光学成像模糊对微扫视性眼球运动影响的研究*
2011-07-16史学锋许丽敏赵堪兴
史学锋 王 婷 许丽敏 赵堪兴
既往关于异常视觉经验对视觉感知系统的响应特性及其功能发育的影响的研究较为广泛,很少关注其对眼球运动系统的影响。然而,视觉感知系统与眼球运动系统是相辅相成、相互作用的[1]。近年来,新的无创性眼球运动追踪技术的出现使得人们对一种特殊的眼球运动形式—微扫视(microsac⁃cade)产生研究兴趣[2]。本研究应用高速眼球运动记录系统对正常屈光成像条件和光学成像模糊条件下人眼微扫视性眼球运动的行为学差异进行比较,以期探讨异常视觉经验对微扫视性眼球运动的影响。
1 对象与方法
1.1 研究对象 2011年1月—3月在天津市眼科医院就诊的屈光矫正正常或无屈光不正的受试者13例,男6例,女7例,年龄7~11岁,平均8.2岁。所有受试者均排除可能存在的眼部疾病,如青光眼、白内障、视网膜疾病、视神经与视路疾病等。所有受试者或其监护人均签署本院伦理委员会批准的知情同意书,理解此项无创性实验研究的目的和意义。根据眼别及测试条件分成4组:正常屈光成像状态下主导眼为Nd组,非主导眼为Nn组,增戴-3.00 D镜片条件下主导眼为Bd组,非主导眼为Bn组。
1.2 研究方法
1.2.1 眼球运动记录 所有受试者均在一间严格安静、光线均匀、亮度较暗的测试室内接受实验检查,室内避免视觉干扰对象的出现。受试者能够舒适地坐于设备前,将其头部尽可能保持不动地放置在一个下颌托上。遇不能配合的受试者,则放弃此项测试。受试者双眼离正前方投影显示视标的距离为2.0 m。采用Eyelink-2000高速眼球运动记录系统(加拿大SR Research公司)进行眼球运动波形的记录,其时间分辨率为0.5 ms或1 ms,空间分辨率为0.01°。数据的采集通过EB软件(加拿大SR Research公司)建立的程序来实现。记录完成后得到的EDF格式数据经转换成ASCII格式文件后导入Matlab(美国MathWorks公司)进行离线分析处理。
1.2.2 视觉刺激任务 令受试者注视眼前屏幕中央一个直径为0.15°视角的亮点。所有测试均在单眼注视条件下进行,即测试时挡住未被测试眼。受试者能够注视目标时令其自行按下启动键开始每一次测试。每次测试前均需进行记录系统的校准与验证,通过者方能开始记录。记录持续6 s,此过程中受试者应能稳定保持注视。如出现任何的注视转移,此次测试记录将被舍弃。每只受试眼必须接受6次以上测试方能结束任务。所有受试者每只眼分别在两种成像状态下接受上述测试:(1)正常屈光成像状态:无屈光不正者裸眼测试,屈光不正者戴最佳矫正眼镜测试。(2)成像模糊状态:在正常屈光状态基础上加戴-3.00D镜片进行测试。
1.2.3 微扫视甄别与分析 采用Matlab编写的一个微扫视自动甄别程序进行微扫视事件的判别和分析,该程序的核心逻辑参照文献[3]并进行了适当调整以达到最佳甄别效果,即同时满足以下条件:(1)眼球运动的角速度超过速度概率分布的6个标准差。(2)事件的时程超过12 ms。(3)相邻事件间隔时间超过20 ms以防止将可能出现的扫视后超射当做一次新的微扫视。如一次测试中存在超过3°视角的扫视或眼球运动记录过程中因眨眼等因素导致部分数据丢失,则舍弃该次测试的数据。经过以上数据甄别与筛选后,再对该测试眼的微扫视数据集进行主序列分析,见图1。以进一步保证所甄别的微扫视事件的正确性。程序分析获得的基本数据包括微扫视幅度、峰值速度、发生频率及微扫视间隔时间等。
1.3 统计学处理 采用Origin 8.0统计软件及Matlab 2008统计学工具包对研究数据进行分析。所有计量资料进行正态性检验及方差齐性检验,多组间比较采用单因素方差分析和Turkey检验,如不满足正态分布或方差不齐采用Krus⁃kal-Wallis法以及Nemenyi法检验。以P<0.05为差异有统计学意义。
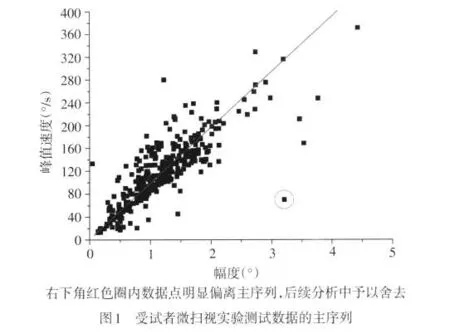
2 结果
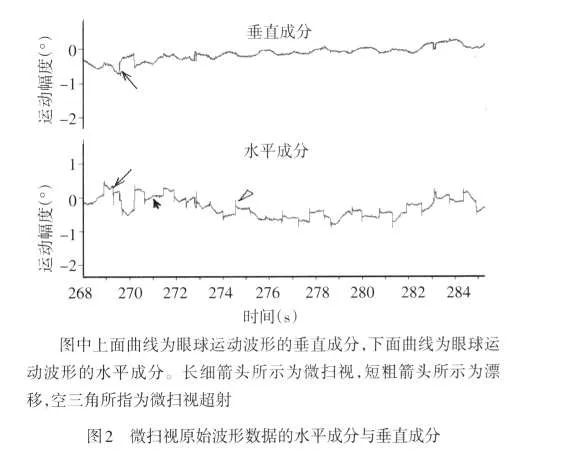
2.1 微扫视性眼球运动的基本特征 微扫视性眼球运动为注视性眼球运动中幅度最大、速度最快的类型,偶见微扫视终止后的超射现象,见图2。
2.2 微扫视幅度与峰值速度 各组间平均微扫视幅度差异无统计学意义(P>0.05),Bd组、Bn组峰值速度较Nd组、Nn组显著性降低(P<0.01),Bd组与Bn组、Nd组与Nn组峰值速度比较差异无统计学意义(P>0.05),见表1。
2.3 微扫视发生频率 Bd组、Bn组微扫视发生频率较Nd组、Nn组明显下降(P<0.01),Bd组与Bn组、Nd组与Nn组微扫视发生频率比较差异无统计学意义(P>0.05),见表1。
2.4 微扫视间隔时间 各组微扫视性眼球运动间隔时间比较,Bd组、Bn组较Nd组、Nn组明显延长(P<0.01),Bd组与Bn组、Nd组与Nn组微扫视间隔时间比较差异无统计学意义(P>0.05),见表1。
表1 各组程序分析获得基本数据的比较 ( ±s)
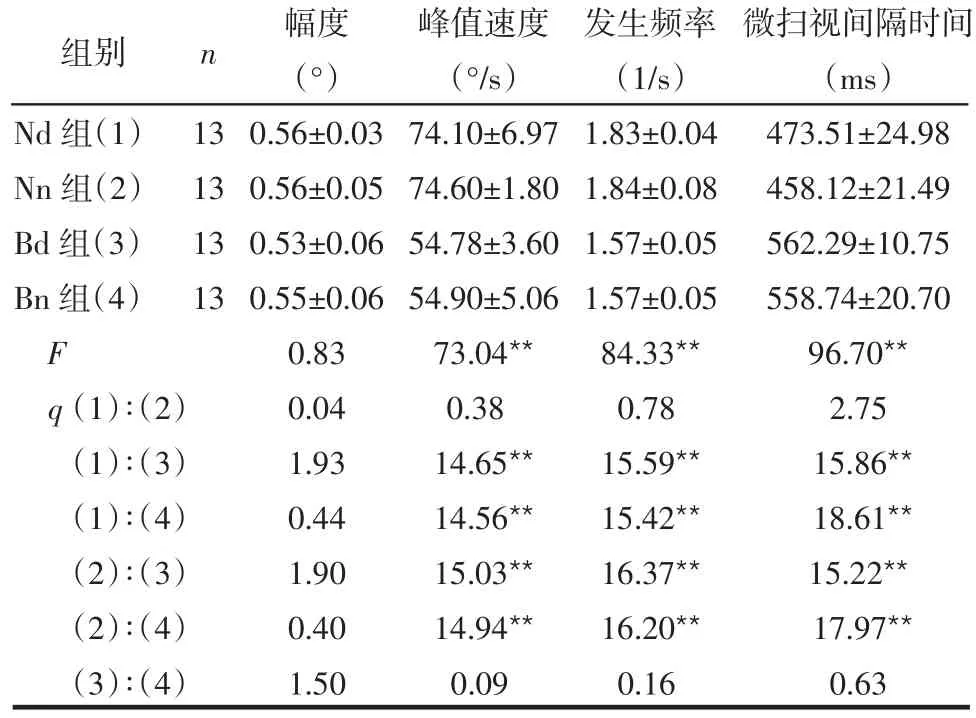
表1 各组程序分析获得基本数据的比较 ( ±s)
**P<0.01
Nd组(1)Nn组(2)Bd组(3)Bn组(4)F q(1)∶(2)(1)∶(3)(1)∶(4)(2)∶(3)(2)∶(4)(3)∶(4)13 13 13 13幅度(°)0.56±0.03 0.56±0.05 0.53±0.06 0.55±0.06 0.83 0.04 1.93 0.44 1.90 0.40 1.50峰值速度(°/s)74.10±6.97 74.60±1.80 54.78±3.60 54.90±5.06 73.04**0.38 14.65**14.56**15.03**14.94**0.09发生频率(1/s)1.83±0.04 1.84±0.08 1.57±0.05 1.57±0.05 84.33**0.78 15.59**15.42**16.37**16.20**0.16微扫视间隔时间(ms)473.51±24.98 458.12±21.49 562.29±10.75 558.74±20.70 96.70**2.75 15.86**18.61**15.22**17.97**0.63组别 n
3 讨论
人眼对外界事物的感知首先依赖于注视,但注视并非是静止的,而是始终保持着微小的肉眼难以分辨的运动状态。注视性眼球运动包括微扫视、漂移与颤动,其中微扫视是注视过程中幅度最大、速度最快的一种眼球运动形式[4]。对该种眼球运动的研究依赖于精细化的眼球运动测量手段以及对细微眼球运动变化的自动甄别技术[2,5]。近10年科学技术发展使得以上研究瓶颈获得解决[6]。本研究采用了目前世界上最高采样速率(可高达2 000 Hz)的眼球运动采集系统对正常屈光成像和光学成像模糊条件下人眼微扫视性眼球运动的行为学差异进行了比较,发现成像模糊导致微扫视的各项特征参数发生改变。
既往研究表明,微扫视幅度和峰值速度呈直线正相关关系,此为主序列分析的基础。Keith等[7]研究提出微扫视幅度控制信号来源于上丘和额视区,其信号具有视网膜拓扑分布特征,而微扫视速度控制信号来源于网状核神经元。网状核视觉区域的神经元也具有精确的视网膜拓扑代表特征。不同来源的控制信号可能最终通过动眼神经核团实现信号整合,并以脉冲宽度的形式决定微扫视幅度和峰值速度。本研究发现正常屈光成像状态和光学成像模糊状态下微扫视平均幅度无明显差异,而光学成像模糊引起微扫视峰值速度的降低,提示光学成像模糊可能只影响眼球运动速度控制信号中枢神经元的活动,而对上丘及额视区可能不造成影响。
本研究发现,和正常成像条件相比成像模糊造成微扫视发生频率降低。这和既往研究在弱视眼上发现微扫视抑制现象类似,表明视觉感知对比度的降低可能是造成微扫视发生频率降低的原因,而视皮层功能及结构性异常并非必要条件。神经心理学研究表明,微扫视具有拮抗持续注视造成的视知觉消退的作用,反之,视知觉的消退易诱发更多的微扫视的发生,但也有研究不支持这一观点[8]。本研究认为,视觉质量降低与视知觉消退现象不同,后者可能反馈性促进视觉注意的水平,而前者则可能降低视觉注意的水平,进而调节微扫视发生频率[8]。此外,本研究还发现和正常成像条件相比,成像模糊造成微扫视间隔时间的延长,这也是造成后者微扫视发生频率降低的原因之一。
综上,异常屈光成像状态可影响人眼微扫视性眼球运动的行为,表现为发生频率降低、微扫视间隔时间延长及峰值速度减低。高速眼球运动记录可作为评价视觉质量、研究视觉感知与眼球运动行为之间的关系的重要工具。
[1]CollewijnH,KowlerE.The significance of microsaccades for vision and oculomotor control[J].J Vis,2008,8(14):20.1-21.
[2]Rolfs M.Microsaccades:small steps on a long way[J].Vision Res,2009,49(20):2415-2441.
[3]Engbert R,Mergenthaler K.Microsaccades are triggered by low retinalim⁃ageslip[J].Proc Natl Acad Sci USA,2006,103(18):7192-7197.
[4]Rolfs M,Kliegl R,Engbert R.Toward a model of microsaccade genera⁃tion:the case of microsaccadic inhibition[J].JVis,2008,8(11):5.1-23.
[5]Otero-Millan J,Troncoso XG,Macknik SL,et al.Saccades and micro⁃saccades during visual fixation,exploration,and search:foundations for a common saccadic generator[J].JVis,2008,8(14):21.1-18.
[6]Troncoso XG,Macknik SL,Martinez-CondeS.Microsaccades counter⁃act perceptual filling-in[J].JVis,2008,8(14):15.1-9.
[7]Keith GP,Blohm G,Crawford JD.Influence of saccade efference copy on the spatiotemporal properties of remapping:a neural network study[J].J Neurophysiol,2010,103(1):117-139.
[8]Poletti M,Rucci M.Eye movements under various conditions of image fading[J].J Vis,2010,10(3):6.1-18.
