周边γ-氨基丁酸通过GABAB受体调控骶髓后联合核神经元谷氨酸能突触
2011-05-29马红雨吕建晓
马红雨,林 凯,罗 丹,吕建晓,杨 鲲
(1.空军总医院临床检验中心,北京 100142;2.第四军医大学解剖学教研室暨梁球锯脑研究中心,陕西 西安 710032;3.美国马里兰大学生理系,巴尔地摩 21201,美国)
骶髓后联合核(sacral dorsal commissural nucleus,SDCN)位于脊髓灰质腰骶段中央管后外侧。在大鼠,SDCN出现在腰6(L6)至骶4(S4)段,是接受躯体感觉和内脏感觉初级传入的重要部分,并将初级传入信息向对侧脊髓以上核团投射[1]。谷氨酸是哺乳动物中枢神经系统最重要的兴奋性神经递质。SDCN神经元接受广泛的谷氨酸能传入[1]。而γ-氨基丁酸(GABA)是中枢神经系统主要的抑制性神经活性物质。在脊髓,GABA受体分为离子型GABAA受体和代谢型GABAB受体两种[2]。在成年哺乳动物,GABAA受体被激活后,引起氯离子(Cl-)通道开放,导致胞外的Cl-内流,神经元超极化,进而抑制神经元的活动[3]。与GABAA受体等离子门控型受体不同,GABAB受体属于具有7个跨膜区段并与胞内G蛋白偶联的代谢型受体[3]。形态学研究表明[4-5],GABAB受体在脊髓背角和SDCN均有广泛表达。在中枢神经系统和脊髓,于突触结构(突触前膜、突触后膜、突触间隙)外,存在一定水平的周边神经活性物质(ambient neuroactivities)。这些神经活性物质,可能参与突触递质释放的调节[6]。在脊髓背角和 SDCN,GABAB受体被其外源性受体激动剂激活,进而调节突触前递质释放和突触后神经元兴奋性已有很多研究[4,7-8]。但存在于突触周边的内源性GABA是否通过对谷氨酸能突触上的GABAB受体的影响,进而调节谷氨酸释放,尚不清楚。本实验拟用电生理学方法研究上述问题。
1 材料与方法
1.1标本制作实验动物为5~8周成年♂Sprague-Dawley(SD)大鼠。脊髓薄片制作方法如前所述[7,9-10]:用乌拉坦将动物腹腔注射麻醉(1.5 g·kg-1体质量),沿背正中线由骶髓水平向上至胸髓水平剪开皮肤,以组织剪沿椎骨棘突两侧剪开肌肉,暴露横突,行椎板切除术,小心暴露脊髓。将脊髓于胸段水平横断,向下至全骶髓连同蛛网膜和软膜一并取出,迅速置于冷的人工脑脊液(artificial cerebrospinal fluid,ACSF)中。除去蛛网膜和软膜,修剪去前根和背根。振动切片机(DTK-1 000,Dosaka EM,日本)切取厚度为500 μm的腰骶部位脊髓横切薄片。将标本转移到记录槽中,ACSF液持续灌流(速度~10 ml·min-1;(36±1)℃。预孵育30 min开始记录。
1.2药品与溶液GABAB受体特异性拮抗剂3-[[[(3,4-dichlorophenyl)methyl]amino]propyl](diethoxy-methyl)phosphinic acid(CGP52432),河豚毒素 (tedrodotoxin,TTX),6-cyano-7-nitroquinoxaline-2,3-dione(CNQX),GABAA受体拮抗剂 picrotoxin和胞内G蛋白偶联受体阻断剂guanosine 5'-[β-thio]diphosphate trilithium salt(GDP-βS)均购自 Sigma公司(美国)。其他药品购自试剂公司。ACSF成分为(mmol·L-1):NaCl 117,KCl 3.6,CaCl22.5,MgCl21.2,NaH2PO41.2,NaHCO325,glucose 11,picrotoxin 0.05。记录电极细胞内液成分为(mmol·L-1):K-gluconate 135,KCl 5,CaCl20.5,MgCl22,EGTA 5,Hepes 5,Mg-ATP 3.6,GDP-βS 1。电极内液中的GDP-βS能够抑制被记录神经元(突触后神经元)胞内G蛋白,以排除突触后GABAB受体的作用[7-8]。
1.3全细胞记录与给药方式标本置于灌流槽内,以一银丝缠绕细尼龙线作成的“U”字型锚固定薄片。以“盲插法”(blind patch)形成全细胞电压钳记录,封接方法如另文详述[9]。在钳制电压为 -70 mV时,一般可以观察到自发的兴奋性突触后电流(spontaneous excitatory postsynaptic currents,sEPSCs)。当外液加入河豚毒素 (0.5 μmol·L-1)时,可记录到的微小突触后电流 (miniature EPSCs,mEPSCs)[7,9]。使用同心圆电极给予点刺激(focal stimulation)时,可直接激活突触前终末,引起兴奋性突触后电流 (evoked excitatory postsynaptic currents,eEPSCs)[11]。模式/数字信号转换系统和记录软件分别为 Digidata 1200A analog/digital interface和pClamp 6(均购自Axon Instruments)。
实验前将药物用蒸馏水或二甲基亚砜(DMSO)制成1 000~5 000倍浓度储备液。使用前,以ACSF稀释至终浓度。采用灌流方式给药,分别记录给药前(对照组),给药后(实验组)及洗脱后的电流变化。
1.4数据处理和统计学分析实验结果用Axo-Graph 4.0(Axon Instruments)或Clamp Fit 8.0(Molecular Devices,美国)分析。数据在Stat View(Apple McIntosh,美国)或 Sigma Plot 9.0(Sigma Plot,美国)上进行计算和统计学比较。数值以±s表示。n代表实验的细胞数目。结果处理用Student’st-test(t检验)、χ2检验或 Kolmogorov-Smirnov test(K-S检验)。
2 结果
2.1抑制突触前GABAB受体导致刺激引起的兴奋性突触后电流(eEPSCs)幅度增加钳制电压为-70 mV 时,突触前刺激(100 μs,0.033 Hz)在一定强度下(3~20 V),可诱导出稳定的兴奋性突触后电流eEPSCs。记录5 min稳定的对照电流后,灌流GABAB受体特异性受体阻断剂CGP52432(1 μmol·L-1,8 ~ 10 min)[3]。如 Fig 1A 所示,CGP52432增加eEPSCs的幅度。在所有被观察的11个神经元上,CGP52432灌流8 min增加eEPSCs的幅度至对照水平的(141±18)%(P<0.05,pairedt-test)(Fig 1B)。CGP52432的易化作用,经洗脱后可部分恢复。
我们的既往研究表明[4,7,9],GABAB受体特异性激动剂氯苯氨丁酸(baclofen)减少微小兴奋性突触后电流mEPSCs。本实验进一步观察到,CGP52432(1 μmol·L-1,10 ~15 min)对 mEPSCs的频率或幅度分布均无影响(K-S检验):平均频率在对照组为(3.1±1.0)Hz,CGP52432灌流后为对照数值的(107±10)%;平均幅度在对照组为(21.3±8.2)pA,CGP52432处理后为对照水平的(99±7)%(两组均为P>0.05,t检验,n=13)(Fig 1C)。以上结果表明,以CGP52432阻断谷氨酸能突触前GABAB受体,导致eEPSCs幅度增加(即谷氨酸释放增加),但是对自发微小突触电流无明显影响。
2.2突触前GABAB受体参与eEPSCs幅度增加为进一步确认CGP52432是否作用于突触前GABAB受体,我们选用了同心圆电极间隔刺激法:当两次刺激间隔为50 ms、频率为0.033 Hz、钳制电压为-70 mV时,可以观察到同心圆刺激电极刺激引起的配对刺激比率(paired-pulse ratio;PPR)(Fig 2A)。对照组eEPSCs的PPR(P2/P1)为(0.78±0.07),灌流 CGP52432(1 μmol·L-1,8 ~10 min)使PPR值变为0.65±0.09,较对照值增大(P<0.05,n=12)(Fig 2B)。提示CGP52432增加刺激引起的谷氨酸释放的作用部位是突触前GABAB受体。
2.3抑制突触前GABAB受体导致沉默突触功能恢复上述结果表明,阻断突触前GABAB受体可增加谷氨酸释放,本实验进而研究该效应是否可以将谷氨酸能的沉默突触(silent synapse)变成活跃突触(active synapse)。我们首先寻找沉默突触。方法如前所示[12]:以同心圆电极进行双刺激(间隔为50 ms),在一定强度下,可找到第1个刺激冲动诱发eEPSCs;逐渐降低刺激强度,一般在阈强度90%左右,可找到符合下列标准的沉默突触:①第1个eEPSCs至少连续20次刺激经目视检查,无可分辨的幅度(沉默突触);② 第2个eEPSCs偶然出现。此种类型沉默突触一般认为是因为第1个刺激引起的递质释放不足而引起的“突触前沉默”[12]。如Fig 3A所示,沉默突触在灌流CGP52432后部分恢复功能成为活跃突触 (active synapse)。CGP52432引起第1个突触的平均幅度和诱发eEPSCs成功率均有明显增高(Fig 3B)。在13个被检测的“突触前沉默”的神经元中,CGP52432灌流8 min后,11例转变为活跃突触(P<0.05,χ2检验)。

Fig 1 Evoked excitatory postsynaptic currents in SDCN neuron enhanced by blockade of GABABreceptors by CGP52432
3 讨论
本研究通过用药理学方法先阻断突触后SDCN神经元 GABAB受体,排除突触后作用后,再以CGP52432阻断位于谷氨酸能突触前的GABAB受体,观察到刺激引起的谷氨酸释放增加,且此易化作用发生在突触前。结果提示,在SDCN,突触周边存在的GABA通过GABAB受体,对谷氨酸释放起限制作用。

Fig 2 Paired-pulse ratio decreased by CGP52432.
大部分神经递质在神经元胞体合成后,运输至突触前末梢,参与递质释放。被释放的递质部分与突触后膜上的受体结合,部分经过重摄取(re-uptake)再次进入突触前膜,还有部分溢出(spillover)突触结构,成为周边神经活性物质。分布在突触周边存在的抑制性神经活性物质对突触起着紧张性调节(tonic modulation)作用。周边GABA的来源,至少有两个:除上述的溢出外,最新研究表明胶质细胞也可能释放GABA[13]。本研究中,尚不能判断周边GABA的具体来源。
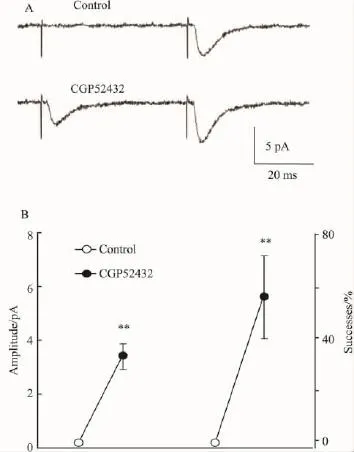
Fig 3 A presynaptic silent neuron becoming active to application of CGP52432
经典神经电生理学通过对PPR的分析,可以判定某种特定药物的作用部位(突触前或突触后)。在PPR实验中,由于两次刺激间隔的时间短,实施第2次刺激时,突触前已经流入的Ca2+还处于较正常为高的水平,影响第2次刺激引起的Ca2+内流,因而第2次刺激引起的变化小。给予某种药物时,如果对第1次刺激的相对作用大于对第2次刺激的相对作用,则可以判断为突触前作用。本实验中,CGP52432明显降低PPR的比率,因而推断增加谷氨酸释放的作用位点为突触前GABAB受体。令人感兴趣的是,CGP52432阻断GABAB受体,却对静息条件下的微小突触电流(mEPSCs)没有影响(Fig 1C),提示上述的调节作用只存在于突触前动作电位诱发的谷氨酸释放。
需要指出的是,脊髓SDCN神经元的突触前和突触 后 均 有 GABAB受 体 分 布[4-5],因 而 用CGP52432可能对突触前后的GABAB受体后都产生作用。在本实验中,突触后GABAB受体的作用被通过记录电极施与的 GDP-βS 阻断[4,8,11],因而可以排除GABAB受体的突触后作用。
SDCN接受广泛的初级传入,其中部分传入介导伤害性躯体感觉与内脏感觉信息。这些信息经过SDCN,由投射神经元再进一步向延髓和丘脑等目标核团投射[1]。本研究中发现的周边 GABA通过GABAB受体影响谷氨酸的释放,很可能参与这种伤害性信息传递的调制。GABAB受体在影响突触前兴奋性谷氨酸释放的同时,也能调节抑制性神经递质,如GABA自身和甘氨酸(glycine)的释放(杨鲲等,待发表资料)。而抑制性递质释放的减少被认为对突触后神经元有间接的兴奋作用,因而周边GABA对SDCN信息传递调节的具体机制,对信息加工的“净作用”(net effect),尚需进一步研究确认。
脊髓作为伤害性信息传递与调控的第一站,有许多位点和可作为镇痛药的靶位。最近的研究表明,蛋白激酶C(protein kinase C,PKC)参与脊髓水平炎性疼痛的表达[14]。因而,影响PKC的药物,可能有潜在的药理学价值。N-甲基-D-天冬氨酸(N-methyl-D-aspartate,NMDA)受体NR2B亚型也有类似的作用[15]。长期以来,GABAB受体作为新型药物开发的热点,主要集中在如何开发其配体(激动剂及变构体)[2-3]。本研究观察到,内源性周边GABA作用于谷氨酸能突触上的GABAB受体,且只参与动作电位(突触前刺激)引起谷氨酸释放调节,对开发新型镇痛药物将有重要的参考价值。
[1]李继硕.脊髓[M].//李继硕.神经科学基础.北京:高等教育出版社,2002:116-44.
[1]Li J S.Spinal cord[M]//Li J S.Basic Neuroscience.Beijing:High Education Press,2002:116-44.
[2]Malcangio M,Bowery N G.GABA and its receptors in the spinal cord[J].Trends Pharmacol Sci,1996,17(12):457-62.
[3]Bowery NG,Bettler B,Froestl W,et al.International Union of Pharmacology.XXXIII.Mammalian γ-aminobutyric acid B receptors:structure and function[J].Pharmacol Rev,2002,54(2):247-64.
[4]Yang K,Wang D,Li Y Q.Distribution and depression of the GABABreceptor in the spinal dorsal horn of adult rat[J].Brain Res Bull,2001,55(4):279-85.
[5]Margeta-Mitrovic M,Mitrovic I,Riley R C,et al.Immunohistochemical localization of GABABreceptors in the rat central nervous system[J].J Comp Neurol,1999,405(3):299-321.
[6]Kirmse K,Kirischuk S.Ambient GABA constrains the strength of GABAergic synapses at Cajal-Retzius cells in the developing visual cortex[J].J Neurosci,2010,26(16):4216-27.
[7]马红雨,杨鲲.氯苯氨丁酸抑制脊髓背角神经元谷氨酸释放的机制[J].中国药理学通报,2004,20(9):981-4.
[7]Ma H Y,Yang K.Mechanism for baclofen inhibition on quantal glutamate release in spinal dorsal horn neurons[J].Chin Pharmacol Bull,2004,20(9):981-4.
[8]Yang K,Kumamoto E,Furue H,et al.Pre-and postsynaptic inhibition mediated by GABABreceptors in rat ventrolateral periaqueductal gray neurons[J].Biochem Biophys Res Comm,2003,302(2):233-7.
[9]Yang K,Li Y Q,Kumamoto E,et al.Voltage-clamp recordings of postsynaptic currents on substantia gelatinosa neuronsin vitroand its applications to assess synaptic transmission[J].Brain Res Protocols,2001,7(3):235-40.
[10]Yang K,Feng Y P,Li Y Q.Involvement ofN-methyl-D-aspartate receptor in the C-fiber-evoked excitatory postsynaptic currents in substantia gelatinosa of adult rat spinal cord slice[J].Chin Pharmacol Bull,2000,16(special issue):30-3.
[11]Yang K,Fujita T,Kumamoto E.Adenosine inhibits GABAergic and glycinergic transmission in adult rat substantia gelatinosa neurons[J].J Neurophysiol,2004,92(5):2867-77.
[12]Dobrunz L E,Stevens C F.Heterogeneity of release probability,facilitation,and depletion at central synapses[J].Neuron,1997,18(6):995-1008.
[13]Lee S,Yoon B E,Berglund K,et al.Channel-mediated tonic GABA release from glia[J].Science,2010,330(6005):790-6.
[14]索占伟,杨 娴,曹 静,等.脊髓背角PKC在慢性炎性疼痛中的作用及其机制[J].中国药理学通报,2011,27(3):316-9.
[14]Suo Z W,Yang X,Cao J,et al.Involvement of spinal PKC in inflammatory pain and its underlying mechanisms[J].Chin Pharmacol Bull,2011,27(3):316-9.
[15]傅 璐,郭瑞先,莫利求,等.脊髓c-Jun在NMDA受体NR2B亚基介导的大鼠吗啡镇痛耐受中的作用[J].中国药理学通报,2011,27(2):248-52.
[15]Fu L,Guo R X,Mo L Q,et al.Role of c-Jun of spinal cord in NMDA receptor subunit NR2B mediated morphine antinociceptive tolerance in rats[J].Chin Pharmacol Bull,2011,27(2):248-52.
