不同条件下Camembert奶酪中无毒害李斯特菌空间生长分布
2011-11-02刘少伟
黄 微,刘 雪,刘少伟,﹡
(1.华东理工大学,上海200237;2.中国农业大学,北京100083)
不同条件下Camembert奶酪中无毒害李斯特菌空间生长分布
黄 微1,刘 雪2,刘少伟1,﹡
(1.华东理工大学,上海200237;2.中国农业大学,北京100083)
研究了在Camembert奶酪成熟过程中无毒害李斯特菌存活和生长的能力。用含有5lg CFU/mL无毒害李斯特菌的巴氏灭菌全脂牛奶精心制备Camembert奶酪。所有Camembert奶酪在以下三种情况下成熟:室温(约20℃)和相对湿度60%(36h);12℃、相对湿度93%(2周);7℃、相对湿度85%(3周)。在成熟过程中,分别在1、5、10、15、20、25、30、35d时,利用选择性培养基对奶酪表面和内部的无毒害李斯特菌进行计数。结果显示,在成熟第1d时,无毒害李斯特菌数量为7.16lg CFU/g,比初始值的4.76lg CFU/g增加了2.40lg CFU/g;而在第20d时,无毒害李斯特菌数量降低至6.54lg CFU/g;在第35d时,李斯特菌数量增加至7.38lg CFU/g。总体而言,无毒害李斯特菌在奶酪表面的生长快于中心位置。
无毒害李斯特菌,Camembert奶酪,成熟,存活,生长
1 材料与方法
1.1 材料与仪器
巴氏杀菌牛奶、培养基 市场购得;L.innocua、L.monocytogenes菌种保藏中心提供;酵母提取物 喝彩食品有限公司;大豆酪蛋白胨琼脂培养基 上海振宇化工科技有限公司。
离心机Super T21 Sorvall 美国Newton公司;均质机bilon-08 上海比朗仪器有限公司。
1.2 实验方法
1.2.1 菌种培养 将L.innocua保存在含0.6%的酵母提取物的胰酶解大豆酪蛋白胨琼脂培养基斜面中,储存在4℃下,每个月进行再次培养。接种前,使L.monocytogenes在150mL含0.6%的酵母提取物的大豆酪蛋白胨琼脂溶液中于37℃下培养24h,然后在4℃下用离心机以3300×g离心25min。倾倒出上清液,将细胞再次放入150mL 0.1%的蛋白胨水中以获得8lg CFU/mL的活性细胞群,接种到被污染的巴氏杀菌牛奶中。
1.2.2 Camembert奶酪的制备 本研究所有制备卡门贝尔奶酪的原料奶都经过巴氏杀菌,牛奶由超市购买,将24h菌龄的无害李斯特菌添加到牛奶中,使菌体含量达到之前所述的104~106CFU/mL。
1.2.3 制备奶酪所需的培养基和酶 奶酪制备用培养基从市场购买。培养基含有多种菌种:乳酸乳球菌乳酸亚种,乳酸乳球菌乳脂亚种,乳酸乳球菌乙酰乳酸亚种,明串珠菌乳脂亚种培养基储存在-20℃的干燥环境中。
Pencillium candidum作为制备卡门贝尔奶酪的霉菌培养基。霉菌培养基也同样储存在-20℃的干燥环境中。先将600mg的霉菌培养基在1L的牛奶中溶解后,再加入到22L的牛奶中。
1.2.4 实验室制备卡门贝尔奶酪 本实验中卡门贝尔奶酪制备需重复一次。在使用前,将直接接触牛奶,奶酪凝乳液和干酪的所有仪器(如大桶,刀具,模具纱布)及盐水,都经高压蒸汽灭菌以保证整个制备过程在无菌状态下操作。卡门贝尔奶酪的制备步骤如表1所示[6]。

表1 卡门贝尔奶酪典型制备及成熟数据列表
1.2.5 无害李斯特菌的计数 每一批实验,将三个含有无害李斯特菌的牛奶样品(1mL)分别放入装有20mL 0.1%无菌蛋白胨水中的100mL试管中。每块接种无害李斯特菌的奶酪样品放在400mL无菌振荡均质机内,并添加20mL 0.1%无菌蛋白胨水。所有样品在振荡均质机里混合2min。经0.1%无菌蛋白胨水连续稀释后,稀释液通过MOX平板计数。所有平板在30℃下培养24h。最后,计数平板上由无害李斯特菌形成的典型菌落。
1.2.6 实验设计 此实验制备圆柱形卡门贝尔奶酪块状的大小(直径108mm,高32mm),如图1所示。

图1 实验室制备卡门贝尔奶酪典型奶酪块大小
实验用的奶酪块状的形状和大小与市场上销售的卡门贝尔奶酪一致。一次实验平行制备三批奶酪(如图2所示),三批奶酪均使用含有无害李斯特菌的牛奶制备,根据成熟过程中的测定次数的不同,每批制备8块奶酪。为了验证圆柱型奶酪的轴对称性,将奶酪分割成120°的三个截面进行测量,成熟过程中分别对每块奶酪三个截面的无害李斯特菌进行计数。每个截面分成五个不同的测量位置,如图3所示。选择这些测量位置主要根据以下几分方面:a.根据Radhakrishnan[12]的初步测定结果,无害李斯特菌存活和生长有明显的区域差异;b.无害李斯特菌存活和生长的空间分布的确定,对数学模型的建立有积极的作用;c.成本和时间的约束。

图2 一批卡门贝尔奶酪实验设计方案
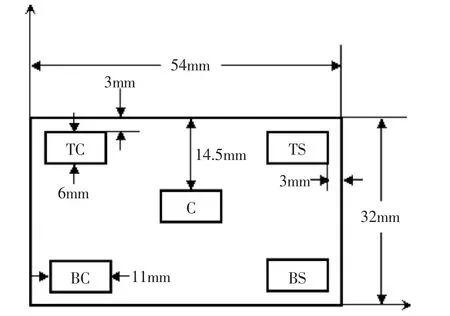
图3 测定无害李斯特菌存活和生长情况下样品取点位置
卡门贝尔奶酪成熟4到5周后可进行食用[12]。基于这些考虑,本实验关于无害李斯特菌存活和生长的实验设计方案如表2所示。

表2 无害李斯特菌存活和生长实验方案
1.2.7 统计分析 每次实验的数据都由Minitab software(Minitab 13,Minitab Inc.,State College,PA)进行统计分析,所有统计分析的检验水平为0.05。根据一般线性模型对数值进行比较,取测得的细菌数与标准偏差值的平均值。
2 结果与讨论
对三个批次的卡门贝尔奶酪成熟过程中的无害李斯特菌进行计数。其中每块奶酪有三个截面,五个测量位置。
2.1 卡门贝尔奶酪制备中无害李斯特菌的存活情况
卡门贝尔奶酪由原料奶和巴氏杀菌牛奶制备。本实验中所有的巴氏杀菌牛奶都接种经过24h培养的无害李斯特菌培养基。将1mL经过24h培养的菌体培养基添加到22L的巴氏杀菌牛奶后,牛奶中无害李斯特菌的细胞数大约保持在105CFU/mL。在制备卡门贝尔奶酪的过程中,因为起子培养基活性使pH下降,无害李斯特菌的生长得到抑制。但排出乳清后,无害李斯特菌在凝乳中得到了浓缩,结果导致干酪凝块中无害李斯特菌数目升高到107CFU/g,如表3所示。
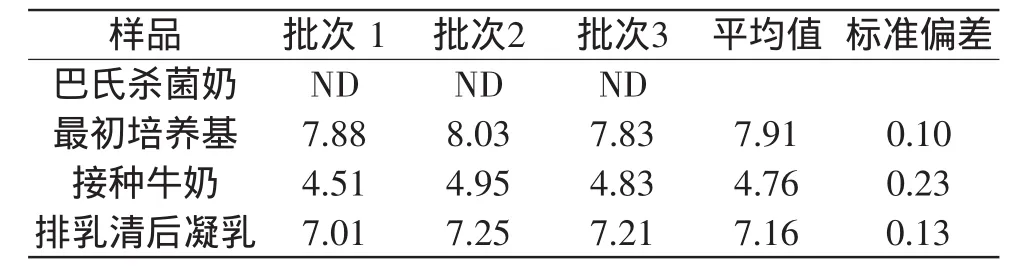
表3 奶酪制备过程中心上部(TC)无害李斯特菌菌落数情况(lg CFU/g)
2.2 卡门贝尔奶酪成熟过程中无害李斯特菌的存活情况
2.2.1 卡门贝尔奶酪成熟过程中无害李斯特菌存活的典型数据 成熟的前20d,所有奶酪块中的无害李斯特菌总数逐渐减少,这主要是起子培养基活性降低了pH。第20d后,无害李斯特菌总数开始增长,这可能是由于奶酪中pH的升高[6]。Ramsaran等人[13]研究表明在卡门贝尔奶酪表面附近,因蛋白质水解释放氨,pH较高,细胞存活性较高。图4为TC处无害李斯特菌存活和生长趋势。

图4 卡门贝尔奶酪成熟过程中TC位置无害李斯特菌存活和生长情况
通过卡门贝尔奶酪成熟过程中物理、化学和微生物性质的变化可以帮助理解无害李斯特菌的生长情况。成熟35d后,pH趋于一个平稳分布,这归因于霉菌独特的空间生长情况,霉菌在奶酪表面比在内部能更好地存活和生长。因此,奶酪表面的pH比中心的要高。此外卡门贝尔奶酪成熟过程中还存在蛋白质水解和酸化代谢[6]。
Lenoir[5]发现奶酪的表面特征导致P.camemberti的快速生长和表面霉菌产生的蛋白酶扩散到奶酪里。由于起子培养基和P.camemberti反应后的蛋白质水解,蛋白质水解或蛋白质降解产生了含氨物质,导致低分子含氮化合物的形成[5]。随着奶酪的成熟,在奶酪外皮和表面的氨浓度升高,这导致奶酪pH升高,同时乳酸代谢形成弱酸或其他化合物携带氢离子也会使pH升高。一般来说,低pH高温可以导致病微生物高效失活。在前20d的成熟过程中,无害李斯特菌的生长受到低pH和高温的影响。经过后15d的成熟,奶酪放在具有较低温度的冰箱中培养。细菌有了一个较利于存活和生长的环境。但因为细菌不断地恢复活性,细胞总数在第20d急剧下降。随着菌体活性的完全恢复,无害李斯特菌群落在后续的成熟过程中继续增加。
2.2.2 影响卡门贝尔奶酪成熟过程中无害李斯特菌数目的几个因素
2.2.2.1 位置比较 每天成熟过程中,奶酪不同位置处菌体数有显著性差异(P<0.05)。图5所示为5个菌数测定位置的菌体情况。
从测量数据可以看出,奶酪表面的菌落数(TS)高于奶酪中心的菌落数(见图5),这可能是因为奶酪中心的pH低而表面pH迅速升高的关系。TC,BC和 BS位置由于这三个位置相似的pH,无害李斯特菌生长速率相似。总之,奶酪块中细菌总数在前20d中下降1个对数数量级,在之后持续的成熟过程中升高1个对数数量级。因为pH低,奶酪中心位置的无害李斯特菌细胞生长率最低(C)。无害李斯特菌在中心上部位置(TC),中心下部位置(BC)和表面下部位置(BS)具有相似的生长速率,但TC处的无害李斯特菌菌落数比BC和BS位置的相对较高。这可能是因为在这些位置具有相似的pH[6]。总之,所有奶酪块的菌落数在前20d成熟过程中下降,而后在20~35d成熟过程中升高。
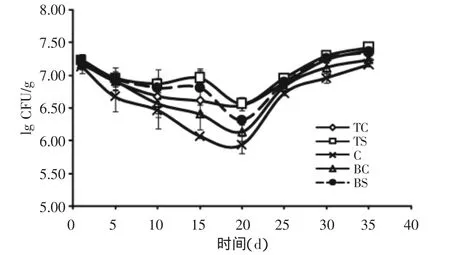
图5 卡门贝尔奶酪成熟过程中无害李斯特菌存活和生长情况
2.2.2.2 批次和截面比较 每天每批次和截面的数据没有显著性差异(P>0.05)。换句话说,在奶酪块中无害李斯特菌有相同的存活和生长趋势。结果显示,任何截面的测量都可代表奶酪块的测量数据。
2.2.2.3 成熟时间比较 不同的成熟时间对菌体数目存在显著性差异(P<0.05)。制备后,因低pH李斯特菌细胞数稍有降低。第1d所有奶酪块的细胞数平均值为7.16lg CFU/g。20d的成熟后降到了6.54lg CFU/g,随后在35d成熟时间后升高到7.38lg CFU/g(如图5所示)。无害李斯特菌在奶酪中不能生长的原因为pH较低,低pH降低了微生物的活菌数。在前20d的成熟过程中,奶酪中的pH低于5.5。在pH高于5.5时,无害李斯特菌菌数增加,这和之前报道过的文献一致。
3 结论
3.1 由于细胞浓缩,凝乳比接种过的牛奶具有更高的细胞数目。接种过无害李斯特菌的牛奶有细菌4.76lg CFU/g,浓缩成凝乳后升高到7.16lg CFU/g。
3.2 前20d成熟过程中,因奶酪块中的低pH,菌落数减少,其后在持续成熟过程中菌落数增加。
3.3 上表面区域(TS)具有较高的生长速率,然而,中心(C)区域生长速率较低。但是,中心顶部(TC),中心底部(BC)和表面底部(BS)区域具有相似的生长速率。
3.4 在批次和截面中无显著性差异。但时间和位置对无害李斯特菌的存活和生长有显著影响。
卡门贝尔奶酪成熟过程中无害李斯特菌存活和生长的数据可以建立一个动态预测模型。
[1]Curiale,M S,C Lewus.Detection of Listeria monocytogenes in samples containing Listeria innocua[J].J of Food Prot,1994,57: 1048-1051.
[2]Foegeding,P M,N W Stanley.Listeria innocua transformed with and antibiotic resistance plasmid as thermal-resistant indicator for Listeria monocytogenes[J].J of Food Prot,1991,54(7):519-523.
[3]Greenwood,M H D Robert,P Burden.The occurrence of Listeria species in milk and dairy products:A national survey in England and Wales[J].Int J Food Microbiology,1991,12:197-206.
[4]Laëtitia P,D Eliane,J C Cheftel.Inactivation of Listeria innocua in dairy fluids by pulsed electric fields:influence of electric parameters and food composition[J].Innovative Food Sci and Emerging Technology,2002(3):357-369.
[5]Lenoir J.The surface flora and its role in the ripening of cheese[J].Int Dairy Federation,1984,171:3-20.
[6]Liu S,V M Puri.Spatial pH distributions during ripening of Camembert cheese[J].Trans of the ASABE,2005,48:279-285.
[7]MacDonald F,A D Sutherland.Important differences between the generation times of Listeria monocytogenes and Listeria innocua in two Listeria enrichment broths [J].J of Dairy Research,1994,61:433-436.
[8]Manuela V,F Fátima,S Manuela,et al.Is Listeria innocua 2030c,a tetracycline-resistant strain,a suitable maker for replacing L.monocytogenes in challenge studies with coldsmoked fish?[J].Food control,2001(12):361-364.
[9]Margolles A,A Rodriguez,C G de los Reyes-Gavilán.Some chemical and bacteriological characteristics of regional cheeses from Asturias,Spain[J].J of Food Prot,1996,59:509-515.
[10]Margolles A,B Mayo,C G de los Reyes-Gavilán. Susceptibility of Listeria monocytogenes and Listeria innocua strains isolated from short-ripened cheeses to some antibiotics and heavy metal salts[J].Food microbiology,2001,18:67-73.
[11]Petran R L,K M J Swanson.Simultaneous growth of Listeria monocytogenes and Listeria innocua[J].J of Food Prot,1993,56: 616-618.
[12]Radhakrishnan S.Survival and growth modeling of Listeria monocytogenes during ripening of Camembert cheese[D].MS Thesis,Penn State University,University Park,PA.2000.
[13]Ramsaran H,J Chen,B Brunke,et al.Survival of bioluminescent Listeria monocytogenes and Escherichia coli O157:H7 in soft cheese[J].Journal of Dairy Science,1998,81: 1810-1817.
The spatial growth distribution of Listeria innocua under various conditions of Camembert cheese
HUANG Wei1,LIU Xue2,LIU Shao-wei1,*
(1.Eastern China University of Science and Technology,Shanghai 200237,China;2.China Agriculture University,Beijing 100083,China)
The ability of Listeria innocua to survive and grow during ripening of Camembert cheese was studied. Pasteurized whole milk containing about 5lg CFU/mL of L.innocua was used to elaborate in Camembert cheese.All cheeses were ripened in three stages:room temperature (about 20℃)and relative humidity 60% (36 hours),12℃and relative humidity 93%(2 weeks),7℃and relative humidity 85%(3 weeks).During ripening,surface and interior of cheeses were analyzed for population of L.innocua by surface plating on modified Oxford (MOX)agar.The data were collected on 1,5,10,15,20,25,30,35d during ripening.Results showed that population of L.innocua on the first day was around 7.16lg CFU/g which was 2.40lg CFU/g more than the initial inoculation of 4.76lg CFU/g.For subsequent 20 days,L.innocua population density declined to 6.54lg CFU/g. Thereafter,L.innocua population increased to 7.38lg CFU/g on 35d of ripening.Generally,the growth of L.innocua was faster in surface than in center location.
Listeria innocua;Camembert cheese;ripening;survival;growth
TS252.59
A
1002-0306(2011)10-0166-04
无毒害李斯特菌是一种革兰氏阳性、不产生孢子、杆状、嗜冷细菌、非溶血性及非致病的微生物[4]。两种李斯特菌物种可在相同的自然环境下生长,通常可从巴氏灭菌牛奶、软奶酪、日常产品和其他食品中分离出来。先前已有几篇文章研究了无害李斯特菌(L.innocua)的存活和生长情况。Margolles等[9]报道了在阿斯图里亚斯(西班牙北部)地区,可以从短期成熟的奶酪中分离出L.innocua品种。Greenwood等[3]研究指出在软质牛奶奶酪中屡次发现L.innocua和L.monocytogenes(单增李斯特菌)。L.innocua已被成功作为评估热处理过程对脱脂牛乳中L.monocytogenes的致死力的检测[2]。目前,还没有人研究在软奶酪成熟过程中L.innocua的空间存活和生长情况。本文用类似于商业产品形状的圆柱体Camembert奶酪来测量L.innocua的数量,此外,利用受污染的奶酪来测量L.innocua存活和生长的实际情况。因此,本文的研究目的一方面为测量Camembert奶酪在成熟过程中L.innocua存活和生长情况;另一方面评估和分析Camembert奶酪在成熟过程中L.innocua的空间分布状态。
2010-09-03 *通讯联系人
黄微(1986-),女,硕士,研究方向:低碳饮食。
上海市浦江人才计划项目(09PJ1403700);华东理工大学启动项目(YF0142122)。
