低温与光照对茶树叶片叶绿素荧光参数的影响
2011-09-18孔海云张丽霞王日为
孔海云 张丽霞* 王日为
(1.山东农业大学园艺科学与工程学院,山东泰安 271018;2.作物生物学国家重点实验室,山东泰安 271018;3.山东农业大学应用化学与材料科学学院,山东泰安 271018)
所谓光抑制是指光合结构吸收的光能超过光合作用本身所利用的能量而引起光合效率下降的现象。长时间的光抑制可引起光合机构的光氧化破坏。Hodgon等[1]将植物在低温下发生的光抑制称为低温诱导光抑制(chill-induced photoinhibition)或低温光抑制(chill-temperature photoinhibition)。低温是影响许多植物产量和地理分布的一个主要环境因素。起源于热带和亚热带的植物对温度反应尤为敏感,当遇到稍低于其适宜生长温度时,就会受到低温伤害,植物发生冷害的临界温度一般在10~12℃,当温度降至引起冷害的临界温度时,光合作用就会受到抑制,而且低温会增加冷敏感植物和抗冷植物发生光抑制的可能性[2],在低温下冷敏感植物对光强特别敏感,中等光强即可引起严重的光抑制。
叶绿素荧光动力学是以光合作用理论为基础,利用叶绿素荧光作为天然探针[3],研究和探测植物光合生理状况及各种外界因子对其细微影响的新型植物活体测定和诊断技术。叶绿素荧光不仅能反映光能吸收、激发能传递和光化学反应等光合作用的原初反应过程,而且与电子传递、质子梯度的建立及ATP合成和CO2固定等过程有关。几乎所有光合作用过程的变化均可通过叶绿素荧光反映出来,而荧光测定技术不需破碎细胞,不伤害生物体,因此,该技术具有快速、对环境变化十分灵敏和非破坏性等优点,是研究植物光合作用的良好探针。
近年来,国内外利用叶绿素a荧光动力学方法研究了温度、水分等环境因子对多种植物光合作用的影响。而对茶树的研究主要集中在水分胁迫、高温、强光等方面,有研究表明光照可以加速低温引起的茶树光合作用的降低,但应用叶绿素a荧光参数来研究和探测低温对茶树发生光抑制的研究还未见报道。因此,本试验通过研究茶树在不同低温光照下叶绿素荧光参数的变化,从而探明低温光照对茶树叶片发生光抑制的影响,以期为山东冬季茶树栽培管理提供理论依据。
1 材料与方法
1.1 供试材料
供试材料为健壮、长势较一致的福鼎大白一年生无性良种茶籽实生苗,栽种在高242 mm、口部直径161 mm、底部直径95 mm的塑料钵中,栽培基质配方为草炭∶珍珠岩∶石英砂=2∶1∶1。放置于玻璃温室内培养,期间养分、水分供应充足。
1.2 试验设计
设温度和光照两因素5水平试验,其中温度设为:0、5、10、15℃和25℃,其中25℃作 CK,光照强度设为:3、4、6、10、15.5 klx,共计 25 个处理。
1.3 测定方法
1.3.1 材料预处理 将盆栽的茶苗在光照培养箱内预培养12 h后,每处理各选取10片生理状态较一致成熟叶片(做标记)进行叶绿素荧光参数测定。
1.3.2 叶片叶绿素荧光参数的测定及计算方法采用英国Hansatech公司生产的FMS-Ⅱ便携调制式荧光仪测定叶绿素荧光参数。将茶苗叶片暗适应15 min后,采用Fv/Fm模式测定暗适应下茶树功能叶片的荧光参数,先照射弱检测光0.12 μmol·m-2·s-1,测得初始荧光(Fo),再照射饱和脉冲光4000 μmol·m-2·s-1,测得最大荧光(Fm),然后计算出暗适应最大可变荧光(Fv=Fm-Fo)、暗适应下PSⅡ最大光化学效率(Fv/Fm)、PSⅡ潜在活性(Fv/Fo);再照射作用光后,分别依次照射检测光和饱和脉冲光,测得作用光存在时的最大荧光(Fm'),关闭作用光后,立即照射远红光 1.67 μmol·m-2·s-1,测得作用光关闭后初始荧光(Fo'),光化学猝灭系数(qP)及稳态下荧光(Fs),然后计算出光适应下可变荧光(Fv'=Fm'-Fo')、光适应下PSⅡ最大光化学效率(Fv'/Fm')、PSⅡ实际光化学效[ΦPSII=(Fm'-Fs)/Fm']、非光化学猝灭系数[qN=(Fm-Fm')/(Fm -Fo')]。
2 结果与分析
2.1 低温光照对茶树叶片Fo、Fm的影响
基础荧光Fo表示PSⅡ反应中心全部开放即原初电子受体QA全部氧化时的荧光水平;最大荧光Fm是PSⅡ反应中心完全关闭时的荧光产量,反映了通过PSⅡ的电子传递情况。从图1-A可以看出,在25℃下幼苗叶片Fo几乎处在同一水平,变化幅度极小,但在低温胁迫下呈现递增趋势,且在同一光照强度下,随着处理温度的降低,Fo逐渐升高。表明低温光照使PSⅡ反应中心破坏或可逆失活。图1-B中Fm在25℃下变化不明显,随着光照的增强呈略微上升趋势,但在低温10、5、0℃胁迫下呈现明显递减趋势,光照越强,则下降幅度越大;另外,在5℃和0℃、光照在3~10 klx时,Fm急剧下降,随后下降趋势缓慢。
2.2 低温光照对茶树叶片Fv/Fm、Fv/Fo的影响
Fv/Fm和Fv/Fo的比值分别代表PSII原初光能转化效率和PSⅡ的潜在活性,Fv/Fm下降是光合作用光抑制的重要特征。从图1-C可以看出,Fv/Fm值在25℃条件下,基本维持在0.83左右,保持稳定水平;当光照为3 klx,在不同低温下,该参数下降幅度极小,基本保持在0.80左右,光抑制不明显,随着光照的增强,Fv/Fm下降越明显;在0℃、3 klx~10 klx之间呈现明显的急剧下降趋势,而10 klx以后,下降趋势略缓,这可能与此时茶苗已经发生冻害,反应机制受损有关;在0℃、15.5 klx时,Fv/Fm下降到0.60,光抑制程度最大;由图1-D可知,Fv/Fo变化趋势与Fv/Fm相似。这说明低温光照降低了茶苗PSⅡ的原初光能转化效率和潜在活性,使茶树PSⅡ反应中心受到伤害。
2.3 低温光照对茶树叶片ΦPSII、Fv'/Fm'的影响
ΦPSII常用来表示植物光合作用电子传递的量子产额,可作为植物叶片光合电子传递速率快慢的相对指标;Fv'/Fm'代表了光适应下PSII最大光化学量子产率,或者表征在实际光适应条件下PS反应中心原初光能捕获效率,它反映有热耗散存在时PSII反应中心完全开放时的光化学效率。从图1-E可以看出,ΦPSII值在25℃条件下,随着光照的增强而呈上升趋势,但在低温下,随着光照的增强而下降,且在同一光照下,随着温度降低而下降;由图1-F可知,Fv'/Fm'变化趋势与ΦPSII相似,且在同一光照下,随着温度下降,Fv'/Fm'下降幅度较大。由以上分析可知,低温光照降低了茶苗叶片光合电子传递速率及光适应下PSII最大光化学效率。
2.4 低温光照对茶树叶片qP、qN的影响
荧光猝灭是叶绿体耗散能量的一种途径,分为光化学猝灭qP和非光化学猝灭qN两种。从图1-G可以看出,qP在25℃下,随着光照的增强而上升,但在低温胁迫下呈现明显下降趋势,在0℃条件下降幅度最大,且在同一光照下,随着温度的降低而下降,同时,光照越强,下降幅度越大;从图1-H可以看出,qN值与qP值变化趋势相反,在25℃条件下,随着光照的加强而降低,而在低温胁迫下呈上升趋势,且在同一光照条件下,随着温度的降低而上升。
3 讨论
叶绿素荧光参数Fv/Fm在非胁迫条件下变化极小,一般在0.83左右[4],不受物种和生长条件的影响,而胁迫条件下该参数明显下降。因此,在光抑制中常以叶绿素荧光Fv/Fm比值的下降作为衡量光抑制的指标。本研究结果表明:茶树叶片在25℃下,Fv/Fm基本维持在0.83左右,保持稳定水平,未受到光抑制;在15℃下,Fv/Fm有轻微下降趋势,随着处理温度的下降,该参数在不同光强下呈现不同程度的下降,且温度越低,茶苗产生光抑制所需的光强越小。这说明低温诱导了光合作用光抑制,这可能与低温降低了植株 1,5-二磷酸核酮糖羧化酶/加氧酶(Rubisco)、果糖-1,6-二磷酸酶(FBPase)和景天庚酮糖-1,7-二磷酸酶(SBPase)等碳代谢酶的活性以及核酮糖1,5-二磷酸(RuBP)再生速率有关[5]。
低温光照下荧光产量显著降低,表明低温光照胁迫使茶树叶片PSⅡ活性中心受损,光合作用原初反应过程受抑制,光合电子由PSⅡ反应中心向质体醌(QA、QB及PQ)库传递过程受到影响,不利于激发能从天线色素蛋白复合体(LHC)向PSⅡ传递。本研究的结果表明,低温光照能使荧光参数ΦPSⅡ下降,表明低温胁迫使光合电子传递过程受抑制和损伤,光合电子传递速率下降,光合作用强度减弱。这与在杨梅[6]、黄瓜[7]、辣椒[8]、甜瓜[9]、西瓜[10]等植物方面的研究结果一致。
随着低温光照逆境程度的加剧,qP呈现下降趋势而qN呈上升趋势。qP与PSII的光化学反应有关,反映了PSⅡ原初电子受体QA的氧化还原状态,qP下降表明低温光照下PSII的电子传递活性减小,从而使茶树光合碳同化能力受到影响;qN指由非辐射能量耗散等引起的荧光猝灭,qN上升说明茶树幼苗通过耗散多余能量以保护光合机构免受低温伤害的能力增强。
综上所述,本模拟试验通过对茶苗在不同低温光照下叶绿素荧光参数变化的研究,探明了茶苗在不同低温及光强条件下发生光抑制的情况。这为合理判断北方茶区秋冬及早春低温条件下发生光抑制和光损伤的情况提供理论依据,进一步指导北方茶树安全越过低温期。
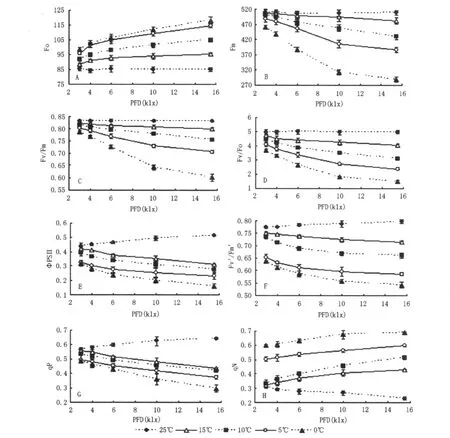
图1 不同低温光照对茶苗叶绿素荧光参数的影响Fig 1 Effects of different low temperature and light on chlorophyll fluorescence kinetics Parameters in tea seedlings
1 Hodgon R.A.J.,Orrg R.,Raison J.K..Inhibition of photosynthesis by chilling in the light[J].Plant.Sci.,1987,49:75 - 79.
2 Hetherington S.E.,He J.,Smillie R.M..Photoinhibition at low temperature in chilling sensitible and resistant plants[J].Plant Physiol.,1989,90:1609 -1615.
3 张守仁.叶绿素荧光动力学参数的意义及讨论.植物学通报,1999,16(4):444 -448.
4 胡文海,黄黎锋,肖宜安.夜间低温对2种光强下榕树叶绿素荧光的影响.浙江林学院学报,2005,22(1):20 -23.
5 Hutchison R.S.,Groom Q.,Ort D.R..Differential effects of chilling induced photooxidation on the redoxregulation of photosynthesis enzymes.Biochemistry,2000,39:6679 -6688.
6 刘 辉,郭延平,胡美君.杨梅光合作用的低温光抑制.热带亚热带植物学报,2005,13(4):338 -342.
7 陈启林,山 仑,程智慧.低温下光照对黄瓜叶片光合特性的影响.中国农业科学,2001,34(6):632 -636.
8 张俊峰,颉建明,郁继华.低温对辣椒幼苗光化学效率的影响.甘肃农业大学学报,2009,44(1):89 -92.
9 和红云,薛 琳,田丽萍.低温胁迫对甜瓜幼苗叶绿素含量及荧光参数的影响.北方园艺,2008,(4):13 -16.
10 刘慧英,朱祝军,史庆华.低温胁迫下嫁接对西瓜光合特性及叶绿素荧光参数影响的研究.石河子大学学报,2007,25(2):163-166.
