青海高寒半干旱区沙木蓼水势研究
2011-06-07李安超贺康宁郭倩倩孙晓涵曹广月刘帅华
李安超,贺康宁,郭倩倩,孙晓涵,王 辉,曹广月,刘帅华
(北京林业大学水土保持学院,水土保持与荒漠化防治教育部重点实验室,北京 100083)
沙木蓼(Atraphaxis mandshurica),为蓼科灌木,直立生长,株高2 m~3 m,冠幅3 m×3 m;老枝皮褐色,纵裂;1年生枝伸长,节间2.5 cm~4 cm;叶圆形,鲜绿色;花少色,总状花序,淡玫瑰色,生于一年生枝上部苞腋;瘦果,卵状三角形,深褐色,有光泽。沙木蓼主要分布于库不齐沙漠、乌兰布和沙漠、毛乌素沙地、宁夏河东沙地、腾格里沙漠、巴丹吉林沙漠、河西走廊等地,生境主要为流动沙丘和丘间低地。沙木蓼根系发达,吸水力强,适应沙生环境、不怕风蚀、耐沙埋、耐旱、耐寒,造林成活率高,生长迅速,是荒漠与半荒漠地区优良的固沙植[1-5]。
植物的叶水势反映了植物的水分状况和植物从土壤中吸收水分的能力,是土壤—植物—大气连续体(SPAC)中水分运转驱动力的重要组成部分和重要环节[6],是衡量植物抗旱性的一个重要生理指标[7]。目前,国内主要研究沙木蓼的引种与栽培技术,对其水势方面的研究还鲜有报道。笔者对沙木蓼叶水势的动态变化及其与蒸腾速率、气象因子、土壤含水量的关系等进行了研究分析,以期为西部干旱半干旱区植被恢复与重建提供理论依据和指导。
1 材料与方法
1.1 试验地概况
试验地点位于青海省大通县东峡林场,地理坐标36°56′27.33″N,101°41′49.48″E,海拔2 475 m。该地区属温带高寒大陆性气候,年平均气温2.8℃,极端最高气温30.9℃,极端最低气温-33.1℃,全年≥5℃的积温平均为1 908℃,无霜期一般70~120 d。多年平均降雨量508 mm,年内分配不均,全年雨水主要集中在7~8月。年平均蒸发总量1 290 mm。土壤类型为黄土母质上发育的山地棕褐土和栗钙土。田间最大持水量30.2%,表层土壤平均容重1.06 g/cm3。
1.2 试验材料
选取由青海省林科院和青海省大通县林业局实验苗圃提供的生长健壮的3年生沙木蓼,在生长季5~8月份对沙木蓼的根、茎、叶水势进行了测定,同时对沙木蓼灌木林地土壤水分及其主要气候因子进行了观测。
1.3 试验方法
1.3.1 植物叶水势的测定 采用美国产的PSYPRO露点水势仪,在2008年5~8月每月选择1~2个典型晴天为测定日,从7∶00~19∶00,每隔2 h,选择树冠中上部向阳面生发育良好的叶片,用打孔器取叶,放入C-52样品室,每5 min记录一次数据,取平均值作为该时刻植物的叶水势(LWP)。
1.3.2 蒸腾速率的测定 采用美国生产的Li-1600稳态气孔计,选用与测量叶水势相同的叶片进行同步观测,每个时刻重复3次求其平均值作为该时刻的蒸腾速率(Tr)。
1.3.3 气象因子的测定 采用美国生产的Vantage pro 2自动气象站,每10 min记录一次气温(T)、2 m处相对湿度(RH)、光量子通量密度(PAR)、降雨量等气象因子,连续观测记录整个植物生长季。
1.3.4 土壤含水量 2008年6月份开始在温室大棚内进行盆栽水分胁迫试验。每隔两天清晨(6∶00左右)测定苗木叶水势,同时用美国生产的TDR水分速测仪测定盆栽土壤体积含水量(SWC),每次测定3个重复。土壤水分速测仪在测定前用烘干法进行标定。
1.4 数据处理
所获得的数据用EXCEL和SPSS统计软件进行分析处理。
2 结果与分析
2.1 沙木蓼叶水势的日、月变化
2.1.1 沙木蓼水势的日变化规律 图1为沙木蓼叶水势的日变化图,由图中可看出各月叶水势最高值出现在清晨(均值为-2.01 MPa)。日出后,随着光照强度增强,气温升高,植物蒸腾作用、光合能力逐渐增强,当根向叶片供水能力小于植物体各项生理活动的耗水速率时,沙木蓼体内水分出现亏缺,叶水势将逐渐降低,从13∶00~15∶00叶水势下降至最低值(均值-3.40 MPa),15∶00以后至夜间随着光照强度减弱,随着气温降低,植物蒸腾作用减弱,叶水势开始回升,直到19∶00又达到较高水平。但此时沙木蓼的叶水势通常低于清晨叶水势。此外,由于中午光照强度大,气温高,空气湿度低,植物为了保持体内水分,防止因蒸腾而流失,叶片气孔会减小或关闭,所以此时叶水势会有所升高。沙木蓼叶水势的日变化规律表明,植物水势的日变化和环境因子的日变化关系密切[8-9]。
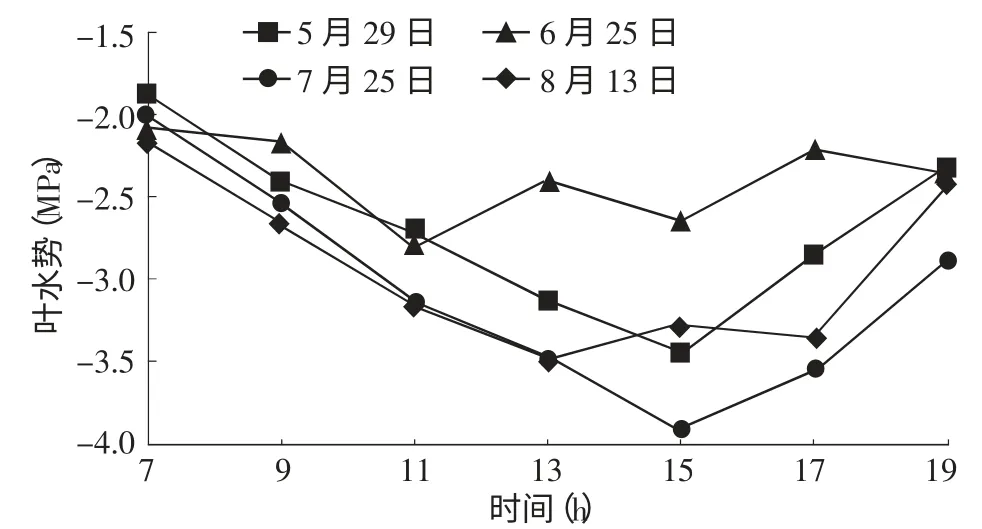
图1 沙木蓼叶水势的日变化
2.1.2 沙木蓼叶水势的月变化 早晨的叶水势是在植物经过一夜的吸水恢复后达到的较稳定值,比其他时间的叶水势更能反映叶水分的亏缺情况。因此,在进行沙木蓼叶水势月变化分析时,选择各月清晨(7∶00)的叶水势为标准进行比较,并利用各月叶水势的均值和中午(13∶00~15∶00)值进行辅助分析。如图1和表1所示,虽然不同时期的叶水势值大小存在差异,但无论是清晨、中午的叶水势还是其月均值在不同时期的变化规律基本一致。5月、7月的清晨叶水势较高,分别为-1.85,-1.98 MPa,6月较低,为-2.05 MPa,8月最低,为-2.14 MPa。这是由于5月是植物的萌芽返青期,对水分的需求相对较少,土壤水分还有盈余,此时光照强度较弱,植物的蒸腾作用也较弱,因此植物叶水势较高。6月植物进入生长旺盛期,气温升高,光照强度增大,植物体为了获得水分满足正常的生理活动,蒸腾强度显著增加,虽然降雨量的增加使土壤水分略有恢复,但还不能满足叶片蒸腾的需要,叶片处于缺水状态,因此叶水势也保持在较低的水平。7月,随着雨季的到来,大气相对湿度、土壤含水量有所升高,植物水分欠缺状况得到了缓解,致使叶水势有所升高。8月,植物开始进入生长末期,生长缓慢,虽然降雨使土壤水分得到了一些补充,但当地在这一时期光照依然很强,气温较高,根系供水相对于蒸腾耗水速率来说显得有些不足,故叶水势较低。从结果上看,沙木蓼生长最旺盛的6~7月是对水分要求较高的时期,这一时期的水分条件对沙木蓼的生长起到了关键作用。
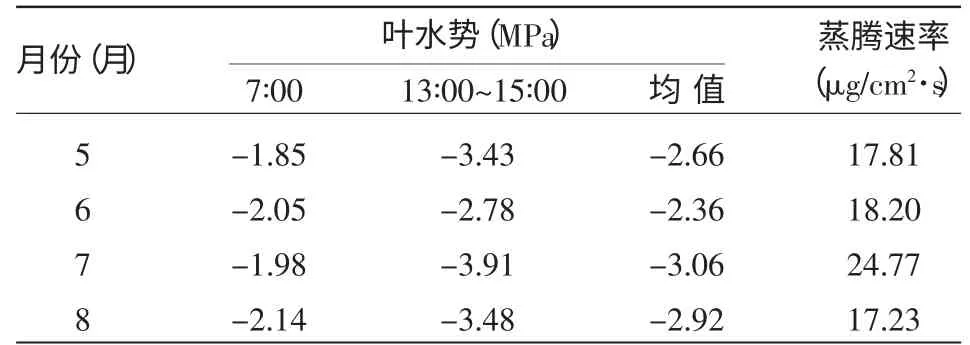
表1 不同月份沙木蓼叶水势日均值变化
2.2 沙木蓼叶水势和蒸腾速率的关系
从表1看出,沙木蓼叶水势和蒸腾速率的月变化趋势基本相反。从6到7月,蒸腾速率从18.2 μg/cm2·s升高到24.77μg/cm2·s,对应的叶水势从叶水势-2.36 MPa降至-3.06 MPa,8月的蒸腾速率降至17.23μg/cm2·s,而叶水势升高到-2.92 MPa。从图1、图2可以看出,沙木蓼叶水势和蒸腾速率的日过程曲线有一定的相关性。在一天内土壤水分充足且处于相对稳定的状态下,清晨随着光照增强,植物的气孔导度增大,叶内外的水汽压差增大,蒸腾速率不断提高,叶水势随之降低,直至13∶00~15∶00蒸腾速率达到峰值,叶水势也基本降至最低,其后叶水势随蒸腾速率的降低而升高,直到进入夜间后,当蒸腾耗水速率低于根系的吸水速率时,叶片的细胞才得以补充水分。其中7,8月份的蒸腾速率日变化呈双峰曲线,这是因为在光照最强时,植物气孔会关闭一段时间,随之蒸腾速率下降,这样可以保护叶片不被灼伤、维持体内水分平衡,也是沙木蓼适应干旱、自我保护的一种方式。根据整个生长季(5~8月)所采集的数据,经统计分析,建立沙木蓼叶水势和蒸腾速率之间的回归模型为:LWP=-1.932-0.042 Tr,R2=0.574,n=28,F=34.966,sigf=0.000。同时相关性分析表明:在0.01水平下,R=0.757,则叶水势与蒸腾速率之间有极显著的负相关。
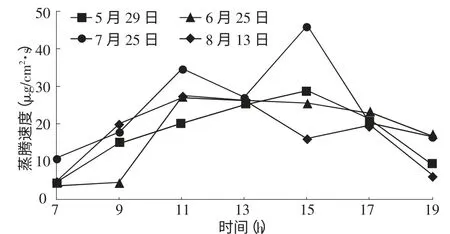
图2 沙木蓼蒸腾速率日变化
2.3 沙木蓼叶水势与各气象因子的关系
叶水势是反映叶片细胞水分状况的一个重要指标,叶水势越小叶片细胞越缺水,它的吸水能力越强,因此叶水势的变化可以反映植物适应干旱环境的能力[10]。本文以代表性较好的7月为例分析气象因子对沙木蓼叶水势影响规律。
2.3.1 叶水势与气温的关系 从图3叶水势和气温的变化曲线来看,两者有着相反的变化趋势。即早晨气温较低,叶水势较高,但随着气温的不断升高,叶水势开始逐渐降低。15∶00~17∶00为一天中气温最高的时段,最高温度27.9℃,15∶00左右叶水势降到最低值-3.91 MPa。17∶00之后,由于温度不断降低,植物蒸腾光合等各项生理活动开始减缓,叶水势回升。
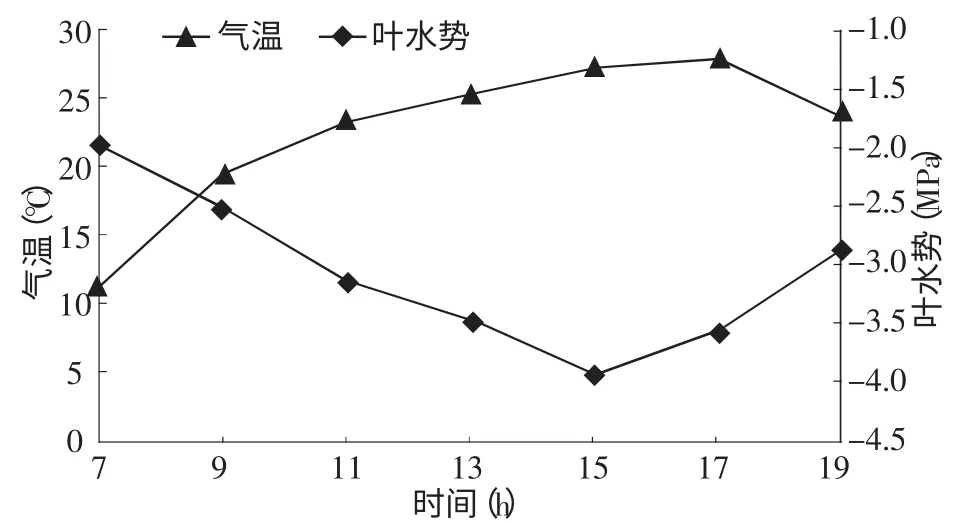
图3 叶水势与气温的关系
2.3.2 叶水势与大气相对湿度的关系 从图4叶水势与大气相对湿度的变化曲线可以看出,一天中清晨大气相对湿度最高,值为84%,此时叶水势也处在最高时段,值为-1.98 MPa;随着相对湿度不断降低植物叶水势也不断降低,15∶00~17∶00左右大气相对湿度与叶水势都降到最低值,分别为35%、-3.91 MPa。之后空气湿度回升,叶水势开始回升。
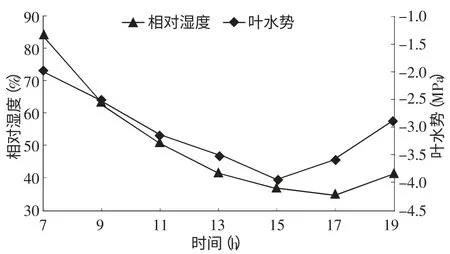
图4 叶水势与大气相对湿度的关系
2.3.3 叶水势与辐射强度的关系 从图5叶水势与辐射强度的变化曲线可以看出,清晨随太阳高度角不断增大,光照不断增强,叶水势不断降低。光照强度在13∶00左右出现最大值为765μmol/m2·s,叶水势则在15∶00左右出现最低值为-3.91 MPa。在13∶00~15∶00,光照强度继续降低,但叶水势并未升高,这表明叶水势相对于光照强度而言,存在明显的滞后性。
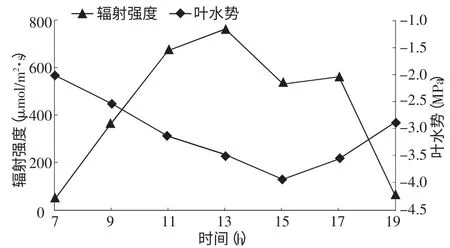
图5 叶水势与辐射强度的关系
用日过程对应时间所测得的叶水势平均值(LWP,MPa)分别与对应气候因子:大气温度(T,℃)、相对湿度(RH,%)、辐射强度(PAR,μmol/m2·s)进行拟合,关系模型见表2。从表2中可以看出,叶水势与光辐射强度的相关性较大气温度、相对湿度低,说明温度和相对湿度对叶水势变化的影响大于太阳辐射的影响。
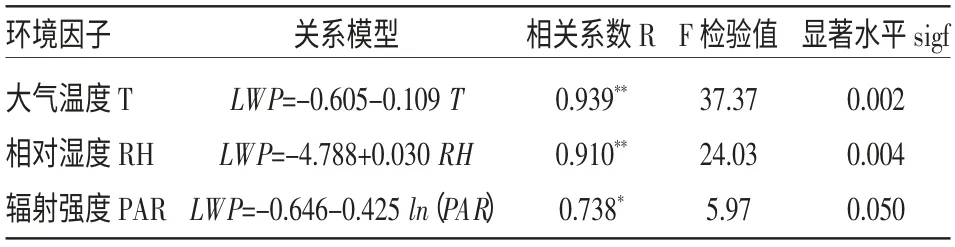
表2 叶水势日变化与单个环境因子的关系模型
2.4 沙木蓼各月叶水势与综合气象因子的关系
叶水势(LWP)与大气温度(T)、大气相对湿度(RH)及光辐射强度(PAR)三者之间有显著的相关关系。表3对三者综合因素与叶水势的关系作了分析。从沙木蓼整个生长季(5~8月)来看,综合因素对叶水势的影响要高于单因子对叶水势的影响。经过所观测的资料进行多元回归分析得到沙木蓼生长季(5~8月)的最优拟合方程LWP=-1.687-0.051 T+0.002 RH-0.001 PAR,其相关系数R=0.885,显著水平Sigf=0.00,n=28。

表3 各月叶水势与综合气象因子的关系模型
2.5 沙木蓼清晨叶水势与土壤含水量的关系
在充分供水条件下叶水势仅与气象条件有关,但当土壤水受到限制时,作物根系吸水满足不了蒸腾需要,此时叶水势与土壤含水量有着密切关系。为得到土壤含水量较低时植物叶水势的变化状况,在6月份进行盆栽沙木蓼充分灌水后的干旱胁迫试验,到苗木叶子出现萎蔫结束,前后持续了35 d。图6为这一过程中清晨叶水势随土壤含水量的变化曲线,由图可以明显看出沙木蓼叶水势随土壤含水量的降低而降低,这说明在相同天气条件下,随着土壤含水量的降低植物根系吸水速率越来越小,直至满足不了叶片蒸腾失水的需要,叶水势开始降低。经过回归分析,沙木蓼叶水势与土壤含水量之间有显著的正相关性,相关系数R=0.972,F=134.89,Sigf=0.000,回 归 方 程 为LWP=-3.917+0.716ln(SWC),式中:LWP为叶水势,单位为MPa;SWC为土壤含水量,单位%。
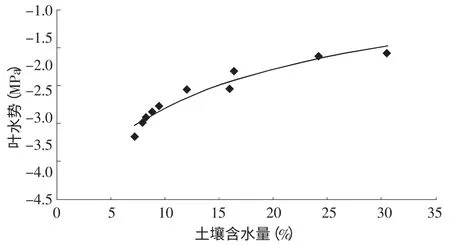
图6 清晨叶水势与土壤含水量的关系
3 结论
(1)沙木蓼叶水势日变化在6、8月份呈双峰曲线,5、7月呈单峰曲线;每日最高水势值出现在黎明前后,均值为-2.01 MPa;最低值出现在正午前后,均值为-3.40 MPa。沙木蓼叶水势月变化与植物生长发育阶段有密切关系,5月为沙木蓼生长初期,植物体内含水量较高,基本能满足其自身需要,叶水势也维持在较高的水平;6~7月,植物生长旺盛期,植物需水量较大,虽然集中降雨对缓解植物水分亏缺有所帮助,但是生理需水量和耗水量均大幅度增加,故叶水势不断降低。这也说明6~7月的水分状况对沙木蓼的生长起着关键作用。
(2)沙木蓼叶水势的变化主要取决于叶片水分的供求关系,即根系的供水能力与叶片的蒸腾速率。从整个生长季看,叶水势与蒸腾速率的变化趋势呈负相关。当蒸腾速率大于吸水速率时。蒸腾速率对叶水势起决定作用,即蒸腾速率升高。叶水势降低;反之亦然。
(3)叶水势作为SPAC系统中介于大气和土壤中间部分,叶水势的变化必然受两者的影响与控制。本文研究表明,在充分灌水条件下,沙木蓼叶水势与大气温度和光辐射强度呈负相关,相关系数分别为0.939,0.738,与大气相对湿度呈显著正相关,相关系数为0.910。而且,从沙木蓼整个生长季来看,综合气象因子对叶水势的影响要高于单个因子对叶水势的影响。当土壤水受到限制时,沙木蓼清晨叶水势与土壤含水量有显著正相关性,相关系数为0.972,Sigf=0.000。
[1] 孙丽娟.一种优良的固沙植物——沙木蓼[J].林业科技通讯,2001,10:38-39.
[2] 耿生莲,王占林.霸王、沙木蓼和银水牛果在不同水分条件下的生理研究[J].山西林业科技,2007,4:24-30.
[3] 郭其强,郭连金,次仁旦增,等.水柏枝的根系特征及固沙效果研究[J].安徽农业科学,2010,(28):15786-15787,15789.
[4] 李玉俊.对沙区优良薪材沙木蓼推广应用可能性的探讨[J].甘肃林业科技,1991,2:4-8.
[5] 王 北,王 谋,等.沙木蓼的引种与栽培[J].宁夏农林科技,1990,3:24-25.
[6] Philip J R.Plant water relations some physical aspects[J].Ann Rev.of Plant Physiol,1966,(17):245-268.
[7] 王沙生,高荣孚,吴贵明.植物生理学[M].北京:中国林业出版社,1990.
[8] 柴宝峰,李洪建.刺槐和海红水势与环境因子的关系及其抗旱性研究[J].山西大学学报(自然科学版),1996,19(2):223-227.
[9] 段焰青,李天星.云南松松针水势日变化的研究[J].云南大学学报(自然科学版),1999,21(5):410-412.
[10]刘玉燕,王艳荣,杨迎春.半干旱地区草坪草和主要杂草水势的日变化特征分析[J].内蒙古大学学报,2003,5,34(3):308-311.
