介[5]。一些研究发现,HGF可通过c-met诱导ERK磷酸化,激活MAPK家族中的p38β/β,同时,伴随着卫星细胞的激活,抑制p38β/β可使卫星细胞保持其安静状态,使其与外界信号刺激隔绝[18,34]。c-met作为HGF受体,同时,也是卫星细胞的一种标记物,很多研究证实在所有卫星细胞均表达c-met且不分肌纤维类型,活体实验也在安静的和再生的骨骼肌中均检测到c-met蛋白,此外,急性损伤的骨骼肌纤维被大量的单核细胞所包围且表达c-met,即意味着c-met同样可以标记MPCs(生肌性祖细胞)[14]。这些研究表明,c-met可以作为卫星细胞的一种独特标记物,方便了我们用免疫组织化学的方法来鉴别卫星细胞。
Wozniak[36]等通过离体培养单根肌纤维模型,研究了c-met在卫星细胞激活过程中的作用和变化,机械牵拉正常的肌纤维,c-met的表达增多,在培养基中添加放线菌酮(抑制蛋白质合成)也不会抑制c-met的表达,证实c-met和c-fos一样属于即刻早期应答基因。目前尚未发现有关运动训练对骨骼肌中c-met影响的相关文献或报道。在本实验研究中发现,耐力和力量训练均使C57BL/6小鼠胫骨前肌c-met mRNA表达水增多,且具显著和极显著差异,混合训练组虽然与安静组相比c-met mRNA表达有所增多,但没有显著差异。c-met突变小鼠不能形成正常的骨骼肌群[12],说明在NO依赖HGF-c-met激活卫星细胞的信号通路上,c-met重要作用。c-met mRNA表达水增多可能是单个卫星细胞表面蛋白的表达增多,也可能是卫星细胞数量的增多。运动训练对骨骼肌中卫星细胞作用的相关文献大都是对力量运动训练的研究,而有关耐力运动训练的很少。Verney等对人体试验的研究中,让受试者进行下肢耐力运动训练和上肢力量训练相结合的一种训练方式,14周以后发现三角肌和股外侧肌中卫星细胞的数量与训练前相比增加了38%左右,而且这种增加主要发生在Ⅱ型肌纤维中[33]。耐力运动训练诱导的骨骼肌适应性反应主要是通过改善心肺功能,增加骨骼肌毛细血管密度和毛细血管管腔的肥大,可能通过血液循环为骨骼肌提供丰富的相关生长因子,增加卫星细胞的激活数量。本实验的研究结果表明,nNOS、HGF、c-met mRNA表达水平在运动训练尤其是力量运动训练的刺激下均有所增加,可能促使NO依赖HGF-c-met激活卫星细胞的信号通路的作用增强,从而使得卫星细胞激活、增殖增加,卫星细胞的数量增加。
4 结论
1.C57BL/6小鼠体重增长趋于稳定时,3种运动训练组体重均显著低于安静组,说明运动训练能较好地控制体重的增加。
2.与安静组相比,3组运动训练组C57BL/6小鼠胫骨前肌与体重的比值增加。耐力运动组的增加可能因体内脂肪的减少;力量运动组的增加可能因胫骨前肌适应性肥大;混合训练组可能因胫骨前肌适应性肥大及体脂的减少。
3.力量训练组上调C57BL/6小鼠胫骨前肌NOMMP-2-HGF-c-met卫星细胞激活通路各基因转录水平。推测力量运动对骨骼肌中卫星细胞的激活具有明显的促进作用,在维持骨骼肌卫星细胞池的数量和促进骨骼肌肥大中具有显著意义。
4.耐力训练组上调C57BL/6小鼠胫骨前肌NOMMP-2-HGF-c-met卫星细胞激活通路中nNOS、HGF、cmet基因转录水平,推测耐力运动也能促进卫星细胞的激活,在防止骨骼肌中卫星细胞数量的减少,维持骨骼肌质量和力量上,也有其可取之处。
5.混合训练组对C57BL/6小鼠胫骨前肌NO-HGF-cmet卫星细胞激活通路基因转录水平影响较小,仅HGF表达水转录的上调达到显著水平。在本研究中,混合运动的效果并不是预期所设想的耐力和力量运动效果的叠加,提示在维持卫星细胞数量和促进骨骼肌肥大上应考虑运动强度、时间和频率,合理安排耐力和力量运动。
[1]ALLEN R E,GOLL D E.Cellular and developmental biology of skeletal muscle as related to muscle growth[A].SCANES CG.Biology of Growth of Domestic Animals[M].Lowa State Press,2003:148-169.
[2]ANDERSON J,PILIPOWICZ O.Activation of muscle satellite cells in single-fiber cultures[J].Nitric Oxide,2002,7(1):36-41.
[3]ANDERSON J E.A role for nitric oxide in muscle repair:nitric oxide-mediated activation of muscle satellite cells[J].Mol Biol Cell,2000,(11):1859-1874.
[4]BAUMO,GANSTER M,BAURNGARTNER I,et al.Basement membrane remodeling in skeletal muscles of patients with limb ischemia involves regulation of matrix metalloproteinases and tissue inhibitor of matrix metalloproteinases[J].J Vasc Res,2007,(44):202-213.
[5]BIRCHMEIER C,GHERARDI E.Developmental roles of HGF/SF and its receptor,the c-Met tyrosine kinase[J].Trends Cell Biol,1998,(8):404-410.
[6]BONNEFOY M.Sarcopenia,muscle function and prevention[J].
Nut Clin Metab,2004,(18):175-180.
[7]BOTTARO D P,RUBIN J S,FALETTO D L,et al.Identification of the hepatocyte growth factor receptor as the c-met protooncogene product[J].Sci,1991,(251):802-804.
[8]BROOKS S V,FAULKNER J A.The magnitude of the initial injury induced by stretches of maximally activated muscle fibres of mice and rats increases in old age[J].J Physiol,1996,497(2):573-580.
[9]C BOGDAN.Nitric oxide and the regulation of gene expression[J].Trends Cell Biol,2001,(11):66-75.
[10]CARMELI E,MOAS M,LENNON S,et al.High intensity exercise increases expression of matrix metalloproteinases in fast skeletal muscle fibres[J].Exp Physiol,2005,90(4):613-619.
[11]DERKSEN P W,KEEHNEN R M,EVERS L M,et al.Cell surface proteoglycan syndecan-1mediates hepatocyte growth factor binding and promotes Met signaling in multiple myeloma[J].Blood,2002,(99):1405-1410.
[12]F BLADT,D RIETHMACHER,S ISENMANN,et al.Essential role for the c-met receptor in the migration of myogenic precursor cells into the limb bud[J].Nature,1995,(376):768-771.
[13]HAWKE T J,GARRY D J.Myogenic satellite cells:physiology to molecular biology[J].J Appl Physiol,2001,91(2):534-551.
[14]HOTERMAN C E,RUDNICKI M A,et al.Molecular regulation of satellite cell function[J].Semin Cell Dev Biol,2005,(16):575-584.
[15]J S STAMLER,G MEISSNER.Physiology of nitric oxide in skeletal muscle[J].Physiol Rev,2001,81(1):209-237.
[16]JEFF R S LEITER,JUDY E.Anderson.Satellite cells are increasingly refractory to activation by nitric oxide and stretch in aged mouse-muscle cultures[J].Int J Biochem Cell biol,2009:132-136.
[17]JOHNSON S E,ALLEN R E.Activation of skeletal-muscle satellite cells and the role of fibroblast growth-factor receptors[J].Experimental Cell Res,1995,219:449-453.
[18]JONES N C,TYNER K J,NIBARGER L,et al.The p38alpha/beta MAPK functions as a molecular switch to activate the quiescent satellite cell[J].J Cell Biol,2005,169:105-116.
[19]KADI F.Cellular and molecular mechanisms responsible for the action of testosterone on human skeletal muscle.A basis for illegal performance enhancement[J].Br J Pharmacol,2008;154(3):522-528.
[20]KOSKINEN S O,WANG W,AHTIKOSKI A M,et al.Acute exercise induced changes in rat skeletal muscle mRNAs and proteins regulating type IV collagen content[J].Am J Physiol Regul Integr Comp Physiol,2001,280:R1292-R1300.
[21]LEE S,FARRAR R P.Resistance training induces muscle-specific changes in muscle mass and function in rat[J].J Exe Physicol Online,2003,6(2):80-87.
[22]MAURO A.Satellite cell of skeletal muscle fibers[J].J.Biophys Biochem Cytol,1961,(9):493-495.
[23]MCCONELL G K,BRADLEY S J,STEPHENS T J,et al.Skeletal muscle nNOSμprotein content is increased by exercise training in humans[J].Am J Physiol,2007,(62):821-828.
[24]PAPY-GARCIA D,BARBOSA I,DUCHESNAY A,et al.Glycosaminoglycan mimetics(RGTA)modulate adult skeletal muscle satellite cell proliferation in vitro[J].J Biomedical Materials Res,2002,(62):46-55.
[25]ROTH S M,MARTEL G F,IVEY F M,et al.Skeletal muscle satellite cell characteristics in young and older men and women after heavy resistance strength training[J].J Gerontol A Biol Sci Med Sci,2001,56(6):B240-247.
[26]RULLMAN E,NORRBORN J,HAAS T,et al.Endurance training activates matrix metalloproteinases in human skeletal muscle[J].J Appl Physiol,2009,(106):804-812.
[27]SMYTHE G M,SHAVLAKADZE T,RPBERTS P,et al.Age influences the early events of skeletal muscle regeneration:studies of whole muscle grafts transplanted between young(8 weeks)and old(13-21months)mice[J].Exp Gerontol,2008,(43):550-62.
[28]SOMERVILLE J,ASPDEN R,ARMOUR K,et al.Growth of C57Bl/6Mice and the Material and Mechanical Properties of Cortical Bone from the Tibia.[J]Calcif Tissue Int,2004,(74):469-475.
[29]TATSURNI R,ALLEN R E.Mechano-biology of resident myogenic stem cells:Molecular mechanism of stretch-induced activation of satellite cells[J].Anim Sci J,2008,79(3):279-290.
[30]TATSUMI R,HATTORI A,IKEUCHI Y,et al.Release of hepatocyte growth factor from mechanically stretched skeletal muscle satellite cells and role of pH and nitric oxide[J].Mol Biol Cell,2002,13(8):2909-2918.
[31]TIDBALL J G,LAVERGNE E,LAU K S,et al.Mechanical loading regulates NOS expression and activity in developing and adult skeletal muscle[J].Am J Physiol Cell Physiol,1998,(275):C260-C266.
[32]VERDIJK L B,KOOPMAN R,SCHAART G,et al.Satellite cell content is specifically reduced in type II skeletal muscle fibers in the elderly[J].Am J Physiol Endocrinol Metab,2007,292(1):E151-E157.
[33]VEMEY J,KADI F,CHARIFI N,et al.Effects of combined lower body endurance and upper body resistance training on the satellite cell pool in elderly subjects[J].Muscle Nerve,2008,(38):1147-1154.
[34]VOLONTE D,LIU Y,GALBIATIE F,et al.The modulation of caveolin-1expression controls satellite cell activation during muscle repair[J].Faseb J,2005,(19):237-239.
[35]W SONG,H B KWAK,J H KIM,et al.Exercise training modulates the nitric oxide synthase profile in skeletal muscle from old rats[J].J Gernotol,2009,64A(5):540-549.
[36]WOZNIAK A C,ANDERSON J E.Nitric Oxide-dependence of satellite stem cell activation and quiescence on normal skeletal muscle fibers[J].Dev Dyn,2007,236(1):240-250.
[37]YAMADA M,SANKODA Y,TATSUMI R,et al.Matrix metalloproteinase-2mediates stretch-induced activation of skeletal muscle satellite cells in a nitric oxide-dependent manner[J].Int J Biochem Cell Biol,2008,(40):2183-2191.
[38]YAMADA M,TATSUMI R,SANKODA Y,et al.Matrix metalloproteinase-2mediates stretch-induced activation of skeletal muscle satellite cells in a nitric oxide-dependent manner[J].Int J Biochem Cell Biol,2008,40(10):2183-2191.
[39]YAMADA M,TATSUMI R,YAMANOUCHI K,et al.High concentrations of HGF inhibit skeletal muscle satellite cell proliferation in vitro by inducing expression of myostatin:apossible mechanism for reestablishing satellite cell quiescence in vivo[J].J Physiol,2009,298c:465-467.
The Effects of Endurance,Strength and Mixing Type Training on Satellite Cell Activation Signaling Pathway in Adult C57BL/6Mouse Skeletal Muscle
CHEN Tao,CHEN Cai-zhen,LU Jian
Objective:To investigate the effect of endurance,strength and mixing type training on satellite activation.Methods:32three-month-old male C57BL/6mice were randomly distributed into four groups:sedentary(C,n=8),endurance training(E,n=8),strength training(S,n=8),mixing type training(M,n=8).After 28weeks exercise intervening,quantitative real-time PCR method is used to detect nNOS,MMP-2,HGF,c-met mRNA expression in tibialis anterior muscles.Results:1)Compared with the controls,tibialis anterior and the weight ratio increases in three types of exercise training.2)Endurance training significantly increased nNOS,HGF,c-met mRNA transcription except MMP-2mRNA.3)Strength training significantly increased nNOS,MMP-2,HGF,c-met mRNA transcription.4)Mixing type training significantly increased HGF mRNA transcription,but had no apparent effect on nNOS,MMP-2,HGF,c-met mRNA transcription.Conclusion:Strength training plays an important role in satellite cell activation,which may be important in maintaining the number of satellite cell pool and promoting extensive skeletal muscle hypertrophy.Endurance training can also promote the activation of satellite cells and would also have its merits in the prevention the number of satellite cells reduction and maintenance skeletal muscle mass and strength.The mixed type training cannot play a composite effect of endurance and strength training in the activation process of satellite cell.It suggested that we should consider the intensity,time and frequency when we arrange endurance and strength training to sustain the number of skeletal muscle satellite cells and promote muscle hypertrophy.
satellitecell;endurancetraining;strengthtraining;mixingtypetraining;Sarcopenia;neuronalnitricoxidesynthase(nNOS);matrixmetalloproteinase-2(MMP-2);hepatocytegrowthfactor(HGF)
1000-677X(2011)09-0069-08
2011-03-28;
2011-08-10
上海市浦江人才计划资助项目(10PJC029)。
陈涛(1984-),男,安徽临泉人,在读博士研究生,主要研究方向为运动与健康,E-mail:chentwhy@gmail.com;陈彩珍(1967-),女,福建上杭人,教授,博士,主要研究方向为运动生物化学,E-mail:caizhenchen@163.com;卢健(1962-),男,四川绵阳人,教授,博士,主要研究方向为运动与衰老,E-mail:jlu@tyxx.ecnu.edu.cn。
华东师范大学体育与健康学院,上海200241
East China Normal University,Shanghai 200241,China.
G804.7
A
