长芒草不同季节碳氮磷生态化学计量特征
2011-04-25牛得草董晓玉
牛得草,董晓玉,傅 华
(兰州大学草地农业科技学院 农业部草地农业生态系统学重点开放实验室,甘肃 兰州 730020)
生态化学计量学是结合了生物学、化学和物理学等基本原理,研究生物系统能量平衡和多重化学元素[主要是碳(C)、氮(N)、磷(P)]平衡的科学[1-3],它通过元素之间的计量比值把不同尺度、不同生物群系和不同研究领域的生态生物学特征有机地统一起来,成为新世纪生态学研究的有力工具[2,4-8]。生态化学计量学有两个重要的基础理论,分别为内稳性理论和生长速率理论。生物的内稳性是生态化学计量学存在的前提[4],该理论认为一切生物有机体具有在变化的环境中保持其体内化学组成相对恒定的能力[9]。另外,生长速率理论是指生长速率较高的生物具有较低的C∶N、C∶P和N∶P。这是由于生物快速生长需要合成大量的蛋白质,核糖体作为蛋白质合成的工厂,其核糖体RNA是生物体内P的主要贮藏库,因此,生长速率高的生物N、P含量也相对较高[10]。生长速率理论将细胞内化学物质的分配与个体生物学特征结合起来,为诸多生态学过程(如生态位理论、群落稳定性)的研究提供了新思路[11-13]。生物内稳性以及生长速率理论在浮游动物和微生物方面已有很多研究[4,14-15],而在高等植物方面报道较少[13,16-18]。另外,相比于低等的浮游动物和微生物,高等植物由于其存在细胞壁和行贮藏功能的液泡等亚细胞结构,以及高等植物在C固定和N、P吸收途径的分离,使得高等植物体内元素内稳性和生态化学计量特征的研究显得更加复杂,目前仍在广泛讨论之中[19-21]。
随着季节的变化,植物生长各阶段表现出不同的特征,例如,春季待植物返青后随着气温的回升,植物生长迅速,植物体生理代谢旺盛;秋季,植物生长停滞,植物体代谢减弱。植物的整个生长过程,需要光合作用所固定的碳来构建其有机体结构,以及合成的碳水化合物供植物代谢的能量需求,另外植物还需要不时地从土壤中吸收氮磷来合成蛋白质、核酸等生长必需物质。因此,伴随各生育阶段生长模式的改变,高等植物可能会不断地调整体内营养物质的分配。最终,高等植物在不同生长发育阶段体内元素是否具有内稳性,以及植物生长速率和元素含量及比率间的关系如何,成为了该研究关注的焦点[22]。本研究选取黄土高原地区长芒草(Stipabungeana)+冷蒿(Artemisiafrigida)典型草原主要建群种长芒草为研究对象,试图探讨高等植物不同发育阶段C、N、P元素含量及其计量关系的内稳性以及与生长速率的关系,另外对家畜采食对植物内稳性的影响进行初步探讨。
1 材料与方法
1.1研究地点概况 研究区位于甘肃省榆中县兰州大学半干旱气候与环境观测站及周围区域,地理坐标为35°57′ N,104°09′ E,海拔为1 965.8 m。该区属大陆性半干旱气候,年平均气温6.7 ℃,年降水量382 mm,年蒸发量1 343 mm,无霜期90~140 d,年日照时数约2 600 h。地貌为黄土高原残塬梁峁沟壑,土壤主要为灰钙土。植被类型为半干旱典型草原,塬面草地植物主要有长芒草、赖草(Leymussecalinus)、铁杆蒿(A.gmelinii)、冷蒿等[23]。
1.2研究方法
1.2.1样地设置和试验方法 在半干旱气候与环境观测站及周围,选择坡向和地势平缓一致的地段,设置2块面积各为6 hm2的围封地(FG,fenced grassland)和自由放牧地(GG,grazed grassland)样区,每样区内设置3块面积为50 m×50 m 样地。草地为农田退耕20多年,现已恢复到天然草地,主要优势种为长芒草。围封草地于2005年开始围封;自由放牧草地放牧家畜为绵羊,家畜白天放牧,夜晚入圈管理。
于2007年的3、5、7、9月中旬,在每个样地内设置6个1 m×1 m样方,每个处理(样区)共计18个样方(即重复),分别齐地面剪取长芒草地上组织,105 ℃下杀青15 min,65 ℃烘干至质量不变,测定其生物量。并按以下公式计算长芒草地上植物组织的生长速率[μ,g/(g·m2·月)][4]。
μ=[ln (B2/B1)]/dt。
式中,B2和B1分别为时间2和时间1测定的单位面积样方中长芒草地上生物量,dt是两次测定时间间隔(用月表示)。
将烘干的长芒草植物样品,粉碎过直径0.25 mm筛后,装入棕色瓶中,待分析用。采用重铬酸钾(K2Cr2O7)容量法测定植物组织有机碳含量[24],植物样品待凯氏法消化后,用流动注射分析仪(FIAstar 5000,瑞典FOSS公司)测氮含量;钼锑抗比色法测磷含量[25]。
1.2.2数据分析 所有数据均用Microsoft Excel 2003录入作图。用SPSS 16.0对数据进行统计分析,用独立样本T检验分析处理(围封地和自由放牧地)间的差异,用单因素方差分析比较月份之间的差异。
2 结果
2.1长芒草地上生物量及生长速率季节动态 长芒草地上生物量不同生长阶段表现出很大差异(图1)。围封草地长芒草地上生物量从3月到7月逐渐增大,在7月达到最大值,为80.44 g/m2,与7月相比,9月地上生物量略有下降,但差异不显著(P>0.05)。放牧样地长芒草地上生物量3月到9月的变化趋势与围封样地一致,但各测定月生物量都显著低于围封样地(P<0.05)。
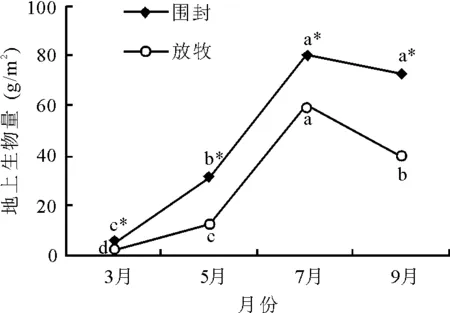
图1 围封与放牧样地长芒草地上生物量季节动态
围封样地长芒草在不同季节生长速率不同(图2),3-5月生长速率最大,为0.96 g/(g·m2·月),5-7月显著降低,为0.48 g/(g·m2·月),7-9月生长速率表现为负值,但经统计分析,与0 g/(g·m2·月)差异不显著。

图2 围封样地长芒草不同季节的生长速率
2.2长芒草C、N、P含量季节变化 随着生育期的延长,围封样地长芒草C含量从3月到5月显著升高(P<0.05),达486.02 mg/g,5月到9月稍有降低,且差异不显著(P>0.05);N含量从3月到9月逐月显著降低(P<0.05),3月最高,为28.80 mg/g;P含量变化规律与N含量相似,3月最高,为1.49 mg/g,7月和9月最低,为0.68~0.69 mg/g(图3)。放牧样地长芒草各月间C、N含量变化规律与围封样地一致,而P含量呈先显著降低到9月又升高的趋势,3月最高,为1.73 mg/g,9月次之,为0.91 mg/g(图3)。

图3 围封与放牧样地长芒草碳、氮、磷元素含量季节动态
围封样地长芒草5月的C含量显著高于放牧样地的(P<0.05),其他月份差异不显著(P>0.05)。长芒草的N含量各测定月均表现为放牧样地显著高于围封样地。除5月外,长芒草P含量放牧样地均显著高于围封样地。
2.3长芒草C、N、P计量特征的季节变化 随着植物生育期的延长,围封与放牧样地长芒草的C∶N逐渐增加,且各月围封样地均显著大于放牧样地(P<0.05)。围封样地长芒草的C∶P不同月份间表现出与C∶N同样的规律性,而放牧样地3-7月逐渐增加,7月达到最大值,然后降低;各月围封样地长芒草的C∶P均显著大于放牧样地(P<0.05)(表1)。 围封样地长芒草N∶P表现出先增大后减小的趋势,5月显著高于其他月份。3月长芒草的N∶P围封与放牧样地无显著差异,5和7月放牧地显著高于围封样地,9月围封样地显著高于放牧样地(表1)。
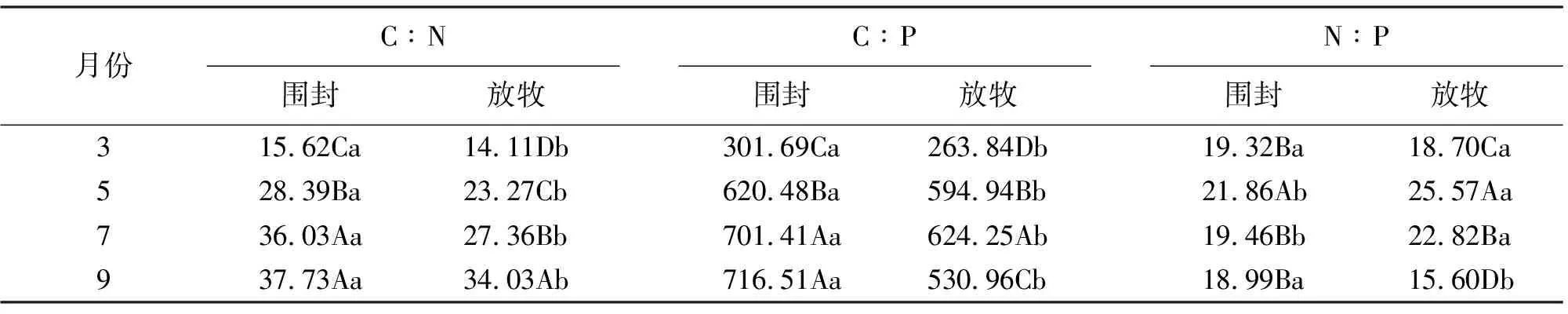
表1 长芒草碳、氮、磷计量特征
2.4长芒草C、N、P含量及其计量比值的季节间变化 从表2可以看出,围封样地内,不同生育时期长芒草C含量和N∶P的变异系数较小,分别为3.65%和6.60%,N、P含量及C∶N和C∶P的变异系数都大于30%。放牧样地长芒草C含量和N∶P变异也较小,但N∶P变异明显大于围封草地。
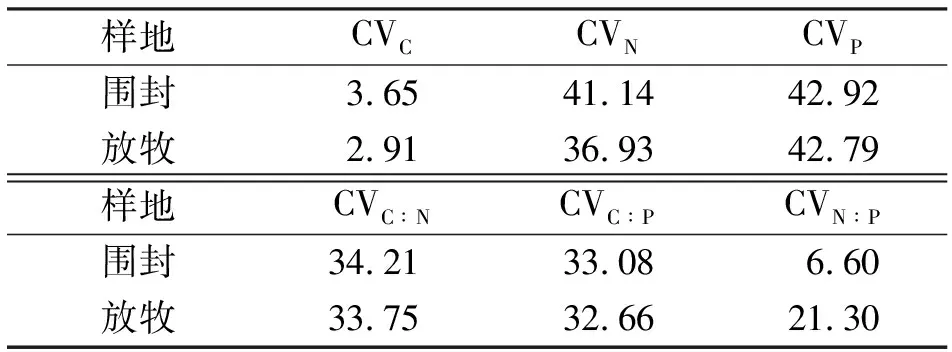
表2 长芒草碳、氮、磷元素含量和元素计量比值的季节间变异系数(CV) %
3 讨论
3.1长芒草C、N、P含量及其生态化学计量学特征 组成植物体的物质可分为3类:结构性物质、功能性物质和贮藏性物质。植物在不同的发育阶段,这3类物质在体内分配比例不尽相同。对于同一植物种来说,其结构性物质含量相对较稳定,不受环境、发育时间的影响;但功能性物质和贮藏性物质则变化较大[4,26]。生长初期,植物体功能性物质会优先向植物幼嫩组织转移,在生殖生长阶段贮藏性物质会向种子转移,繁殖对策上以获得更多的后代。对于多年生草本植物,秋后贮藏性物质会向根部转移,以保证植物越冬芽的存活及来年新生芽的快速生长[27]。本研究中,3-5月,长芒草处于营养生长阶段,生长初期植物体内物质主要集中于其形态建成,生长速率较快,细胞分裂迅速,此阶段植物体内功能性物质比例较大,因此,N、P含量3月较高。随着地上生物量的积累,植物光合叶面积增加,同化能力增强,机械组织进一步发展,植物体内结构性物质所占比例逐渐增大,5月长芒草C含量达最大值。同时,由于植物营养吸收和C的固定途径不同,营养吸收速率往往小于植物的C固定速率[28]。另外,植物体内营养元素可能受稀释效应的影响,长芒草N、P含量随时间的推移逐渐减少[29]。5-7月,环境温度升高,长芒草代谢旺盛,营养吸收和光合作用增强,此时生物量急剧增加,受稀释效应的影响,长芒草N、P含量仍表现为下降的趋势。7-9月,长芒草生长缓慢,处于生殖生长阶段,但由于繁殖对策的不同,多年生植物长芒草可能是侧重于无性繁殖,生殖枝较少,此时,植物体贮藏性物质向果实转移不多,另外,据观察长芒草的枯黄期在11月,这样贮藏物质向根部的转移可能在9月以后,所以长芒草7-9月C、N、P含量变化不大。
植物营养吸收和C的同化是不同途径,一般情况下,C不会成为植物生长的限制因子,而且C在植物体内含量很高,变异较小,因此,N和P的变化是影响C∶N和C∶P的主要因素[30]。3-7月由于长芒草生长迅速,N、P吸收与C的固定速率不成比例,其C∶N和C∶P显著增大,7-9月,由于长芒草体内贮藏性物质向果实转移不多,因此长芒草叶C∶N和C∶P变化不大。长芒草N∶P变化不明显,但在生长迅速的5月,其值显著增加。生长速率假说认为生长速率较高的生物具有较低的C∶N、C∶P和N∶P[10,31],而本研究结果与之矛盾,原因可能是生长速率理论的提出是基于浮游动物和细菌的研究,而高等植物与浮游动物、微生物的生长机理和物质组成有很大不同,且获得营养的方式也不同,使之生长速率与C∶N∶P之间的关系的研究显得更为复杂。低等的浮游动物和细菌与高等植物相比,并不具有细胞壁[10],此类生物体内结构性C占的比例较低。而对于高等植物,其细胞壁的主要成分为含C较多的果胶和纤维素,细胞壁的存在使得植物体结构性C占据了主要地位。另外,高等植物细胞的高度分化,各细胞在营养代谢与功能行使方面,既相互联系,又相互独立,从而使生长速率理论在高等植物方面的应用显得更加复杂[28]。
生物体都是由元素构成的,C∶N∶P不仅决定了生物体的关键特征,同时也决定了生物体对资源数量和种类的需求[5,32]。在不同的生育阶段,长芒草叶片中N∶P变异系数较小,表现相对稳定,而C∶N和C∶P变异较大。因此,讨论植物的内稳性需要具体到固定的指标。初步的研究表明长芒草在不同生育阶段其N∶P相对稳定,可以作为长芒草不受生育期影响的计量特征参与生态系统问题的分析。
3.2放牧对长芒草C、N、P含量及其生态化学计量学特征的影响 放牧地长芒草的生物量小于围封地,原因是生物量测定时部分生物量被家畜采食。不过,适度放牧能促进植物再生,使植物发生补偿性生长[33],导致植物枝条年龄结构趋于幼小化。放牧样地各季节长芒草C、N含量的变化规律与围封样地相似,但是7-9月放牧草地长芒草P含量较7月有所提高,其原因可能是植物此时处于放牧刺激后的再生长阶段。另外,放牧地长芒草叶片N、P含量均高于围封地的原因可能是放牧刺激了长芒草的再生生长,其植物新长出的幼嫩部分N、P含量相对较高,以至于C∶N和C∶P小于围封草地。5和7月放牧草地N∶P高于围封草地,可能是由于家畜排泄物引起土壤速效N增多,植物获得相对较多的氮素所致。另外,也可能是家畜采食不仅刺激牧草生长,还促进地上N、P向幼嫩器官重新分配,N的转移量大于P。
从各项指标的变异系数上可以看出,放牧草地长芒草C∶N、C∶P和N∶P变化较大。分析原因,可能是受家畜粪便干扰,或者家畜不均匀采食以及N、P的转移速率不一致引起。另外,放牧与围封草地长芒草碳含量比较差异不显著,表明植物体C含量相对稳定,同时体现了高等植物由于细胞壁的存在使得结构性C对总组织C含量的主导作用。
[1]Elser J J,Sterner R W,Gorokhova E,etal.Biological stoichiometry from genes to ecosystems[J].Ecology Letters,2000(3):540-550.
[2]曾德慧,陈广生.生态化学计量学:复杂生命系统奥秘的探索[J].植物生态学报,2005,29(6):1007-1019.
[3]贺金生,韩兴国.生态化学计量学:探索从个体到生态系统的统一化理论[J].植物生态学报,2010,34(1):2-6.
[4]Sterner R W,Elser J J.Ecological Stoichiometry:The Biology of Elements from Molecules to the Biosphere[M].Princeton:Princeton University Press,2002:1-50.
[5]Michaels A F.The ratios of life[J].Science,2003,300:906-907.
[6]曲浩,赵学勇,赵哈林,等.陆地生态系统凋落物分解研究进展[J].草业科学,2010,27(8):44-51.
[7]阎恩荣,王希华,郭明,等.浙江天童常绿阔叶林、常绿针叶林与落叶阔叶林的C∶N∶P化学计量特征[J].植物生态学报,2010,34(1):48-57.
[8]杨阔,黄建辉,董丹,等.青藏高原草地植物群落冠层叶片氮磷化学计量学分析[J].植物生态学报,2010,34(1):17-22.
[9]Kooijman S A L M.The stoichiometry of animal energetics[J].Journal of Theoretical Biology,1995,177:139-149.
[10]Elser J J,Acharya K,Kyle M,etal.Growth rate-stoichiometry couplings in diverse biota[J].Ecology Letters,2003,6:936-943.
[11]Sterner R W,Hessen D O.Algal nutrient limitation and the nutrition of aquatic herbivores[J].Annual Review of Ecology and Systematics,1994,25:1-29.
[12]Moen R A,Pastor J,Cohen Y.Antler growth and extinction of Irish elk[J].Evolutionary Ecology Research,1999,1:235-249.
[13]Yu Q,Chen Q,Elser J J,etal.Linking stoichiometric homeostasis with ecosystem structure,functioning,and stability[J].Ecology Letters,2010,13:1390-1399.
[14]Sterner R W,George N B.Carbon,nitrogen,and phosphorus stoichiometry of cyprinid fishes[J].Ecology,2000,81:127-140.
[15]Goldman J C,Caron D A,Dennett M R.Regulations of gross growth efficiency and ammonium regeneration in bacteria by substrate C∶N ratio[J].Limnology and Oceanography,1987,32:1239-1252.
[16]Zhang L X,Bai Y F,Han X G.Differential responses of N∶P stoiehiometry ofLeymuschinensisandCarexkorshinskyito N additions in a steppe ecosystem in Nei Mongol[J].Acta Botanica Sinica,2004,46:259-270.
[17]Cernusak L A,Winter K,Turner B L.Leaf nitrogen to phosphorus ratios of tropical trees:experimental assessment of physiological and environmental controls[J].New Phytologist,2009,185:770-779.
[18]Matzek V,Vitousek P M.N∶P stoichiometry and protein:RNA ratios in vascular plants:an evaluation of the growth-rate hypothesis[J].Ecology Letters,2009,12:765-771.
[19]Güsewell S.N∶P ratios in terrestrial plants:variation and functional significance[J].New Phytologist,2004,164:243-266.
[21]陈伏生,胡小飞,葛刚.城市地被植物动脉叶片氮磷化学计量比和养分在吸收效率[J].草业学报,2007,16(8):47-54.
[22]吴统贵,吴明,刘丽,等.杭州湾滨海湿地3种草本植物叶片N、P化学计量学的季节变化[J].植物生态学报,2010,34(1):23-28.
[23]Li X D,Fu H,Li X D,etal.Effects of land-use regimes on carbon sequestration in the Loess Plateau,Northern China[J].New Zealand Journal of Agricultural Research,2008,51:45-52.
[24]董鸣.陆地生物群落调查观测与分析[M].北京:中国标准出版社,1997:152-153.
[25]鲍士旦.土壤农化分析[M].第三版.北京:中国农业出版社,2000:268-270.
[26]Kerkhoff A J,Fagan W F,Elser J J,etal.Phylogenetic and growth form variation in the scaling of nitrogen and phosphorus in the seed plants[J].American Naturalist,2006,168:103-122.
[28]Berman-Frank I,Dubinsky Z.Balanced growth in aquatic plants:Myth or reality[J].BioScience,1999,49:29-37.
[29]Sardans J,Penuelas J.Drought changes nutrient sources,content and stoichiometry in the bryophyteHypnumcupressiformeHedw.growing in a Mediterranean forest[J].Journal of Bryology,2008,30:59-65.
[30]Hedin L O.Global organization of terrestrial plant-nutrient interactions[J].Proceedings of the National Academy of Sciences of the United States of America,2004,101:10849-10850.
[31]Makino W,Cotner J B,Sterner R W,etal.Are bacteria more like animals than plants? Growth rate and resource dependence of bacterial C∶N∶P stoichiometry[J].Functional Ecology,2003,17:121-130.
[32]Schimel D S.All life is chemical[J].BioScience,2003,53:521-524.
[33]刘王锁,谢应忠,代红军,等.荒漠草原不同放牧强度下长芒草种群补偿性生长的研究[J].农业科学研究,2008,29(2):50-53.
