不同草地植被群落的结构特征研究
2011-04-25郭君喜张鹏莉胡远彬
吕 杰,郭君喜,陈 俊,张鹏莉,胡远彬
(西北农林科技大学动物科技学院,陕西 杨凌 712100)
草地是畜牧业发展的重要物质基础和农牧民赖以生存的基本生产资料,近些年由于全球气候变暖及长期以来草地开垦、超载过牧、滥采乱挖、违法征占等自然和人为因素,草地植被遭到严重破坏,草地生态环境持续恶化[1]。草地退化不是生态环境破坏的根本原因,而是生态环境破坏的结果。尽管我国草地保护建设近年来取得了较大成就,截至2005年,我国已有多个县实行了封山禁牧、舍饲养畜,其中北京、河北、陕西、青海、宁夏5个省(区、市)实行了全境禁牧[2],但草地生态环境“局部改善、总体恶化”的局面仍未根本改观[1]。目前,全国90% 的可利用天然草地不同程度的退化[3],陕西榆林地区作为我国半干旱农业区向干旱草地区的过渡地带,在自然地理上有其独特的空间分布,在生态上具有显著的脆弱的生境特征和演化规律[4]。近些年,大量煤焦化小工厂的建立,严重破坏了陕西省神木县的草地和生态环境。因此,开展草地生态系统多样性及其空间分布规律的专题研究,在草地实际生产、草地保护研究中具有一定的意义和重要的学术价值。
围栏禁牧能够保持物种的多样性,增强生态系统的稳定性;能够有利于适宜当地植物种的生长发育,增强草群的抗逆性;是恢复退化草地的有效途径之一[1,5-6],已经在研究与实践中得到了广泛应用。因此,对植被群落结构特征的研究极其重要,所以本研究以植被群落结构特征为切入点,通过对草地植物群落物种多样性和空间异质性的定量研究,再运用β-二项分布模型的解析方法[7-9],对草地植被的调查结果进行分析,探究黄花蒿(Artemisiaannua)草地、羊草(Leymuschinensis)草地和百里香(Thymusmongolicus)草地的物种数、物种出现频率及其空间分布的基本规律。以期通过草地植被空间异质性的定量研究,为半干旱草地的合理利用和科学管理提供理论依据。
1 材料与方法
1.1调查地概况 调查地位于陕西省榆林市神木县的中鸡镇栅子沟村(110°10′28″ E,39°11′13″ N;海拔1 254 m)、孙家岔镇柳树峁村(110°12′24″ E,39°08′55″ N;海拔1 251 m)和石场峁村(110°15′07″ E,39°10′03″ N;海拔1 234 m),分别属于黄花蒿草地、羊草草地和百里香草地3种典型草地类型(均已禁牧5年)。神木县地处陕西省最北端,是黄土高原与内蒙古鄂尔多斯荒漠草地过渡地带,是陕西省面积最大的县。神木县属温带半干旱大陆性季风气候,主要特点是:春多风、夏干旱、秋阴雨、冬严寒,日照充足,风沙频繁,雨季迟且雨量年际变化大[4]。县境北部和中部为长城沿线风沙草滩区,属半荒漠草地,南部为黄土高原丘陵沟壑区,属干草地;海拔738.7~1 448.7 m;全县年平均气温8.5 ℃,极端最低气温-28.1 ℃,极端最高气温38.9 ℃;年平均降水量440.8 mm,70%以上集中于7-9月;相对湿度平均为55%;无霜期平均169 d[10]。
1.2调查方法与时间 野外调查于2010年9月5-10日进行,首先在每种草地上分别选择有代表性的样地,每个样地上各拉一条45 m长的样线,然后顺着样方线摆放90个50 cm×50 cm的铁制样方框,称为大样方(以下简称为L-样方),将L-样方均等地分割为4个25 cm×25 cm的小样方(以下称为S-样方)。记录S-样方内出现的所有植物种。如果植物群落内某种植物在L-样方内的4个S-样方中都没出现时,这种植物在L-样方内的出现次数为0,如果只在1个S-样方内出现,那么这种植物在L-样方内的出现次数就为1,以此类推,如果该植物在L-样方内的4个S-样方内都出现,那么它在L-样方内的出现次数就为4,得到出现次数数据。最后沿着样线每隔5 m将样方内(每条样线测定9个样方)的所有植物齐地面剪下,带回实验室,在70 ℃恒温干燥箱内烘干至质量不变,得到生物量数据。
1.3数据解析方法 本研究采用的数据解析方法为β-二项分布模型,它可以简单表述为在植物种A存在的场所B内,放置一定面积的多个样方,然后把每个样方均等地分为n个等面积的小样方。假设A在B内空间分布是不均一的,即处于异质性分布状态,那么,在一定面积内A所占据的小样方数i遵循β-二项分布[8,11-13]。i个小样方被植物种A所占据的概率p(i)可用下式来表示:
式中,B(α,β)是包含参数α和β的β函数(α,β>0),α,β是决定β-二项分布形状的两个参数。β函数可用下式表示:
另外,植物种A在L-样方内的0,1,2,…,n个S-样方内出现的频率p(0),p(1),p(2),…,p(i),…,p(n)可用以下的公式来表示:
式中,p为植物种A出现的S-样方数在所调查的所有L-样方内的S-样方总数中所占的比例,i=1,2,3,…,n。
式中,n为L-样方内的S-样方数,本调查中n=4;μ为平均数;σ2为L-样方内出现次数的方差。
根据α和β的取值不同,β-二项分布模型有5种基本情况(图1)。它们有直线型、“U”字型、“山峰”型、“倒J”型、“J”字型(分别如图1a、b、c、d、e)。图1a表示随机分布,其他均为程度不同的异质性分布,在实际的草地植物群落中完全的均匀分布很少存在。比如,某种植物在调查的100个L-样方中,都以1次或者2、3、4次出现,这种情况属于均匀分布。
1.4空间分布的异质性指数
1.4.1组成群落各个植物种的空间异质性 如果用vi表示每种植物在L-样方内实测出现次数的方差,mi表示植物种在L-样方内实测出现次数的平均数,pi表示某植物种i的实测出现频率,ρi表示某植物种i的空间分布异质性指数。那么空间分布的异质性指数ρi就可用下式来定义[8]:
式中,pi=mi/n;0≤pi≤1;-1/3≤ρi≤1。
空间异质性程度的判断标准如下:
当ρi=0时,表示植物种在草地上呈现随机分布状态。
当ρi>0时,表示比随机分布具有强的空间异质性分布,越接近于1,异质性程度越大。
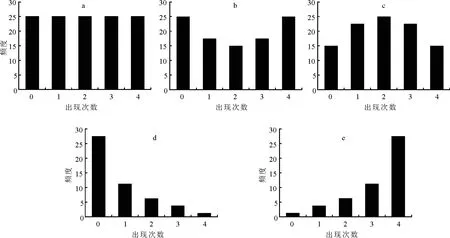
图1 β-二项分布模型的5种情况
当ρi<0时,表示植物种比随机分布具有弱的空间异质性,越接近于-1/3,越趋近于均匀分布。
1.4.2群落整体的空间异质性指数 群落整体的空间分布异质性指数ρc是在综合考虑组成群落的各个植物种的出现频率与其空间异质性程度的大小的基础上得出的,用下式表示[8,14-15]:
式中,S为群落内出现的植物种数。根据异质性指数ρc对植物群落整体异质性程度的评价标准与ρi相同。
1.5物种多样性指数及均匀度指数 物种多样性指数采用Shannon-Wiener的H′指数,用下式表示:
式中,pi′为出现频率pi的相对值,用下式计算:
均匀度指数采用Pielou[16]的J′均匀度指数,计算式如下:
2 结果与分析
2.1草地组成植物种对β-二项分布模型的吻合性 表1表示3种草地的所有植物种对β-二项分布模型的吻合性检验结果,其中黄花蒿草地共出现24种植物,利用卡方检验得到的结果,有22种植物实测值的出现频度与利用β-二项分布模型计算得到的理论频度之间没有显著性差异(P>0.05),符合β-二项分布模型,只有内蒙古西风芹(Seseliintramongolicum)和草木樨状黄耆(Astragalusmelilotoides)两种植物实测值的出现频度与理论频度之间存在着显著性差异(P<0.05),说明这两种植物不符合β-二项分布模型;百里香草地植物种共出现31种,其中有29种植物实测值的出现频度与利用β-二项分布模型计算得到的理论频度之间没有显著性差异(P>0.05),符合β-二项分布模型,只有莎草(Cyperaceaesp.)、老鹳草(Geraniumwilfordii)两种植物实测值的出现频度与理论频度之间存在着显著性差异(P<0.05),说明这两种植物不符合β-二项分布模型;羊草草地植物种共出现28种,其中有27种植物实测值的出现频度与利用β-二项分布模型计算得到的理论频度之间没有显著性差异(P>0.05),符合β-二项分布模型,只有阿尔泰狗娃花(Heteropappusaltaicus)实测值的出现频度与理论频度之间存在着显著性差异(P<0.05),说明这种植物不符合β-二项分布模型。

表1 对β-二项分布模型的吻合检验结果
综上所述,3种草地植物群落的构成植物种总体上符合β-二项分布模型,即可利用β-二项分布模型对3种草地植物群落的空间分布进行解析。
2.2草地的群落总体特征 3种草地共同的植物种有16种,总物种数表现为黄花蒿草地<羊草草地<百里香草地;物种多样性指数表现为羊草草地<黄花蒿草地<百里香草地;均匀度指数表现为羊草草地<百里香草地<黄花蒿草地;L-样方内的平均物种数和L-样方内的物种多样性指数均表现为羊草草地<百里香草地<黄花蒿草地;群落整体的空间分布异质性指数表现为黄花蒿草地<羊草草地<百里香草地;L-样方内的平均生物量表现为百里香草地<黄花蒿草地<羊草草地(表2)。
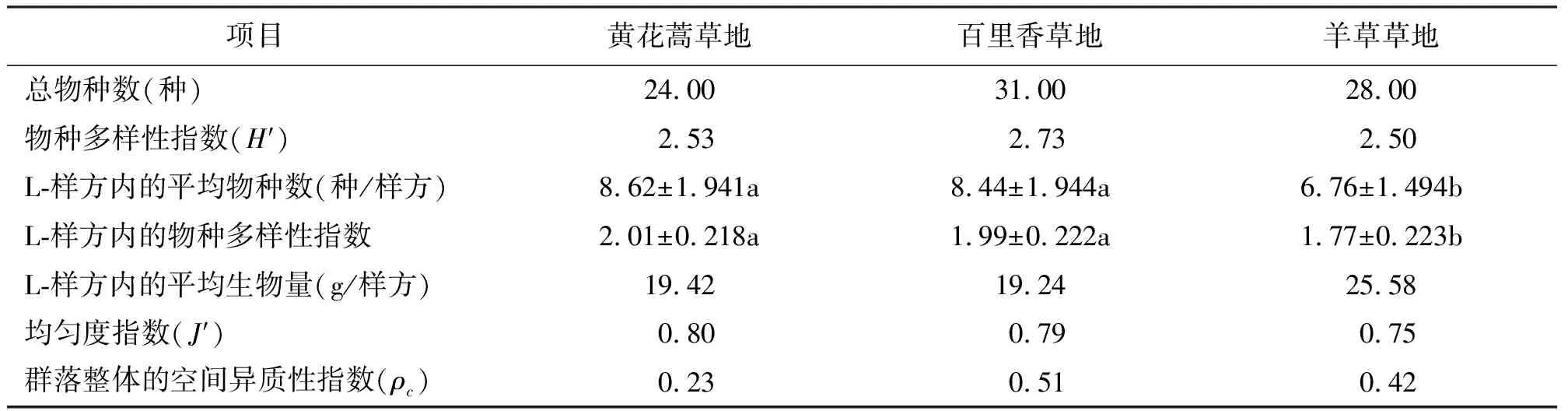
表2 群落总体特征
2.3组成群落的主要植物种 3种草地中出现频率排在前10位的共同植物种有4种,为黄花蒿、阿尔泰狗娃花、达乌里胡枝子(Lespedezadavurica)和草木樨状黄耆(表3)。主要构成植物种的出现频率(pi)大于0.6的植物种为优势植物种,黄花蒿草地中优势种有黄花蒿、远志(Polygalatenuifolia)、阿尔泰狗娃花和达乌里胡枝子,第5位的糙隐子草(Cleistogenessquarrosa)的出现频率已降到0.303。而百里香草地优势种发生了很大的变化,在黄花蒿草地中出现频率很低的糙隐子草和猪毛菜(Salsolacollina)跃居为百里香草地的优势种,其出现频率分别为0.686和0.658,而在黄花蒿草地中出现频率很高的黄花蒿和远志在百里香草地中出现频率跌为0.264和0.189。此外,百里香草地的优势种还有百里香和达乌里胡枝子。羊草草地的优势种有羊草和黄花蒿,从第3位植物种开始出现频率呈急剧下降趋势,而黄花蒿草地和百里香草地的组成植物种从第1位到第10位的出现频率呈现逐渐下降趋势。
2.4出现频率与空间分布异质性指数之间的关系 在黄花蒿草地中黄花蒿和远志的出现频率均大于0.90,并且异质性指数维持在0.2左右,使群落整体的空间异质性保持在较低水平;阿尔泰狗娃花、达乌里胡枝子异质性指数大于0.3,且出现频率大于0.60,使得群落整体的空间异质性维持在较高水平;其余出现频率均低于0.31,对群落空间异质性的影响并不大,特别是角蒿和白花草木樨(Melilotusalba)的空间异质性指数均接近于0,属于随机分布,但由于出现频率也接近于0,所以对群落空间异质性的影响很小(图2)。
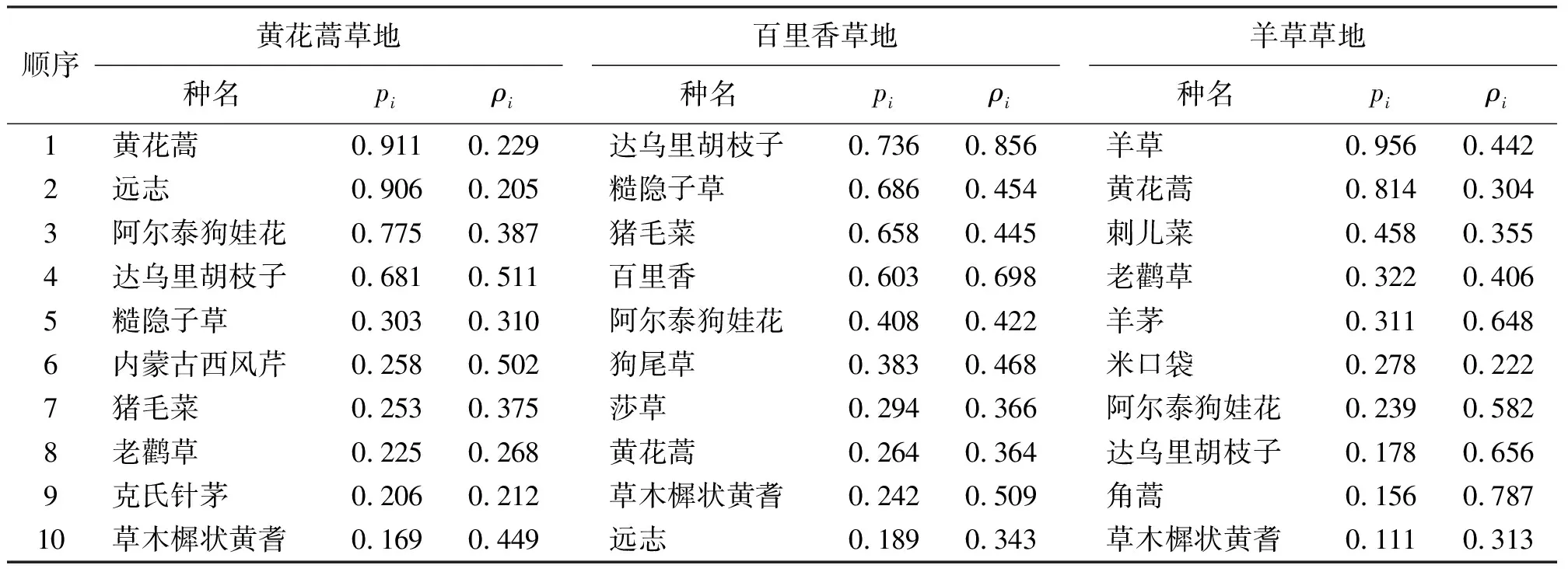
表3 3种草地中出现频率排在前10位植物种
在百里香草地,达乌里胡枝子、百里香、糙隐子草、猪毛菜4种植物出现频率大于0.6,且空间异质性指数大于0.4,它们大大提高了群落整体的空间异质性;阿尔泰狗娃花、狗尾草、草木樨状黄耆的出现频率和空间异质性指数均较高,使得群落维持较高的水平;异质性较高但出现频率较低的植物有油蒿(A.ordosica)和黄花草木樨(M.officinalis),对群落整体的空间异质性的影响很小;空间异质性指数接近于0,属于随机分布的植物冠芒草(Enneapogonborealis)、 地锦(Euphorbiahumifusa)、地梢瓜(Cynanchumthesioides)、虎尾草(Chlorisvirgata),但是出现频率也接近于0,所以对群落整体的空间异质性的作用也接近于零(图2)。
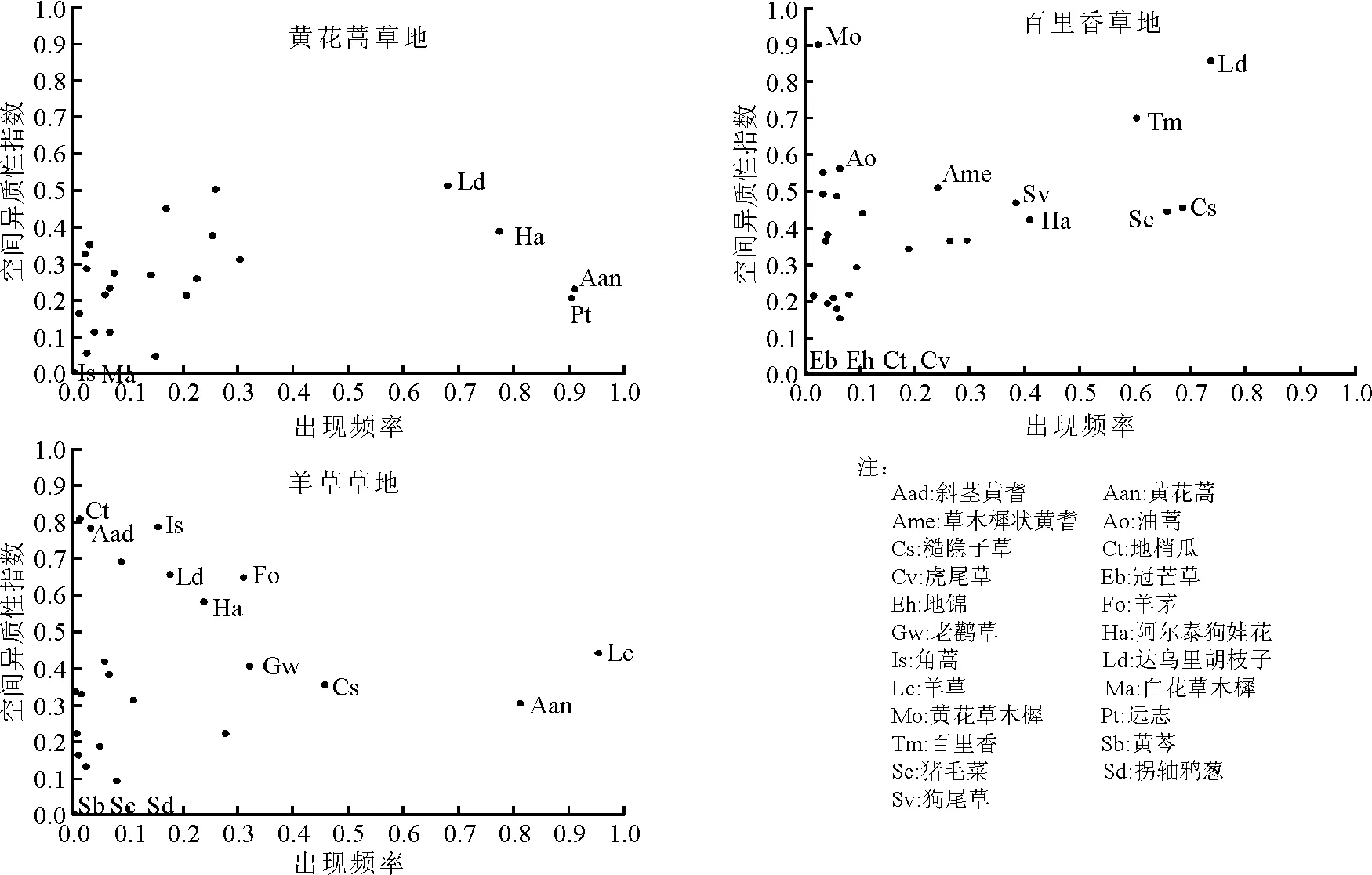
图2 出现频率(pi)与空间异质性指数(ρi)之间的关系
在羊草草地,使群落整体的空间异质性维持在较高水平的植物有羊草和黄花蒿;出现频率和空间异质性指数都处于中等水平,对提高群落空间异质性也具有一定的影响的植物有糙隐子草、老鹳草、羊茅、阿尔泰狗娃花和达乌里胡枝子等;地梢瓜、斜茎黄耆(Astragalusadsurgens)和角蒿的空间异质性指数很高(0.8左右),但是它们的出现频率却很低,因此对群落的空间异质性影响不大;黄芩(Scutellariabaicalensis)、猪毛菜和拐轴鸦葱(Scorzoneradivaricata)的空间异质性指数均接近于0,属于随机分布,但由于出现频率也接近于0,所以对群落空间异质性的影响很小(图2)。
3 讨论与结论
本研究所采用的野外调查方法与β-二项分布模型解析法相结合,通过卡方检验探明了3种草地的组成植物种对β-二项分布模型具有很好的吻合性。且利用β-二项分布模型,能定量地判明组成群落的所有植物种及其群落整体空间分布的异质性程度。本研究所采用的野外调查方法简单易行,且对草地植被没有破坏性影响,特别是对低草型草地的调查与研究更为适用[17-21]。
通过本项研究得出,3种草地共同的植物种有16种,总物种数顺序为黄花蒿草地<羊草草地<百里香草地,物种多样性指数为羊草草地<黄花蒿草地<百里香草地,均匀度指数为羊草草地<百里香草地<黄花蒿草地;0.25 m2样方内(大样方)的平均物种数和物种多样性指数均为羊草草地<百里香草地<黄花蒿草地,群落整体的空间分布异质性指数为黄花蒿草地<羊草草地<百里香草地,平均生物量表现为百里香草地<黄花蒿草地<羊草草地。黄花蒿草地和百里香草地属于梁地草地类型,其草地0.25 m2样方内的平均物种数和物种多样性指数差异不显著,且平均生物量和均匀度指数非常接近。总物种数和物种多样性指数在梁地草地之间比较时呈现了正相关性,群落整体的空间异质性指数百里香草地>黄花蒿草地,其原因是百里香草地中百里香和达乌里胡枝子两种优势植物都属于匍匐茎型营养繁殖的植物,对别的植物生长有很大的抑制性,而黄花蒿草地中黄花蒿和远志两种优势植物是种子繁殖的植物,所以空间异质性相对较小,这与陳俊和塩見正衛[20]、安田泰輔等[22]的研究结果相一致,即通常种子繁殖的植物异质性程度低,而营养繁殖的植物种异质性高。对梁地草地和滩地草地(羊草草地属于滩地草地)进行比较时发现,0.25 m2样方内的平均物种数和物种多样性指数差异极显著,滩地草地物种多样性指数和均匀度指数均低于梁地草地,而平均生物量却高于梁地草地,其原因是滩地草地水分含量相对较高,草地的植物总体长势很好,梁地草地受到水分的影响,植物相对比较稀疏。此外,羊草草地的群落整体的空间异质性指数相对较高,因为羊草属于根茎型禾草,可以大大提高群落整体的空间异质性。
本研究通过运用β-二项分布模型的数理解析方法对小尺度范围内的草地植物群落及其组成群落的各个植物种的空间异质性进行定量分析,为将来进一步地探明小尺度上草地群落空间异质性与生物量、物种多样性之间的关系做铺垫,通过异质性的定量研究,可为准确测算草地生产力,维持草地的物种多样性提供理论依据。
[1]刘荣.依托退牧还草项目鄂尔多斯实施禁牧、休牧、划区轮牧快速恢复草原生态[J].内蒙古草业,2010,22(4):14-17.
[2]孟昭丽.我国已有5个省 (区、市)实行了全境禁牧[J].草业科学,2006,23(7):53.
[3]邹声文.我国天然草原90%在退化[J].草业科学,2002,19(4):76.
[4]李忠锋,王一谋,冯毓荪.神木县近十年生态治理效果评价[J].遥感技术与应用,2004,19(5):348-352.
[5]张东杰,都耀庭.禁牧封育对退化草地的改良效果[J].草原与草坪,2006(4):52-54.
[6]杨晓晖,张克斌,侯瑞萍.封育措施对半干旱沙地草场植被群落特征及地上生物量的影响[J].生态环境,2005,14(5):730-734.
[7]Chen J,Hori Y,Yamamura Y,etal.Spatial heterogeneity and diversity analysis of macrovegetation in the Xilingol Region,Inner Mongolia,China,using the beta distribution[J].Journal of Arid Environments,2008,72(6):1110-1119.
[8]Shiyomi M,Yoshimura J.A measures of spatial heterogeneity for species occurrence or disease incidence with finite-counts[J].Ecological Research,2000,15(1):13-20.
[9]Chen J,Shiyomi M,Hori Y,etal.Frequency distribution models for spatial patterns of vegetation abundance[J].Ecological Modelling,2008,211(3/4):403-410.
[10]闫朝,方尚春.神木县草业发展的成就与前景[J].中国畜禽种业,2010(11):26-27.
[11]Skellam J C.A probability distribution derived from the binomial distribution by regarding the probability of success as variable between the sets of trials[J].Journal of the Royal Statistical Society.Series B (Methodological),1948,10(2):257-261.
[12]Kemp C D,Kemp A W.The analysis of point quadrat data[J].Australian Journal of Botany,1956,4(2):167-174.
[13]Boswell M T,Ord J K,Patil G P.Chance Mechanisms Underlying Univariate Distributions[M].Burtonsville,Mal:International Co-operative Publishing House,1979.
[14]Daming H,Shiyomi M,Jun C,etal.Spatial heterogeneity of vegetation in Inner Mongolia,China[J].Grassland Science,2004,50(4):319-328.
[15]塩見正衛,安田泰輔.草地の新しい植生調査法(1):ベータ二項分布を利用した植生調査[J].畜産の研究,2003,57(1):171-175.
[16]Pielou E C.Mathematical Ecology[M].New York:John Wiley & Sons,1977:224-266.
[17]Shiyomi M,Takahashi S,Yoshimura T,etal.Spatial heterogeneity in a grassland community:Use of power law[J].Ecological Research,2001,16(3):487-495.
[18]陳俊,堀良通,塩見正衛,等.半自然草地および森林化した長期放任草地の群集構造[J].日本草地学会誌,2005,51(2):143-151.
[19]塩見正衛,安田泰輔.草地の新しい植生調査法(2):べき乗則による解析[J].畜産の研究,2003,57(2):303-306.
[20]陳俊,塩見正衛.べき乗則による反復植生データの空間的不均一性解析[J].日本草地学会誌,2008,53(4):282-288.
[21]Chen J,Yamamura Y,Hori Y,etal.Small-scale species richness and its spatial variation in an alpine meadow on the Qinghai-Tibet Plateau[J].Ecological Research,2008,23(4):657-663.
[22]安田泰輔,塩見正衛,石川涼,等.ランダム分布する種と集中する分布種[J].日本草地学会誌,2003,49(3):271-274.
