西藏墨脱县多斑按蚊复合体尚未发生击倒抗性突变*
2011-11-13汤林华周水森叶苗青洪结凤俞俊峰
刘 茜,武 松,汤林华,周水森,黄 芳,叶苗青,洪结凤,俞俊峰
2.中国疾病控制中心寄生虫病预防控制所,世界卫生组织疟疾、血吸虫病和丝虫病合作中心,上海 200025
西藏自治区的疟疾流行历史较为久远,病例主要分布于雅鲁藏布江河谷下游林芝地区的墨脱和察隅两县,其中以墨脱县为主,该县为我国疟疾发病最高的地区之一。媒介防制为疟疾消除非常重要的手段,使用拟除虫菊酯类杀虫剂进行媒介控制是当前媒介成蚊防制的重要措施。在杀虫剂控制媒介的过程中应该密切监测蚊虫对杀虫剂的敏感性。目前我国已有多次报道传疟按蚊出现拟除虫菊酯类杀虫剂抗性[1-6],敏感性检测主要采用的是WHO推荐的接触法,该方法要求必须将现场捕获的一定数量的饱血按蚊带回实验室产卵,孵育至子一代蚊虫方可进行测试,对技术要求高、难于实施,尤其对交通极为不便,防疟队伍薄弱的墨脱县,该法几乎不可能实施。笔者对该县的传疟媒介伪威氏按蚊的钠离子通道蛋白基因进行了扩增和测序,结果报道如下。
1 材料与方法
1.1 蚊虫来源 按蚊成蚊采用人帐诱和诱蚊灯诱捕自墨脱县达木乡达木村(调查点海拔1 408m,N:29°49.627,E:95°46.227)与珠村(调查点海拔 1 510m,N :29°31.120,E :95°25.464);墨脱镇墨脱村(调查点海拔 1178m,N :29°19.285,E :95°19.862);低海拔背崩乡地东村(调查点海拔 853m,N:29°21.905,E:95°09.829)和背崩村(调查点海拔831m,N :29°24.563,E:95°17.444)。 现场调查时间为2010年7月14日至2010年8月17日,多斑按蚊复合体具体种型通过分子方法鉴定[7]。多斑按蚊复合体5成员种钠离子通道蛋白基因序列研究中伪威氏按蚊与威氏按蚊来自西藏墨脱县和云南勐腊,多斑按蚊、塞沃按蚊和达罗毗按蚊基因组由第二军医大学马雅军教授惠赠。
1.2 主要仪器和试剂 PCR扩增仪(PTC-200型)购自美国Bio-Rad公司;凝胶成像系统(Alpha Imager HP/EP)购自美国Alpha Innotech公司;电泳仪(DYY-2C型)购自北京六一仪器厂;DNA聚合酶和脱氧核苷三磷酸(dNTPs)购自上海赛百盛基因技术有限公司;RNA酶和蛋白酶K购自上海鼎国生物技术有限公司。
1.3 分子实验方法
1.3.1 基因提取 采用简易快速DNA提取法[8],将1~2个蚊腿置于1.5mL离心管,加入35μ L裂解液(10mmol/L pH8.0 Tirs HCl,1mmol/L EDTA,1%Nonidet P-40,100μ g/mL 蛋白酶 K),匀浆 ;再加入35μ L消毒双蒸水,煮沸 5min;10 000r/min离心2min,取上清液置-20℃备用。
1.3.2 钠离子通道蛋白基因扩增 采用文献报道[9]的简并引物进行多斑按蚊复合体5成员种钠离子通道蛋白基因IIS4-S6区段基因扩增,正向引物Gen1F :5′-GGAGARTGGATYGA RTCNATGTGGGA-3′ , 反 向 引 物 Gen1R:5′-TTNGACAAAAGCAAGGCTAAG AA-3′。PCR反应体系为 50 μ L :10 ×PCR 缓 冲液 ,5.0 μ L,25 mmol/L MgCl24 μ L,10 mmol/L dNTPs 1.0 μ L,正向 、反向引物各0.8 μ L(20 mmol/L),red Taq聚合酶 2.0 μ L(1 U/μ L)、DNA 模板 2.5 μ L,加水补至 50 μ L 。PCR反应条件:95℃10 min,94℃30 s,50℃30 s,72℃45 s,共35个循环,最后72℃温育5min。
1.3.3 测序验证 对上述扩增出的PCR产物进行1%琼脂糖凝胶电泳,并成像拍照,切取目标条带进行纯化,采用测序试剂盒和全自动测序仪对扩增产物双向测序(由上海生工生物工程技术服务有限公司完成),据测序图验证PCR抗性检测结果,多斑按蚊复合体5成员种序列比对采用Bioedit软件完成。
2 结 果
我国多斑按蚊复合体5成员种采用简并引物全部扩增出片段约为208bp的目标条带,目标条带测序比对,获得5成员种VGSC IIS4-S6区段基因序列信息如图1。图中加框所在位置为钠离子通道蛋白基因L1014位点,图中可见多斑按蚊复合体5成员种该位点序列一致,均为TTA,即亮氨酸(L)。

图1 我国多斑按蚊复合体钠离子通道蛋白基因DII6区段基因序列信息Fig.1 Voltage-gated sodium channel DII6 segment nucleotide sequence of five members of Anopheles maculatus complex in China
对调查点捕获的多斑按蚊复合体进行分子种型鉴定后,选取墨脱县达木乡达木村(伪威氏按蚊20只,威氏按蚊20只)与珠村(伪威氏按蚊20只,威氏按蚊20只);墨脱镇墨脱村(伪威氏按蚊20只,威氏按蚊2只);低海拔背崩乡地东村(伪威氏按蚊20只,威氏按蚊12只)和背崩村(伪威氏按蚊20只,威氏按蚊7只)。共伪威氏按蚊100只,威氏按蚊61只,所有标本测序后,均未发现突变,结果见图2。图中加框位置为L1014位点,所有标本该位置测序结果显示均未发生突变。
3 讨 论
多斑按蚊复合体是马来西亚、越南、泰国、缅甸、印度尼西亚、老挝等地的传疟媒介之一[10-12]。董学书[13]研究发现云南省伪威氏按蚊和多斑按蚊的人血指数分别为42.88%和35.71%,其密度的季节消长与疟疾流行关系密切。周红宁[14]采用ELISA方法检测多斑按蚊复合体中疟原虫环子孢子蛋白(CSP)发现阳性。武松[15]等发现西藏疟疾流行区墨脱县多斑按蚊复合体包括伪威氏按蚊和威氏按蚊,并且在伪威氏按蚊唾液腺中PCR扩增出间日疟子孢子基因,从分子角度判定了伪威氏按蚊为墨脱县的传疟媒介。同时笔者于2010年在西藏墨脱达木乡研究发现威氏按蚊种群密度高于伪威氏按蚊,也初步具备传疟条件,因此笔者对西藏的这两种按蚊的钠离子通道蛋白基因均进行了扩增研究。
本次研究采用简并引物,扩增出了我国多斑按蚊复合体5成员种的VGSC IIS4-S6区段基因,为以多斑按蚊复合体为传疟媒介的地区拟除虫菊酯抗性检测提供了基因信息,5成员种L1014位点均为T TA(亮氨酸)。本次共测序伪威氏按蚊和威氏按蚊161只,经仔细核对测序图,所有标本L1014位点均未发生突变。伪威氏按蚊和威氏按蚊在墨脱县不同海拔分布见差异悬殊,因此在低海拔的墨脱镇和背崩乡仅捕获的威氏按蚊19只,全部进行扩增测序分析。中高海拔达木乡(约1 500m),两个自然村捕获的伪威氏按蚊和威氏按蚊均较多,因此每村两种按蚊各选取20只进行分析。研究结果显示墨脱县的伪威氏按蚊和威氏按蚊击倒抗性均尚未发生,意味着两种按蚊对拟除虫菊酯类杀虫剂应该敏感。主要因为该地区村民几乎不使用防蚊药品,同时按蚊滋生地稻田常年水质流动,致使本来使用量就较少的农药杀虫剂对按蚊的抗性产生影响较小。
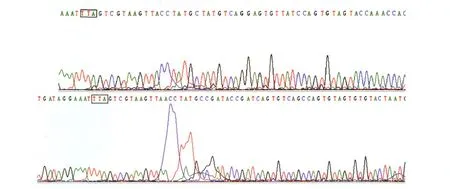
图2 伪威氏按蚊与威氏按蚊L1014位点测序信息图Fig.2 Map of VGSCL1014 of An.pseudowillmori and An.willmori upper part:An.pseudowillmori;below part:An.willmori
本研究初步获得了我国多斑按蚊复合体5成员种的钠离子通道蛋白基因序列信息,并通过测序发现西藏疟疾流行区伪威氏按蚊和威氏按蚊均未发生击倒抗性突变,为该地区在疟疾防治中实施媒介控制方案提供了分子实验的依据,并为该地区实施疟疾消除过程中进行媒介抗性监测提供了前期的分子依据,并为进一步建立分子抗性检测方法提供了基础。
[1]王伟明,金小林,周华云,等.江苏省全国疟疾监测点疫情分析[J].中国血吸虫病防治杂志,2007,19(3):220-223.
[2]沈宝祥,李菊林,周华云.中华按蚊对常用菊酯类杀虫剂抗性现场调查[J].中国血吸虫病防治杂志,2002,14(2):148-149.
[3]潘波,朱泰华,刘勇鹰,等.我国主要传疟媒介对杀虫剂的敏感性现状[J].中国媒介生物学及控制杂志,2001,12(2):145-148.
[4]张华勋,李凯杰,胡乐群,等.中华按蚊对常用杀虫剂的抗药性监测[J].中国热带医学,2010,10(4):400-401.
[5]李菊林,周华云,曹俊,等.江苏省部分地区中华按蚊对溴氰菊酯和氟氯氰菊酯的敏感性调查[J].中国媒介生物学及控制杂志,2009,20(5):438-439.
[6]李志,杨德香,丁俊,等.蚊虫抗药性监测研究[J].中华卫生杀虫药械,2007,13(4):256-258.
[7]武松,潘嘉云,王学忠,等.西藏墨脱县疟疾流行区多斑按蚊复合体种型鉴定[J].中国寄生虫学与寄生虫病杂志.2008,26(4):286-289.
[8]Kambhampati S,Black WC,Rai KS.Random amplified polymorphic DNA of mosquito species and population(Diptera:Culicidae):techniques,statistical analysis,and applications[J].Med Entomology,1992,29(6):939-945.
[9]Enayati AA,Vatandoost H,Ladonni H,et al.Molecular evidence for a kdr-like pyrethroid resistance mechanism in the malaria vector mosquito Anopheles stephensi[J].Medical and Veterinary Entomology,2003,17:138 144.
[10]Rahman WA,Adanan CR,Abu Hassan A.Species composition of adult Anopheles populations and their breeding habitats in Hulu Perak district,Peninsular Malaysia[J].Southeast Asian J T rop Med Public Health,2002,33(3):547-50.
[11]Kengluecha A,Singhasivanon P,Tiensuwan M,et al.Water quality and breeding habitats of anopheline mosquito in no rthwestern T hailand[J].Southeast Asian J T rop M ed Public Health,2005,36(1):46-53.
[12]Vythilingam I,Phetsouvanh R,Keokenchanh K,et al.T he prevalence of Anopheles(Diptera:Culicidae)mosquitoes in Sekong Province,Lao PDR in relation to malaria transmission[J].T rop Med Int Health,2003,8(6):525-35.
[13]董学书.云南多斑按蚊种团的地理分布、生态习性与疟疾的关系[J].寄生虫与医学昆虫学报,1996,3(2):100-105.
[14]周红宁,张再兴,Chris Curtis,等.ELISA检测云南按蚊环子孢子蛋白的评价[J].中国寄生虫学与寄生虫病杂志,2004,22(4):227-230.
[15]Wu S,Pan J Y,Wang XZ,et al.Anopheles pseudowillmori isthe predominant malaria vector in Motuo County,Xizang Autonomous Region[J].Malaria Journal,2009,8:46.
