细胞重编程:调节关键基因获得需要细胞
2011-08-09庞希宁
庞希宁
中国医科大学细胞生物学卫生部重点实验室干细胞与再生医学研究室,沈阳 110001
细胞发育和分化的内部编程是遵照细胞内基因 表达规律进行的。每个细胞都有全能的细胞核及相同的分化潜能。在胚胎发育不同阶段,由于细胞所处微环境不同及细胞定向分化内部编程不同,基因表达就存在差异,即开放某些基因,关闭某些基因,以使细胞合成特异性的蛋白质,产生不同的结构、功能及表型。基因组DNA在细胞分化过程中不是全部表达,而是基因的差异表达,即奢侈基因按一定程序有选择地相继活化表达。调控细胞分化的基因编程是由不同信号分子在特定时间和空间作用于细胞,产生基因表达的内在规律。总之,如能改变基因的表达就能改变细胞的编程,从而改变细胞的分化方向。
细胞重编程是在一定条件下成体细胞的记忆被擦除,重新程序化产生新的表型和功能,导致细胞的命运发生改变。细胞重编程主要发生在不涉及基因组DNA序列改变的基因表达水平。对于细胞重编程的深入研究有助于掌握机体细胞的发生发育机制,为解决再生医学种子细胞来源问题提供理论基础[1]。
细胞重编程的种类
细胞重编程主要分为两类:(1)分化的成体细胞在特定条件下被逆转后恢复到全能性或多能性状态,形成新的多能性干细胞[2],如诱导多潜能干细胞 (induced pluripotent stem cells,iPSCs)。近年来,iPSCs研究为基因调控去分化和转分化提供了分子实验依据,使有目的地调控细胞基因的表达,改变细胞内部编程来改变原来分化方向成为现实。(2)一种胚层来源的细胞或多能性干细胞向同胚层或不同胚层来源的另一种成体细胞或多能性干细胞转化,也称转分化 (transdifferentiation)[3]。
影响细胞重编程的因素
研究表明,基因表达不但受转录因子的调控,还与其DNA和组蛋白表观遗传学修饰有关[4],包括DNA甲基化、组蛋白乙酰化、印记基因表达、端粒长度恢复、X染色体失活等[5]。在真核生物基因组的非蛋白质编码区存在大量非编码RNA(non-coding RNA,ncRNA)基因,这些非编码区域担负着基因表达调控等重要功能,其编码产物可在转录后水平调节靶基因的表达,是调控细胞内基因表达的基本机制之一。ncRNA不仅在干细胞的多能性维持过程中有重要作用,在成体细胞重编程中也发挥重要作用[6],还参与了干细胞分化的调节[7]。
细胞重编程主要针对定向分化的某些关键调节基因,特别是近年发现的某些因子能调控大量基因的表达,对细胞编程起着重要作用。例如:神经元限制性沉默因子 (RE1-silencing transcription factor/neuron restrictive silencer factor,REST/NRSF)通过与调控基因启动子的一段21nt的DNA保守序列—神经元限制性沉默元件 (neuron restrictive silencer element/repressor element 1,NRSE/RE-1)结合,经一系列反应使组蛋白发生甲基化和去乙酰化修饰,使染色质呈凝缩状态,启动子区域无法和转录因子及RNA聚合酶结合,而抑制其转录活性。这种表观遗传学修饰可批量抑制上千个与神经细胞分化相关的基因,其抑制的解除是神经细胞分化的必要条件[8]。最近研究表明,胰岛 β细胞分化基因胰十二指肠同源框1(Pdx1)、胰岛素和神经原质蛋白3(Ngn3)、神经源性分化蛋白1(NeuroD1)和成对盒4(Pax4)均因具有NRSE序列而受REST/NRSF调节[9-11]。Pdx1主要在胰腺前体细胞中表达,是促进早期胰腺发育以及胰岛 β细胞成熟的关键基因,是最受关注的具有正向调节作用的转录激活因子,能和众多与胰岛分化有关的靶基因启动子中的TAAT序列结合,从而在启动胰岛内分泌细胞分化过程中发挥重要的作用[12]。
随着发育分子生物学研究的深入,许多组织细胞的发育机制研究不断深入,提供了大量细胞分化的分子生物学信息,为未来改变细胞的内部编程提供了理论依据。通过调控转录因子和DNA、组蛋白表观遗传学修饰和miRNA来抑制或促进不同基因的表达已成为可能。miRNA可在转录后水平调节靶基因的表达,即通过对mRNA特异序列的抑制,批量调节基因的活性而改变细胞的编程。
研究细胞重编程的方法
目前主要通过逆转录病毒 (主要是慢病毒)、腺病毒、质粒和转座子等介导的方式将转录因子对应的基因或者小分子导入成体细胞,将其进行重编程。
逆转录病毒转导 逆转录病毒又名反转录病毒,是一组RNA病毒,其病毒科下包括慢病毒在内共7属病毒。病毒感染宿主细胞时,在逆转录酶作用下,逆转录病毒首先将其RNA逆转录为DNA,然后将这段逆转录的基因插入细胞基因组中保持整合状态,并传给宿主细胞后代。慢病毒作为目前应用最广泛的逆转录病毒,其优点是转入基因可以长期稳定表达,并且对大部分哺乳动物细胞,包括神经元、干细胞等难转染的细胞,特别是体外悬浮生长的细胞,都有很好的转染效率。缺点是逆转录病毒整合到宿主细胞基因组的位置是随机的,这也就意味着有引起基因突变、激活癌基因的风险[1,13]。
腺病毒转导 腺病毒从腺样组织分离出来,其遗传物质为线型双股DNA,全长约30000~42000 bp。腺病毒的优点是几乎在所有已知细胞中都不整合到染色体中,因此不会干扰其他宿主基因,并且人类感染野生型腺病毒后仅产生轻微的自限性症状。腺病毒具有嗜上皮细胞性,因此对大多数细胞特别是上皮细胞有几乎100%的感染效率。腺病毒系统包装的病毒颗粒滴度高,浓缩后可以达到1013VP/ml,这一特点使其非常适用于基因治疗。腺病毒的缺点是由于其不能整合到宿主细胞基因组中,因此不能长期稳定表达[14-15]。
质粒转染转导 采用脂质体转染的方法,将外源性质粒转导进入目的细胞中表达。优点是细胞中不再留存有任何外源的DNA,不易使基因癌化。缺点是瞬时表达,转染成功细胞的获得率尚待提高[16]。
piggyBac转座子转导 piggyBac转座子是一个自主因子,遵循 “剪切—黏贴”机制,在生物体染色体中特征性的TTAA四核苷酸序列位点准确地切入和转座,并可以在作用一段时间后采用转座酶切除外源性插入序列。piggyBac转座子受生物体种类的限制较少,适用范围较广,转座频率较高。其作为非病毒体系提高了安全性,不易使基因癌化。缺点是需要多次使用转座酶去除转入序列,但仍然可能会留下一些痕迹[17-18]。
蛋白直接诱导 可以通过4个蛋白 (Oct4、Sox2、Klf4和c-Myc)诱导成体细胞重编程为iPSCs,优点是诱导过程不存在外源基因,缺点是诱导效率没有病毒载体诱导的效率高[19]。
RNA干扰 通过RNA干扰抑制某个或某几个基因的方法来对细胞基因表达进行重新编程,用十四烷基聚精氨酸肽链 [myristoylated polyarginine peptides,Myr-Ala-(Arg)7-Cys-CONH2,MPAP]将小干扰RNA导入细胞技术的出现[20],将有助于推动RNA干扰方法对成体细胞的重编程。
重编程细胞的来源及其转化方向的最新进展
重编程细胞产生iPSCs 不同胚层发育来源的成体细胞甚至胚外组织均有很多重编程产生iPSCs的报道 (表1),说明分化成熟的细胞都有可能通过重编程擦去原来的记忆去分化为胚胎干细胞。这些研究证实改变细胞基因表达程序 (时空和差异)就能改变细胞的分化方向。
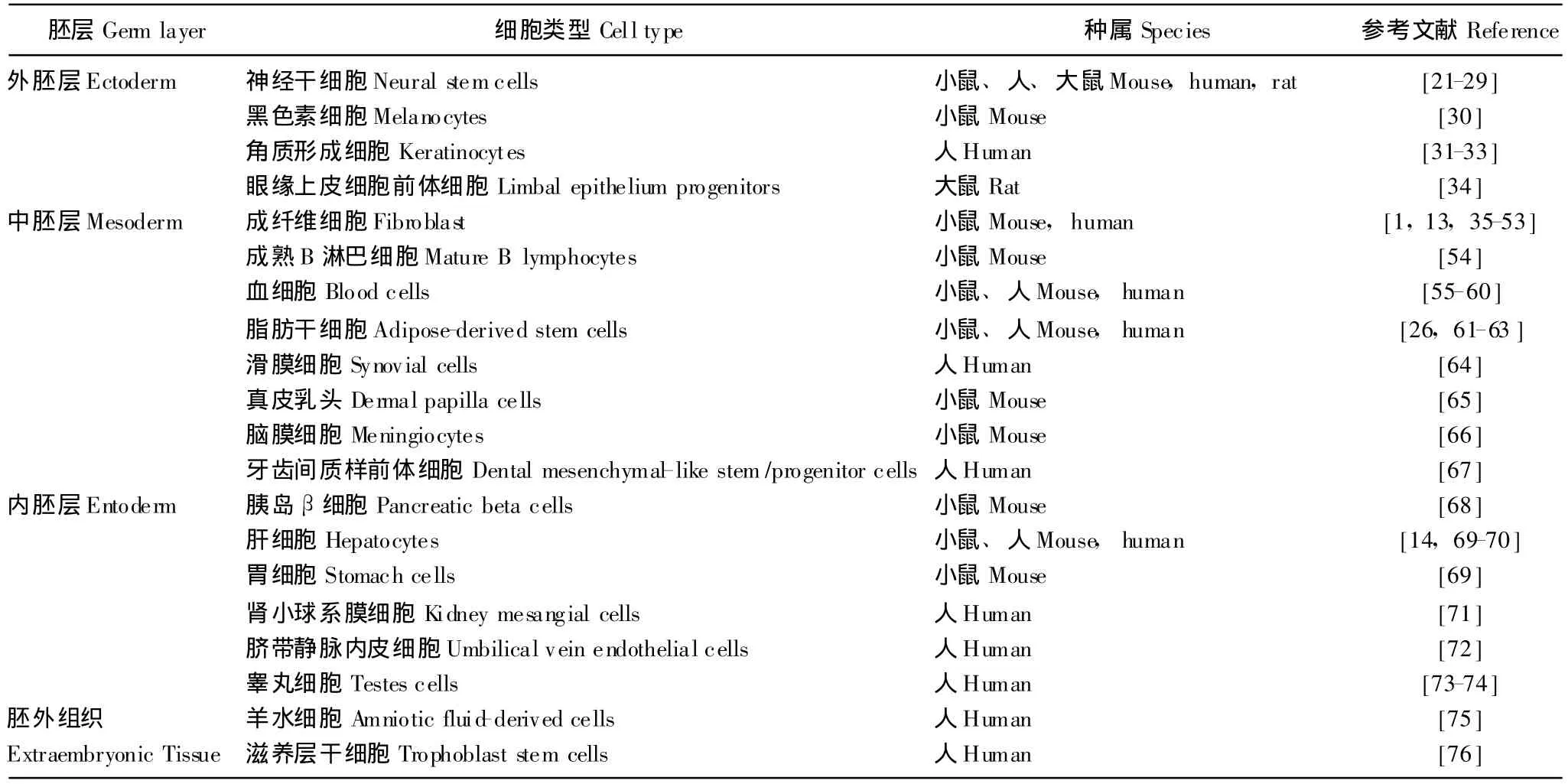
表1 各不同种属不同胚层发育来源的细胞诱导的诱导多潜能干细胞Table 1 Indceced pluripotent stem cells induced from cells of various species and germ layers
重编程后转分化的细胞 不但同一器官中发育于同一内胚层的胰腺外分泌腺泡细胞可以通过重编程向胰岛内分泌细胞 β细胞转化[77],不同器官中内胚层来源的肝细胞也可通过重编程向胰岛 β细胞转化[78]。而胰腺外分泌腺泡细胞还可通过重编程向肝细胞转化[79]。此外,内胚层来源的肝细胞可通过重编程向外胚层来源的神经细胞转化[80],中胚层来源的皮肤成纤维细胞可通过重编程向外胚层来源的神经细胞[81]及内胚层来源的肝样细胞转化[82],中胚层来源的骨髓间充质干细胞重编程向内胚层来源的胰岛β细胞转化,中胚层来源的成纤维细胞重编程向中胚层来源的心肌细胞转化[83],外胚层来源的表皮黑色素细胞重编程为外胚层来源神经嵴干细胞样细胞[84](表2)。综上,各不同胚层发育来源的成体细胞都有可能通过重编程擦去原来的记忆重新转分化为同一胚层或其他胚层发育来源的成体细胞,这同样证实改变细胞基因表达程序 (时空和差异)就能改变细胞的分化方向。
总之,目前正在进入一个可以通过调节关键基因对细胞进行重新编程的时代,人类未来将由此从许多分化的细胞获得更多所需要的另一些分化细胞,这对疾病的细胞替代治疗和细胞分化机制研究具有划时代的意义。
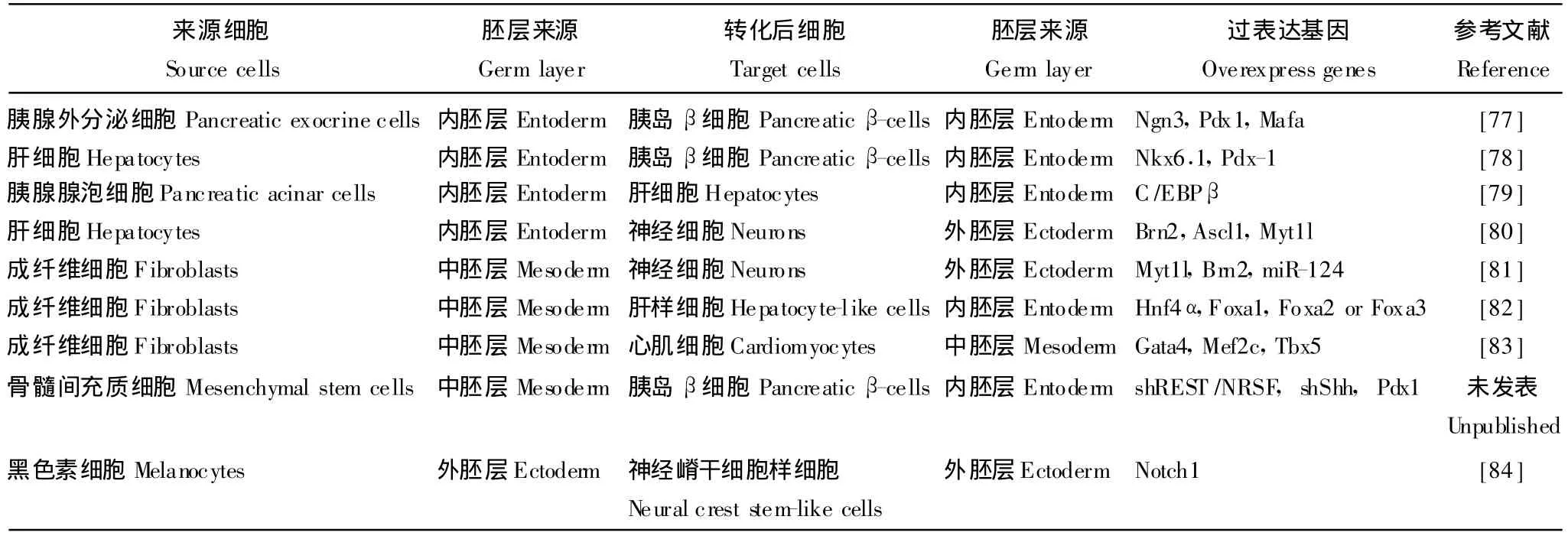
表2 不同胚层发育来源体细胞的重编程转分化Table 2 Reprogramming of adult cells from developmental original of various germ layers
[1]Takahashi K,Yamanaka S.Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors[J].Cell,2006,126(4):663-676.
[2]Yamanaka S.Strategies and new developments in the generation of patient-specific pluripotent stem cells[J].Cell Stem Cell,2007,1(1):39-49.
[3]Eguchi G,Kodama R.Transdifferentiation[J].Curr Opin Cell Biol,1993,5(6):1023-1028.
[4]Wu Ct,Morris JR.Genes,genetics and epigenetics:a correspondence[J].Science,2001,293(5532):1103-1105.
[5]Armstrong L,Lako M,Dean W,et al.Epigenetic modification is central to genome reprogramming in somatic cell nuclear transfer[J].Stem Cells,2006,24(4):805-814.
[6]Bartel DP.MicroRNAs:genomics,biogenesis,mechanism,and function[J].Cell,2004,116(2):281-297.
[7]Kanellopoulou C,Muljo SA,Kung AL,et al.Dicer deficient mouse embryonic stem cells are defective in differentiation and centromeric silencing[J].Genes Dev,2005,19(4):489-501.
[8]Trzaska KA,Reddy BY,Munoz JL,et al.Loss of RE-1 silencing factor in mesenchymal stem cell-derived dopamine progenitors induces functional maturity[J].Mol Cell Neurosci,2008,39(2):285-290.
[9]Johnson DS,Mortazavi A,Myers RM,et al.Genome-wide mapping of in vivo protein-DNA interactions[J].Science,2007,316(5830):1497-1502.
[10]Mortazavi A,Leeper Thompson EC,Garcia ST,et al.Comparative genomics modeling of the NRSF/REST repressor network:from single conserved sites to genome-wide repertoire[J].Genome Res,2006,16(10):1208-1221.
[11]Kemp DM,Lin JC,Habener JF.Regulation of Pax4 paired homeodomain gene by neuron-restrictive silencer factor[J].J Biol Chem,2003,278(37):35057-35062.
[12]Schwitzgebel VM,Mamin A,Brun T,et al.Agenesis ofhu-man pancreas due to decreased half-life of insulin promoter factor 1[J].J Clin Endocrinol Metab,2003,88(9):4398-4406.
[13]Takahashi K,Tanabe K,Ohnuki M,et al.Induction ofpluripotent stem cells from adult human fibroblasts by defined factors[J].Cell,2007,131(5):861-872.
[14]Stadtfeld M,Nagaya M,Utikal J,etal.Induced pluripotent stem cells generated without viral integration[J].Science,2008,322(5903):945-949.
[15]Okita K,Nakagawa M,Hyenjong H,et al.Generation of mouse induced pluripotent stem cells without viral vectors[J].Science,2008,322(5903):949-953.
[16]Yu J,Hu K,Smuga-Otto K,etal.Human induced pluripotent stem cells free of vector and transgene sequences[J].Science,2009,324(5928):797-801.
[17]Kaji K,Norrby K,Paca A,et al.Virus-free induction of pluripotency and subsequent excision of reprogramming factors[J].Nature,2009,458(7239):771-775.
[18]Woltjen K,Michael IP,Mohseni P,et al.piggyBac transposition reprograms fibroblasts to induced pluripotent stem cells[J].Nature,2009,458(7239):766-770.
[19]Zhou H,Wu S,Joo JY,et al.Generation of induced pluripotent stem cells using recombinant proteins[J].Cell Stem Cell,2009,4(5):381-384.
[20]Medarova Z,Pham W,Farrar C,et al.In vivo imaging of siRNA delivery and silencing in tumors[J].Nat Med,2007,13(3):372-377.
[21]Shi Y,Do JT,Desponts C,et al.A combined chemical and genetic approach for the generation of induced pluripotent stem cells[J].Cell Stem Cell,2008,2(6):525-528.
[22]Kim JB,Zaehres H,Wu G,et al.Pluripotent stem cells induced from adult neural stem cells by reprogramming with two factors[J].Nature,2008,454(7204):646-650.
[23]Duinsbergen D,Eriksson M,'t Hoen PA,et al.Induced pluripotency with endogenous and inducible genes[J].Exp Cell Res,2008,314(17):3255-3263.
[24]Kim JB,Sebastiano V,Wu G,et al.Oct4-induced pluripotency in adult neural stem cells[J].Cell,2009,136(3):411-419.
[25]EminliS,Utikal J,Arnold K,etal.Reprogramming of neural progenitor cells into induced pluripotent stem cells in the absence of exogenous Sox2 expression[J].Stem Cells,2008,26(10):2467-2474.
[26]Tat PA,Sumer H,Jones KL,etal.The efficientgeneration of induced pluripotent stem (iPS)cells from adult mouse adipose tissue-derived and neural stem cells [J].Cell Transplant,2010,19(5):525-536.
[27]Kim JB,Greber B,Araúzo-Bravo MJ,et al.Direct reprogramming of human neural stem cells by OCT4[J].Nature,2009,461(7264):643-649.
[28]Chang MY,Kim D,Kim CH,et al.Direct reprogramming of rat neural precursor cells and fibroblasts into pluripotent stem cells[J].PLoS One,2010,5(3):e9838.
[29]Do JT,Joo JY,Han DW,et al.Generation of parthenogenetic induced pluripotent stem cells from parthenogenetic neural stem cells[J].Stem Cells,2009,27(12):2962-2968.
[30]Utikal J,Maherali N,Kulalert W,et al.Sox2 is dispensable for the reprogramming of melanocytes and melanoma cells into induced pluripotent stem cells[J].J Cell Sci,2009,122(Pt19):3502-3510.
[31]Aasen T,Raya A,Barrero MJ,et al.Efficient and rapid generation of induced pluripotent stem cells from human keratinocytes[J].Nat Biotechnol,2008,26(11):1276-1284.
[32]Maherali N,Ahfeldt T,Rigamonti A,et al.A high-efficiency system for the generation and study of human induced pluripotent stem cells[J].Cell Stem Cell,2008,3(3):340-345.
[33]Linta L,Stockmann M,Kleinhans KN,et al.Rat embryonic fibroblasts improve reprogramming of human keratinocytes into induced pluripotent stem cells[J].Stem Cells Dev,[2011-08-05].http://www.liebertonline.com/doi/abs/10.1089/scd.2011.0026.
[34]Balasubramanian S,Babai N,Chaudhuri A,et al.Non cell-autonomous reprogramming of adult ocular progenitors:generation of pluripotent stem cells without exogenous transcription factors[J].Stem Cells,2009,27(12):3053-3062.
[35]Nakagawa M,Koyanagi M,Tanabe K,et al.Generation of induced pluripotent stem cells without Myc from mouse and human fibroblasts[J].Nat Biotechnol,2008,26(1):101-106.
[36]Wernig M,Meissner A,Cassady JP,et al.c-Myc is dispensable for direct reprogramming of mouse fibroblasts[J].Cell Stem Cell,2008,2(1):10-12.
[37]Wernig M,Lengner CJ,Hanna J,et al.A drug-inducible transgenic system for direct reprogramming of multiple somatic cell types[J].Nat Biotechnol,2008,26(8):916-924.
[38]Wernig M,Meissner A,Foreman R,et al.In vitro reprogramming of fibroblasts into a pluripotent ES-cell-like state[J].Nature,2007,448(7151):318-324.
[39]Meissner A,Wernig M,Jaenisch R.Direct reprogramming of genetically unmodified fibroblasts into pluripotent stem cells[J].Nat Biotechnol,2007,25(10):1177-1181.
[40]Okita K,Ichisaka T,Yamanaka S.Generation of germlinecompete induced pluripotent stem cells[J].Nature,2007,448(7151):313-317.
[41]Feng B,Jiang J,Kraus P,et al.Reprogramming of fibroblasts into induced pluripotent stem cells with orphan nuclear receptor Esrrb[J]Nat Cell Biol,2009,11(2):197-203.
[42]Heng JC,Feng B,Han J,et al.The nuclear receptor Nr5a2 can replace Oct4 in the reprogramming of murine somatic cells to pluripotentcells[J].Cell Stem Cell,2010,6(2):167-174.
[43]Yu J,Vodyanik MA,Smuga-Otto K,et al.Induced pluripotent stem cell lines derived from human somatic cells[J].Science,2007,318(5858):1917-1920.
[44]Park IH,Zhao R,West JA,et al.Reprogramming of human somatic cells to pluripotency with defined factors[J].Nature,2008,451(7175):141-146.
[45]Dimos JT,Rodolfa KT,Niakan KK,et al.Induced pluripotent stem cells generated from patients with ALS can be differentiated into motor neurons[J].Science,2008,321(5893):1218-1221.
[46]Hockemeyer D,Soldner F,Cook EG,et al.A drug-inducible system for direct reprogramming of human somatic cells to pluripotency[J].Cell Stem Cell,2008,3(3):346-353.
[47]MaheraliN,Ahfeldt T,Rigamonti A,et al.A high-efficiency system for the generation and study of human induced pluripotent stem cells[J].Cell Stem Cell,2008,3(3):340-345.
[48]Lowry WE,Richter L,Yachechko R,et al.Generation of human induced pluripotent stem cells from dermal fibroblasts[J].Proc Natl Acad Sci USA,2008,105(8):2883-2888.
[49]Liao J,Wu Z,Wang Y,et al.Enhanced efficiency of generating induced pluripotent stem(iPS)cells from human somatic cells by a combination of six transcription factors[J].Cell Res,2008,18(5):600-603.
[50]Mali P,Ye Z,Hommond HH,et al.Improved efficiency and pace of generating induced pluripotent stem cells from human adult and fetal fibroblasts[J].Stem Cells,2008,26(8):1998-2005.
[51]Park IH,Arora N,Huo H,et al.Disease-specific induced pluripotent stem cells[J].Cell,2008,134(5):877-886.
[52]Eggenschwiler R,Loya K,Sgodda M,et al.Hepatic differentiation of murine disease-specific induced pluripotent stem cells allows disease modelling in vitro[J].Stem Cells Int,2011:924782[2011-09-29].http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3184399/?tool=pubmed.
[53]Araki R,Jincho Y,Hoki Y,et al.Conversion of ancestral fibroblasts to induced pluripotent stem cells[J].Stem Cells,2010,28(2):213-220.
[54]Hanna J,Markoulaki S,Schorderet P,et al.Direct reprogramming of terminally differentiated mature B lymphocytes to pluripotency[J].Cell,2008,133(2):250-264.
[55]Eminli S,Foudi A,Stadtfeld M,et al.Differentiation stage determines potential ofhematopoietic cells for reprogramming into induced pluripotent stem cells[J].Nat Genet,2009,41(9):968-976.
[56]Loh YH,Agarwal S,Park IH,et al.Generation of induced pluripotent stem cells from human blood[J].Blood,2009,113(22):5476-5479.
[57]Haase A,Olmer R,Schwanke K,et al.Generation of induced pluripotent stem cells from human cord blood[J].Cell Stem Cell,2009,5(4):434-441.
[58]Giorgetti A,Montserrat N,Aasen T,et al.Generation of induced pluripotent stem cells from human cord blood using OCT4 and SOX2[J].Cell Stem Cell,2009,5(4):353-357.
[59]Takenaka C,Nishishita N,Takada N,et al.Effective generation of iPS cells from CD34+cord blood cells by inhibition of p53[J].Exp Hematol,2010,38(2):154-162.
[60]Kunisato A,Wakatsuki M,Kodama Y,et al.Generation of induced pluripotent stem cells by efficient reprogramming of adult bone marrow cells[J].Stem Cells Dev,2010,19(2):229-238.
[61]Sugii S,Kida Y,Kawamura T,et al.Human and mouse adipose-derived cells support feeder-independent induction of pluripotent stem cells[J].Proc Natl Acad Sci USA,2010,107(8):3558-3563.
[62]Aoki T,Ohnishi H,Oda Y,et al.Generation of induced pluripotent stem cells from human adipose-derived stem cells without c-MYC[J].Tissue Eng Part A,2010,16(7):2197-2206.
[63]Sun N,Panetta NJ,Gupta DM,et al.Feeder-free derivation of induced pluripotent stem cells from adult human adipose stem cells[J].Proc Natl Acad Sci USA,2009,106(37):15720-15725.
[64]Kim MJ,Son MJ,Son MY,et al.Generation of human induced pluripotent stem cells from osteoarthritis patient-derived synovial cells[J].Arthritis Rheum,2011,63(10):3010-3021.
[65]Tsai SY,Clavel C,Kim S,et al.Oct4 and klf4 reprogram dermal papilla cells into induced pluripotent stem cells[J].Stem Cells,2010,28(2):221-228.
[66]Qin D,Gan Y,Shao K,et al.Mouse meningiocytes express Sox2 and yield high efficiency of chimeras after nuclear reprogramming with exogenous factors[J].J Biol Chem,2008,283(48):33730-33735.
[67]Yan X,Qin H,Qu C,et al.iPS cells reprogrammed from human mesenchymal-like stem/progenitor cells of dental tissue origin[J].Stem Cells Dev,2010,19(4):469-480.
[68]Stadtfeld M,Brennand K,Hochedlinger K.Reprogramming of pancreatic beta cells into induced pluripotent stem cells[J].Curr Biol,2008,18(12):890-894.
[69]Aoi T,Yae K,Nakagawa M,et al.Generation of pluripotent stem cells from adult mouse liver and stomach cells[J].Science,2008,321(5889):699-702.
[70]Liu H,Ye Z,Kim Y,et al.Generation of endoderm-derived human induced pluripotent stem cells from primary hepatocytes[J].Hepatology,2010,51(5):1810-1819.
[71]Song B,Niclis JC,Alikhan MA,et al.Generation of induced pluripotent stem cells from human kidney mesangial cells[J].J Am Soc Nephrol,2011,22(7):1213-1220.
[72]Lagarkova MA,Shutova MV,Bogomazova AN,et al.Induction of pluripotency in human endothelial cells resets epigenetic profile on genome scale[J].Cell Cycle,2010,9(5):937-946.
[73]Golestaneh N,Kokkinaki M,Pant D,et al.Pluripotent stem cells derived from adult human testes[J].Stem Cells Dev,2009,18(8):1115-1126.
[74]Conrad S,Renninger M,Hennenlotter J,et al.Generation of pluripotent stem cells from adult human testis[J].Nature,2008,456(7220):344-349.
[75]Li C,Zhou J,Shi G,et al.Pluripotency can be rapidly and efficiently induced in human amniotic fluid-derived cells[J].Hum Mol Genet,2009,18(22):4340-4349.
[76]Wu T,Wang H,He J,et al.Reprogramming of trophoblast stem cells into pluripotent stem cells by Oct4[J].Stem Cells,2011,29(5):755-763.
[77]Zhou Q,Brown J,Kanarek A,et al.In vivo reprogramming of adult pancreatic exocrine cells to beta-cells[J].Nature,2008,455(7213):627-632.
[78]Gefen-Halevi S,Rachmut IH,Molakandov K,et al.NKX6.1 promotes PDX-1-induced liver to pancreatic β-cells reprogramming[J].Cell Reprogram,2010,12(6):655-664.
[79]Al-Adsani A,Burke ZD,Eberhard D,et al.Dexamethasone treatment induces the reprogramming of pancreatic acinar cells to hepatocytes and ductal cells[J].PLoS One,2010,5(10):e13650.
[80]Marro S,Pang ZP,Yang N,etal.Direct lineage conversion of terminally differentiated hepatocytes to functional neurons[J].Cell Stem Cell,2011,9(4):374-382.
[81]Ambasudhan R,Talantova M,Coleman R,et al.Direct reprogramming of adult human fibroblasts to functional neurons under defined conditions[J].Cell Stem Cell,2011,9(2):113-118.
[82]Sekiya S,Suzuki A.Direct conversion of mouse fibroblasts to hepatocyte-like cells by defined factors[J].Nature,2011,475(7356):390-393.
[83]Yang L.From fibroblast cells to cardiomyocytes:direct lineage reprogramming[J].Stem Cell Res Ther,2011,2(1):1.
[84]Zabierowski SE,Baubet V,Himes B,et al.Direct reprogramming of melanocytes to neural crest stem-like cells by one defined factor[J].Stem Cells,2011,29(11):1752-1762.
