大竹蛏稚贝滤水率的研究
2011-09-24吴杨平陈爱华姚国兴张志伟吴建平
吴杨平, 陈爱华, 姚国兴, 张志伟, 吴建平
(江苏省海洋水产研究所, 江苏 南通 226007)
大竹蛏稚贝滤水率的研究
吴杨平, 陈爱华, 姚国兴, 张志伟, 吴建平
(江苏省海洋水产研究所, 江苏 南通 226007)
2008年10~11月间, 采用实验生态学方法对大竹蛏(Solen grandisDunker)稚贝进行了滤水率的研究。比较了不同饵料及浓度、规格、温度对大竹蛏稚贝滤水率(RF)的影响。结果表明: (1)在适宜浓度下,大竹蛏稚贝对5种单胞藻的滤水率均随着浓度的增大而增大; (2)大竹蛏稚贝的滤水率随着体质量的增大而增大, 符合幂函数回归关系:RF=13.029WD0.7309(R2=0.998 8); (3)在一定温度范围内, 随温度升高大竹蛏稚贝滤水率增加; 超过一定温度范围, 随着升高滤水率下降; 当温度为 23℃时, 滤水率达到最大值(1.38±0.01)mL/(个•min)。
大竹蛏(Solen grandisDunker); 滤水率; 规格; 饵料种类; 饵料浓度; 温度
滤水率是滤食性双壳类的重要生理指标之一,是双壳类能量学的基本参数, 可用于推算双壳类在特定海区的生长余力(SFG: Scope for growth) , 是进行养殖容量评估所必须的参数。滤水率反映了贝类生理状态的动态变化, 受环境因素和生物因素的影响较大, 例如水温、饵料浓度和质量、贝类的规格、盐度、pH值、以及海水流速等因素。国内外有关滤食性双壳类滤水率方面已有不少研究, 如在贻贝[1]、青蛤[2]、毛蚶[3]、缢蛏[4]、海湾扇贝和太平洋牡蛎[5]等贝类都已有相关报道, 但有关大竹蛏特别是稚贝滤水率方面尚未见研究报道。
大竹蛏(Solen grandis)个体大, 营养丰富, 是重要的经济贝类, 分布于帝汶岛、菲律宾、泰国湾、朝鲜西海岸、日本房总半岛以南以及我国各海区[6]; 是一种营埋栖生活、主要通过滤水作用摄食海水中浮游生物和有机碎屑的双壳纲(Bivalvia)、异齿亚纲(Heterodonta)、竹蛏科(Solenidae)贝类。2007年江苏省海洋水产研究所在国内首次实现了大竹蛏大规模生产性人工繁殖技术的突破, 培育规格 1.0~1.5 cm大竹蛏稚贝2 000余万粒[7], 不仅实现了大竹蛏增殖放流的生态效益, 而且有利推动了大竹蛏人工养殖的开展。目前大竹蛏养殖业的发展迫切需要滤水率方面的基础研究予以支持, 为苗种培育、人工养殖和增殖提供基本数据和理论指导。作者主要研究了饵料种类及浓度、规格和温度对大竹蛏稚贝滤水率的影响。
1 材料与方法
1.1 实验材料
实验用稚贝是2008年由江苏省海洋水产研究所江苏文蛤良种场育苗车间经催产、孵化及人工培育等一系列措施后获得的大竹蛏稚贝。实验所用单胞藻为海水小球藻(MarineChlorellasp.)、绿色巴夫藻(Pavlova viridis)、亚心形扁藻(Platymonas subcordiformis)、叉鞭金藻(Dicrateriasp.)和微绿球藻(Nannochloropsis oculata), 藻类培养用砂滤海水经高温杀菌消毒后使用, 各种单胞藻均用陈爱华等[8]所用方法培养。
1.2 实验方法
1.2.1 饵料种类和浓度对大竹蛏稚贝滤水率影响实验
每1 000 mL烧杯中放大竹蛏稚贝10粒(A组规格), 实验设3个平行组和1个空白对照组(不放大竹蛏稚贝观察单胞藻的变化情况)。预实验获得产生假粪的藻类阈值浓度以后, 在此浓度以下设置各种单胞藻的浓度梯度。海水小球藻和微绿球藻的浓度梯度为 20×104、30×104、40×104、50×104和 60×104个/mL; 绿色巴夫藻和叉鞭金藻为 5×104、10×104、15×104、20×104和 25×104个/ mL; 扁藻为 2×104、4×104、6×104、8×104和 10×104个/ mL, 实验水温为26 ℃ ,盐度为 23.7, 光照为自然光照。实验开始时和60 min后直接用血球计数板计数藻类浓度, 根据浓度变化计算滤水率。
1.2.2 体质量对大竹蛏稚贝滤水率影响实验
取4种不同规格大竹蛏稚贝分4组(A、B、C、D)研究体质量对大竹蛏稚贝滤水率影响。实验装置设计、水温、盐度同1.2.1。投喂叉鞭金藻10×104个/ mL。大竹蛏稚贝规格见表1。
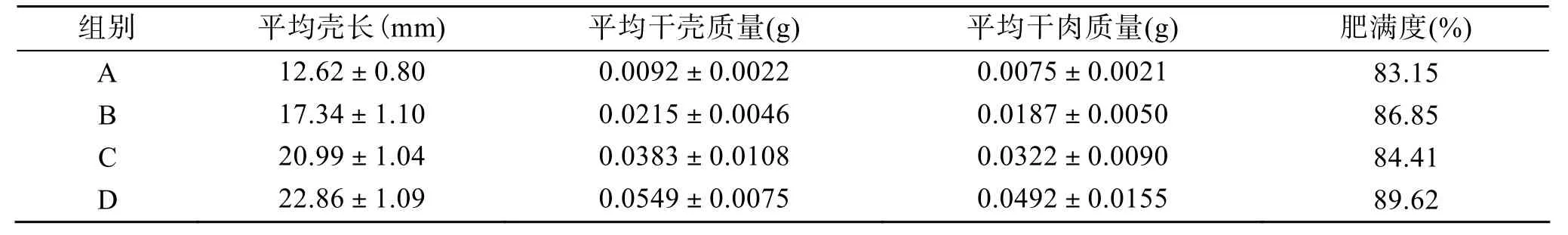
表1 大竹蛏稚贝生物学测定Tab. 1 Biological data of juvenileSolen grandis
1.2.3 温度对大竹蛏稚贝滤水率影响实验
实验设6个温度梯度: 9、14、17、20、23、26℃。室温9℃, 采用加热棒水浴调温至设定温度并控温。实验用大竹蛏稚贝为 D组规格, 投喂单胞藻为叉鞭金藻10×104个/ mL, 装置同1.2.1。
1.3 指标计算
大竹蛏滤水率(Filtration rate,RF)的测定计算公式[8]如下:

式中,C0代表起始藻类的浓度,Ct代表结束时藻类的浓度,V指实验水的体积,N代表实验贝的个数,t指实验时间。
1.4 统计分析
实验数据用SPSS16.0软件进行统计分析。
2 结果与分析
2.1 不同单胞藻种类及其浓度对大竹蛏稚贝滤水率的影响
结果表明, 大竹蛏稚贝对 5种单胞藻的滤水率均随着浓度的增大而增大, 当达到一定浓度时滤水率反而下降。如图1所示, 在浓度为30×104个/mL时, 大竹蛏稚贝对海水小球藻的滤水率最大, 为(1.93±0.09)mL/(个·min), 而微绿球藻在浓度为 50×104个/mL 时滤水率最大, 为(2.34±0.11)mL/(个·min)。方差分析表明, 在藻细胞浓度为 30×104个/mL时, 大竹蛏稚贝对海水小球藻和微绿球藻的滤水率无显著性差异(P>0.05), 在其他浓度下, 滤水率差异显著(P<0.05)。
从图 2可以看出, 在藻细胞密度为 10×104和15×104个/mL时, 大竹蛏稚贝对绿色巴夫藻和叉鞭金藻的滤水率基本相同(P>0.05), 当藻细胞密度为15×104个/mL时, 对绿色巴夫藻和叉鞭金藻的最大滤水 率 分 别 为 (1.29±0.09)mL/(个 ·min)和 (1.32±0.08)mL/(个·min)。在此密度下, 大竹蛏稚贝对两种单胞藻的滤水率无显著性差异(P>0.05)。图3表明, 在密度为6×104个/mL时, 大竹蛏稚贝对扁藻的滤水率达到最大, 为(1.32±0.06)mL/(个·min)。综合图 1、图 2 和图 3,在藻类密度为20×104个/mL时, 大竹蛏稚贝对海水小球藻和微绿球藻的滤水率明显大于对绿色巴夫藻和叉鞭金藻的滤水率(P<0.05); 而当藻类密度为 10×104个/mL时, 大竹蛏稚贝对亚心形扁藻的滤水率与对绿色巴夫藻和叉鞭金藻的滤水率无明显差异(P>0.05)。

图1 大竹蛏稚贝对海水小球藻和微绿球藻的滤水率Fig. 1 Effect of marineChlorellasp. orNannochloropsisoculataon the filtration rate of juvenileS. grandis
2.2 大竹蛏稚贝规格和滤水率的关系
在相同温度和饵料密度的情况下, 大竹蛏的滤水率RF与其软体部干质量存在幂函数关系:RF=13.029WD0.7309,R2=0.998 8, 即滤水率随着大竹蛏壳长、体质量的增加而增加(图 4), 体质量对其滤水率的影响显著(P<0.05)。
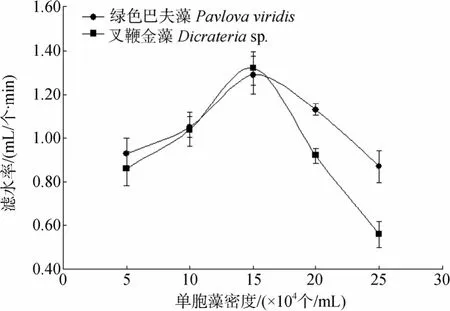
图2 大竹蛏稚贝对绿色巴夫藻和叉鞭金藻的滤水率Fig. 2 Effect ofPavlova viridisorDicrateriasp. on the filtration rate of juvenileS. grandis

图3 大竹蛏稚贝对亚心形扁藻的滤水率Fig. 3 Effect ofPlatymonas subcordiformis. on the filtration rate of juvenileS. grandis

图4 大竹蛏稚贝干肉质量与其滤水率的关系Fig. 4 The correlation of filtration rate and dry weight of juvenileS. grandis
2.3 温度对大竹蛏稚贝滤水率的影响
不同温度条件下大竹蛏稚贝的滤水率如图 5所示。低温(9~17℃)时温度变化对滤水率的影响较小,当温度为 23℃时滤水率达到最大值(1.38±0.01)mL/(个·min), 然后随着温度上升滤水率呈下降趋势。方差检验表明温度变化对大竹蛏稚贝滤水率影响显著(P<0.05)。

图5 温度对大竹蛏稚贝滤水率的影响Fig. 5 Effect of temperature on filtration rate of juvenileS. grandis
3 讨论
3.1 饵料种类及密度对大竹蛏稚贝滤水率的影响
饵料作为影响贝类摄食生理的关键因子之一,其种类和密度对贝类滤水率都有不同程度的影响。在适宜饵料密度范围内, 大竹蛏稚贝对 5种藻类的滤水率随密度增高而增高, 超过一定范围, 密度增高, 滤水率下降, 这与青蛤[2]、大西洋浪蛤[9]等贝类滤水率研究的报道相似。Risgard[1]认为饵料密度过高或过低, 滤水率会受到影响。Robinson等[11]的研究也表明水中高密度悬浮物会降低大西洋浪蛤对海藻的摄入和消化。沈和定等[3]分析藻类密度过高导致滤除率下降的原因时, 认为高密度下饵料颗粒在鳃表面的运动速度减慢, 易造成阻鳃现象, Ward[10]等亦有同样认识。另一方面, 饵料密度过高, 贝类易产生假粪, 对自身的生长不利, 本实验所设密度严格控制在产生假粪的阈值以下, 没有发现假粪的产生。因此, 在实际生产过程中, 一次投饵量不宜过高, 应遵循少投多喂的原则。
王如才等[12]、沈和定[3]、王慧等[2]的研究认为,滤食性贝类对食物的大小和质量有选择性, 但对食物是否有食用价值没有选择性。本实验对 5种单胞藻进行了研究, 绿色巴夫藻(Φ=6 μm)和叉鞭金藻(Φ=6~7 μm)、海水小球藻(Φ=3~5 μm)和微绿球藻(Φ=2~3 μm)、亚心形扁藻(Φ= 11~16 μm)。由图 1 和图 2可见大竹蛏稚贝对粒径相当的单胞藻的滤水率相近, 即表现为对巴夫藻和叉鞭金藻的滤水效果相当, 对小球藻和微绿球藻的滤水效果相当; 当藻类密度都是20×104个/mL时, 大竹蛏稚贝对微绿球藻的滤水效果最好, 小球藻次之, 巴夫藻第三, 叉鞭藻最低。此结果与青蛤对粒径较大藻类滤水效果最好的报道[2]有异, 这可能是由于大竹蛏稚贝鳃未发育完全, 摄取机制以黏液纤毛作用为主, 在此作用下可有效运输较小规格的单胞藻, 因而表现为对较小规格的单胞藻的较高选择性[5]。大竹蛏稚贝的滤食机制以及这种机制与其自身结构特点有何关系还有待进一步的研究。
3.2 体质量对大竹蛏稚贝滤水率的影响
多数研究结果表明海洋双壳滤食性贝类与体质量之间存在一定关系, 即滤水率随着体质量增大而增大, 可用公式表达为RF=aWb, b值在0.60~0.94之间[13]。本实验通过一次性添加饵料法得出关系式中的b值是0.7309, 在此范围内, 与其他多数已报道的双壳类一致。而对于单位干组织质量而言, 其滤水率为RF= aWb/W= aWb-1(b-1<0), 即个体越大, 单位干组织质量的滤水率越小, 从而也可以说明贝类可以通过调节滤水率来满足不同生长阶段的营养需求。方建光等[14]研究发现泥蚶稚贝的滤食能力大大强于浮游幼虫, 也正说明了贝类通过对滤水率的调控来满足不同生长发育时期对营养物质的需求。因此, 在进行大竹蛏稚贝苗种培育时, 应根据这一特点来调节饵料供应量以适应其生长。
3.3 温度对大竹蛏稚贝滤水率的影响
实验结果显示随着温度逐步升高, 大竹蛏稚贝滤水率有个先增后减的变化过程。23℃是大竹蛏稚贝最适宜的温度, 此时, 滤水率达到最大值(1.38±0.01)mL/(个·min)。这种温度对滤水率的影响变化关系在海湾扇贝[5]、太平洋牡蛎[5]、毛蚶[3]等贝类上均有体现。对此, Jorgensen等[15]在研究偏顶蛤(Modiolus modiolus)时认为, 滤食性贝类的滤水率在一定温度范围内随温度的升高而加大, 一方面温度和贝类鳃上侧纤毛的摆动有正相关性, 温度升高使纤毛的摆动频率加快; 另一方面高温可以减小海水的黏滞性, 从而增加滤水率[16]。王芳等[5]分析认为这可能是因为在适宜的温度范围内, 温度升高使贝类的代谢加强, 滤食加快, 滤除率明显, 而当水温超过一定范围时, 贝类处于不正常的生理状态, 导致滤水率的下降。潘鲁青等[4]也认为当温度超出适宜范围时, 贝类则要通过改变代谢状况, 消耗更多的能量来适应外界环境的变化, 就会降低摄食器官的活力,从而导致滤水率的下降。由图5所示, 温度小于15℃时, 大竹蛏稚贝滤水率相对较低, 且随温度升高, 增加的幅度很小; 而当高于15℃后, 滤水率显著增加。这说明 15℃是一个临界点, 低于 15℃时, 大竹蛏稚贝生物活性弱, 只需通过滤水作用摄食少量饵料即可维系基本存活所需; 当温度达到 15℃以后, 生命活动开始变得旺盛, 即表现为稚贝通过增强滤水来满足用于生长的饵料需求, 这与陈爱华等[17]有关15℃ 以下大竹蛏苗种生长滞缓的报道相一致。
[1] Risgard H U, Randlov A, Knstensen P A.Rates of water processing oxygen consumption and efficiency of particle retention in veliger and young post-metamorphicMytilus edulis[J].Ophelia, 1980, 19(1): 37-46.
[2] 王慧, 房斌, 周凯等.不同藻类及浓度对青蛤滤水率的影响[J].海洋渔业, 2008, 30(1): 92-96.
[3] 沈和定, 张饮江, 卢华春等.毛蚶对单胞藻的滤除率、选择性和消化状况[J].上海水产大学学报, 2006,15(2): 195-200.
[4] 潘鲁青, 范德明, 董双林等.环境因子对缢蛏滤水率的影响[J].水产学报, 2002, 26(3): 226-230.
[5] 王芳, 董双林.海湾扇贝和太平洋牡蛎的食物选择性及滤除率的实验研究[J].海洋与湖沼, 2000, 31(2):139-144.
[6] 齐钟彦.中国经济软体动物[M].北京: 中国农业出版社, 1998: 233.
[7] 陈爱华, 张志伟, 姚国兴等.环境因子对大竹蛏稚贝生长及存活的影响[J].上海水产大学学报, 2008,17(5): 259-263.
[8] 陈爱华, 姚国兴.单细胞藻类规模化培养技术要点[J].水产养殖, 2006, 27(5): 20-22.
[9] 郭海燕, 王昭萍, 于瑞海等.饵料密度、温度和体质量对大西洋浪蛤滤水率的影响[J].海洋科学, 2005,29(8): 1-3.
[10] Ward J E, Macdonald B A, Thompson R J. Mechanisms of suspension feeding in bivalves: Resolution of current controversies by means of endoscopy [J]. Limnol Oceanogr, 1993, 38(2): 265-272.
[11] Robinson W E, Wehling W E, Morse M P. The effect of suspended clay on feeding and digestive efficiency of the surf clam, Spisula solidissima(Dillwyn)[J]. Exp Mar Biol Ecol, 1984, 74: 1-12.
[12] 王如才, 王昭萍, 张建中.海水贝类养殖学[M].青岛: 青岛海洋大学出版社, 1993. 40.
[13] 张继红, 方建光.海洋双壳贝类滤水率测定方法概述[J].海洋水产研究, 2005, 26(1): 86-93.
[14] 方建光, 孙慧玲.泥蚶幼虫滤水率和摄食率的研究[J].海洋与湖沼, 1999, 30(2): 167-171.
[15] Jorgensen C B, Larsen P S, Rissgard H U.Effects of temperature on the mussel pump[J].Mar Ecol Prog Ser,1990, 64: 89-97.
[16] Bernard F R.Nutrition ofCrassostrea gigas(Thunberg,1795): an aspect of estuarine energetics[D].Ph D Thesis, University of London, 1972: 448.
[17] 陈爱华, 姚国兴, 张志伟.大竹蛏生产性人工繁育试验[J].海洋渔业, 2009, 31(1): 66-72.
Received: Jul., 16, 2010
Key words:Solen grandis; filtration rate; sizes; diet species; diet density; temperature
Abstract:The effects of temperature, diets, diet density, and sizes on the filtration rate ofSolen grandiswere studies in a laboratory system from October to November 2008. It was found that the filtration rate of five species microalgae was increased with microalgae density increasing at the optimum microalgae density. The relationship between filtration rates and individual weight followed an exponential correlation,RF=13.029WD0.7309(R2=0.998 8).In addition, the filtration rate was increased with the increase of temperature, but was decreased if the temperature exceeded 23℃, at which the filtration rate reached the maximum of (1.38±0.01) ml /(ind·min).
(本文编辑:梁德海)
The filtration rate of juvenileSolen grandisDunker
WU Yang-ping, CHEN Ai-hua, YAO Guo-xing, ZHANG Zhi-wei, WU Jian-ping
(Jiangsu Institute of Marine Fisheries, Nantong 226007, China)
S968.3
A
1000-3096(2011)01-0006-04
2010-07-16;
2010-08-19
江苏省水产三项工程项目(K2008-1); 南通市农业创新科技计划项目(AL2009001)
吴杨平(1982-), 男, 江苏南通人, 助理研究员, 主要从事海洋生物生理生态研究, E-mail: uyping@yahoo.cn; 陈爱华, 通信作者,E-mail: chenah540540@yahoo.com.cn
