预防围产期奶牛酮病的营养策略
2011-03-14乔国华李锦华周学辉王春梅
乔国华 李锦华 周学辉 杨 晓 张 茜 王春梅
(中国农业科学院兰州畜牧与兽药研究所,草业饲料研究室,兰州 730050)
围产期是奶牛饲养管理的关键时期,采用合理而科学的营养调控方法可以很大程度上避免常见代谢疾病(酮病、肝脏水肿、产后瘫痪等)的发病率。围产期奶牛饲喂管理不当会影响奶牛整个泌乳期的生产性能,加之代谢疾病的治疗费用,给奶牛养殖业带来了很大的经济损失。一些国家在过渡期使用莫能菌素来预防奶牛酮病的发生,取得了较好的效果。但是,鉴于饲料安全等方面的考虑,中国禁止饲喂莫能菌素。一些奶业发达国家已经将过渡期列为奶牛饲养的关键时期,在围产期奶牛酮病预防方面,通过饲粮的配制和饲料添加剂的使用获得了较大的成功。本文旨在综述过渡期奶牛酮病预防的营养管理策略,为中国奶牛健康养殖提供必要的理论基础和实践参考。
1 围产期奶牛脂肪代谢特点
在泌乳高峰期的奶牛处于能量负平衡状态,干物质采食量不足,奶牛机体肾上腺素和去甲肾上腺素水平升高,开始进行较大程度上的脂肪动员(fatmobilization),脂肪组织的甘油三酯转化为非酯化脂肪酸(NEFA),血液中的 NEFA浓度升高。据 Pullen等[1]和 Reyno ld等[2]报道,干物质采食量和 NEFA浓度通常呈现负相关的关系。但是,肝脏并没有完全承载 NEFA的能力,因其不能将 NEFA通过异化代谢的方式产生能量,或者经肝脏排出进入血液,而是在肝细胞线粒体内进行不完全氧化,生成酮体(β-羟丁酸,BHBA)进入血液循环。Emery等[3]报道当机体进行脂肪动员时,来自于脂肪组织的 NEFA大部分是以甘油三酯的方式在肝脏微粒体内储存,小部分直接进入乳房用于乳脂肪的合成(图 1)。
通常奶牛进入围产期的产后几周阶段,在肝脏内都要储存大量的甘油三酯。这时,过多的甘油三酯对肝脏的其他合成过程有很大的抑制作用。如:Piepenbrink等[4]的报道,肝脏利用丙酸合成葡萄糖的能力被削弱;Strang等[5]的报道,肝脏利用氨态氮合成尿素的能力也会受到抑制;Zhu等[6]报道,处在产后第 2天的奶牛,肝脏甘油三酯浓度升高,四周组织的氨态氮浓度提高了 1倍。Overton等[7]通过体外培养表明,氨态氮显著抑制肝脏利用丙酸合成葡萄糖的能力。因此,肝脏内积累甘油三酯势必会抑制肝脏利用丙酸合成葡萄糖的过程。很有可能,是由于氨态氮可以调控肝脏合成葡萄糖的过程。但是,氨态氮调控肝脏代谢过程的机理有待于进一步试验来确定。
2 围产期奶牛的营养调控策略
2.1 满足葡萄糖需要来降低 NEFA
奶牛产后,葡萄糖需要提高到了产前的 2倍左右。为了适应奶牛产后对葡萄糖的需要,一些关于碳水化合物营养的试验主要集中在了非纤维性碳水化合物(NFC)的水平上。有学者认为,在干奶期提高奶牛饲粮 NFC水平,可促进瘤胃发酵产生丙酸,使瘤胃微生物尽早适应产犊后的高NFC饲粮[8]。
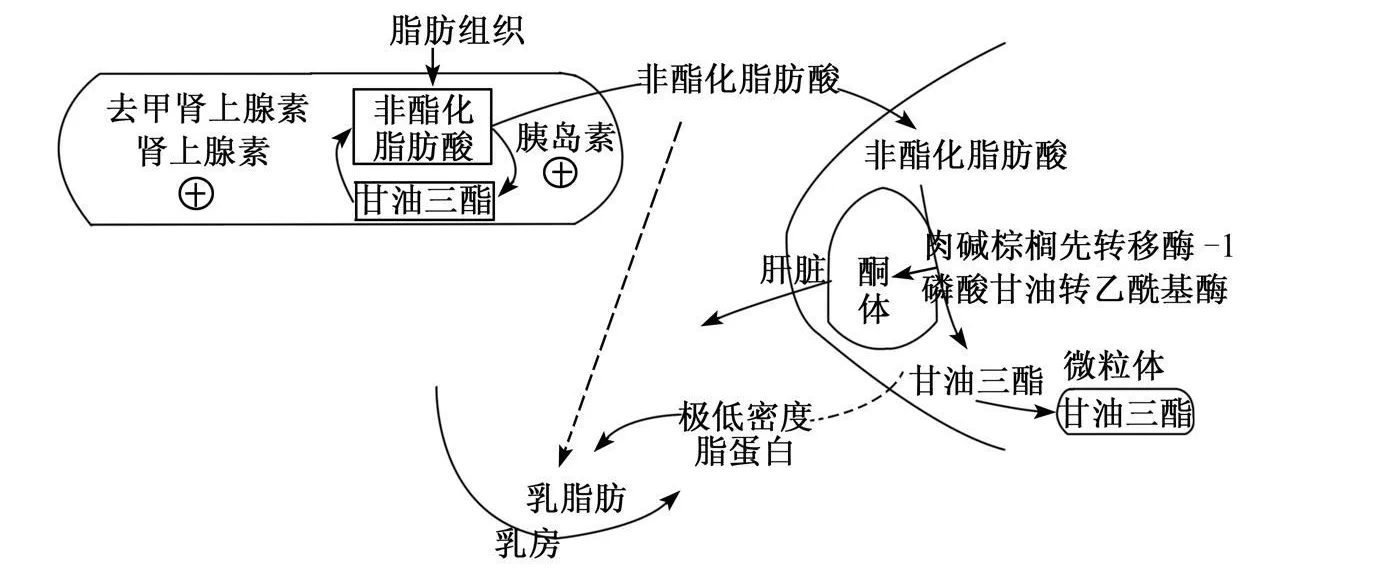
图 1 过渡期奶牛肝脏脂肪代谢通路Fig.1 Pathway of fatmetabolism in the liver of dairy cow s during transition period
由表 1中的结果可知,在干奶期奶牛饲粮中添加较高水平的 NFC可以提高干奶期奶牛产前的干物质采食量。在饲粮 NFC水平对产前和产后的瘤胃发酵影响方面也有了一些报道,Dann等[15]报道,用蒸汽压片玉米替代粉碎的玉米,以加强淀粉在瘤胃内的发酵,不仅提高了产前干物质采食量,而且提高了产后的泌乳性能和血浆胰岛素的水平,这个作用一直持续到了泌乳中期。瘤胃可发酵碳水化合物的提高也降低了产前血浆 NEFA的水平。Ordway等[16]报道,在过渡期奶牛饲粮中直接添加蔗糖 2.7%(干物质基础),其并没有改变产前血浆葡萄糖的水平,也没有改变产后血浆非酯化脂肪酸的水平。Penner等[17]报道,用蔗糖替代过渡期奶牛饲粮 8.4%的 NFC并没有改变瘤胃液丙酸的浓度,对血浆 NEFA和甘油三酯均没有影响,只是提高了干物质采食量。

表 1 饲粮NFC水平对围产期分娩前奶牛代谢和生产性能的影响Table 1 Effects of dietary NFC level onmetabolism and performance of dairy cow s in peripartum during perinatal period
Smith等[18]报道,用甜菜渣、大豆皮等加工副产品替代部分来源于粗饲料的纤维,试验中的处理为 2种不同 NFC水平的饲粮,高 NFC饲粮泌乳净能 1.59 M cal/kg(1 cal=4.18 J)、NFC 40%和淀粉 28%,高非粗饲料纤维饲粮泌乳净能1.54 M cal/kg、NFC 34%和淀粉 18%。结果表明,2种饲粮对奶牛产前和产后干物质采食量没有影响,对产后的生产性能也没有影响,对血液葡萄糖水平也没有影响。由于甜菜渣和大豆皮中的纤维成分在瘤胃内的降解程度较高,提高了瘤胃液内丙酸的浓度。虽然饲粮淀粉水平比较低,但是奶牛血糖浓度并没有降低。这项研究结果说明,产前的饲粮 NFC水平对血糖浓度没有影响,瘤胃内可发酵的碳水化合物的水平是影响血糖浓度的关键因素。
2.2 直接添加生糖前体物质
2.2.1 直接添加丙二醇
丙二醇是生成葡萄糖的前体物质,一直以来都被用来治疗奶牛的酮病。丙二醇通常在奶牛产后第 2~3天以丸剂的形式添加。据 Stokes等[19]和 Pickett等[20]报道,奶牛饲喂丙二醇后,血浆非酯化脂肪酸的浓度和 β-羟丁酸浓度都下降。
2.2.2 直接添加丙酸
丙酸是奶牛肝脏用以合成血糖的主要前体物质,在饲料中添加丙酸从理论上讲可以提高血浆葡萄糖的水平,丙酸通常以其钙盐的形式添加,或者是其他的离子盐。但是,Burhans等[21]报道,在奶牛饲粮中添加 300 g/d的丙酸钙并没有改变奶牛血浆非酯化脂肪酸的水平,也并没有提高奶牛的产奶量。Mandebvu等[22]报道,产后奶牛添加110 g/d丙酸钙虽然没有提高产奶量,但是降低了血浆非酯化脂肪酸和尿液中酮体的水平。添加丙酸钙为什么没有提高奶牛的产奶量?为什么没有改变血浆非酯化脂肪酸的水平?目前还不清楚。需要进一步的试验研究来确定。
2.3 围产期饲粮添加脂肪
Kronfeld[23]报道,在干奶期奶牛饲粮中添加脂肪可以降低血液非酯化脂肪酸的水平和酮体的水平(β-羟丁酸)。饲粮长链脂肪酸被直接吸收进入淋巴系统,并不会进入肝脏。这种脂肪可以为四周组织和乳腺提供能量。K ronfeld提出了一个假说,这种长链的脂肪酸可以降低机体的脂肪动员,降低非酯化脂肪酸的水平。但是,Douglas等[24]和 Bertics等[25]报道,添加长链脂肪酸并没有降低血液酮体和非酯化脂肪酸的水平,但为了防止奶牛酮病所引发的经济损失,这种添加剂在很多奶牛养殖厂得到了较广的应用。Grum等[26]报道,整个干奶期奶牛饲粮添加 6.7%脂肪降低了肝脏中甘油三酯的水平,但是,也降低了奶牛的干物质采食量。之后,Douglas等[24]经过分析后报道,肝脏甘油三酯的降低完全是由于干物质采食量降低了造成的。Doepel等[27]报道,在奶牛干奶期采用高能量饲粮(高 NFC水平)可以降低围产期奶牛的 NEFA的水平和肝脏甘油三酯的水平。从以上结果来看,是饲粮的 NFC水平而不是饲粮的脂肪水平在起作用。
2.4 特定脂肪酸对 NEFA供给的影响
过去的研究主要集中在特定的脂肪酸对奶牛代谢的作用上。在过渡期奶牛上主要是 2个方面:1)反 -10,顺 -12共轭亚油酸 (CLA)是一种可以明显降低乳脂肪合成量的脂肪酸[28-29]。这种脂肪酸的添加会因其降低了乳脂肪的产出而节约能量,从而,缓解奶牛泌乳高峰期的能量负平衡。Giesy等[30]报道,从产后第 13~80天饲喂奶牛共轭亚油酸的钙盐,并没有改变第 14~28天的产乳性能 (产奶量,乳成分)。但是,第 35~80天的乳产量提高了,乳脂率和乳脂产量都降低了。在这 2个时间段内的能量平衡状况并没有得到改善。Bernal-Santos等[31]报道,给产后第 14~140天的奶牛饲喂共轭亚油酸的同分异构体混合物,乳脂率和乳脂肪的产量都显著降低了。然而,给奶牛饲喂过瘤胃保护的共轭亚油酸却提高了奶牛的产奶量,奶牛的能量平衡状况并没有受到任何的影响。2)Selberg等[32]报道,饲喂给过渡期和泌乳高峰期奶牛反式十八碳烯酸降低了肝脏甘油三酯的浓度。相比而言,Bernal-Santos等[31]报道,共轭亚油酸(CLA)并不能降低奶牛肝脏甘油三酯的浓度。这 2种脂肪酸对肝脏脂类代谢有不同影响作用,需要进一步的试验来确定其具体的对肝脏脂类代谢的影响作用。
2.5 降低肝脏中 NEFA积累的营养策略
降低血液循环中的 NEFA水平,从而降低进入肝脏中的 NEFA的水平。这种营养策略旨在减少 NEFA在肝脏内合成甘油三酯。NEFA在肝脏内可以在过氧化物酶体内发生 β氧化,也可以合成甘油三酯以极低密度脂蛋白的形式进入血液。但是,Grummer[33]报道,后者的程度与单胃动物(猪和鸡)比较起来程度很有限。
2.5.1 胆碱
胆碱是一种类维生素,在哺乳动物代谢上有很广泛的代谢功能。是细胞膜磷酸卵磷脂的组成部分,是神经递质——乙酰胆碱的构成成分。也是甜菜碱甲基代谢的前体物质。在过渡期奶牛上,胆碱的研究主要集中在其对肝脏脂类代谢的影响上,胆碱是磷酸卵磷脂的主要成分,肝脏利用磷酸卵磷脂进一步合成极低密度脂蛋白(VLDL),然后分泌进入血液。Yao等[34]报道,胆碱缺乏的大鼠肝脏甘油三酯含量提高了 6倍,用体外研究方法研究胆碱缺乏大鼠的肝细胞,在胆碱和蛋氨酸浓度提高时,极低密度脂蛋白的合成与分泌也增加了。据 Piepenbrink等[4]和 Zahra等[36]报道,在过渡期奶牛上使用过瘤胃保护胆碱趋于降低脂肪酯化的产物,这个研究结果说明,极低密度脂蛋白的输出与胆碱的供给量呈正相关。Hartwell等[35]、Scheer等[37]、Piepenbrink等[4]和 Pinotti等[38]报道,在过渡期奶牛饲粮中添加过瘤胃保护胆碱可以提高乳和乳脂肪的产量,说明饲粮添加过瘤胃保护胆碱所引起的奶牛肝脏脂肪酸代谢变化可以提高泌乳早期奶牛生产性能。
2.5.2 油酸和亚油酸
油酸和亚油酸是生物机体不可或缺的营养素,亚油酸是合成二十二碳六烯酸和二十碳五烯酸的前体物质,是合成极低密度脂蛋白 B100的重要物质[39]。体外奶牛肝细胞培养研究表明,亚油酸对细胞甘油三酯积累的减少有作用[40]。Piepenbrink等[4]报道,产后奶牛肝组织切片与油酸和亚油酸混合物共培养发现,其可以降低脂肪酸的酯化作用。这些研究的结果可能有 2种解释,油酸可以作用在极低密度脂蛋白在肝脏内的合成与分泌上,也有可能作用在线粒体或者过氧化物酶体的 β氧化上,或者是二者皆有。需要进一步的试验研究来证实。
3 小 结
①奶牛产前饲粮高 NFC含量对肝脏酮体的生成没有作用。关键是要提高瘤胃的发酵过程中丙酸的生成量来提供机体所需能量。
②饲粮直接添加蔗糖并不能提高血糖的供给,不能改变机体能量负平衡的状况。
③奶牛产后饲粮添加生糖前体物质(丙二醇和丙酸盐)可以降低血液 NEFA和酮体的浓度。
④添加过瘤胃保护胆碱,是提高肝脏利用甘油三酯合成和分泌极低密度脂蛋白的有效途径。
[1] PULLEN D L,PALMQUIST D L,EMERY R S.Effecto f days of lactation and methioninehyd roxy analog on incorporation of p lasma fatty acids into p lasma trig lycerides[J].Journal of Dairy Science,1989,72(1):49-58.
[2] REYNOLDSC K,A IKMAN P C,LUPOLI B,et al.Sp lanchnic metabolism of dairy cow s during the transition from late gestation through early lactation[J].Journal of Dairy Science,2003,86(4):1201-1217.
[3] EMERY R S,LIESMAN JS,HERDT T H.Metabolism of long-chain fatty acids by rum inant liver[J].Journalo f Nutrition,1992,122(3):832-837.
[4] PIEPENBRINK M S,OVERON T R.Hepatic palm itatemetabolism of peripartrientdairy cow s as affected by nutrients supplied in vitro[J].Journal of Dairy Science,2003,86(Suppl.1):220.
[5] STRANG B D,BERTICS S J,GRUMMER R R,et al.Effecto f long-chain fatty acids on trig lyceride accumulation,gluconeogenesis,and ureagenesis in bovine hepatocytes[J].Journal o f Dairy Science,1998,81(3):728-739.
[6] ZHU L H,ARMENTANO L E,BREMMER D R,et al.Plasma concentration of urea,ammonia,g lutam ine around calving,and the relation of hepatic triglyceride,to plasma ammonia removal and blood acid-base balance[J].Journal of Dairy Science,2000,83(4):734-740.
[7] OVERTON T R,DRACKLEY J K,OTTEMANNABBAMONTE C J,etal.Substrate utilization for hepatic g luconeogenesis is altered by increased glucose dem and in rum inants[J].Journal of Animal Science,1999,77(7):1940-1951.
[8] RABELO E,REZENDE R L,BERTICS S J,et al.Effects of transition diets varying in dietary energy density on lactation performance and rum inal parameters of dairy cow s[J].Journal of Dairy Science,2003,86(3):916-925.
[9] GRUM D E,DRACKLEY JK,YOUNKER R S,et al.Nutrition during the dry period and hepatic lipid metabolism o f periparturient dairy cows[J].Journal of Dairy Science,1996,79(10):1850-1864.
[10] M INOR D J,TROWER S L,STRANG B D,et al.Effects o f nonfiber carbohyd rate and niacin on periparturientmetabolic status and lactation of dairy cow s[J].Journal of Dairy Science,1998,81(1):189-200.
[11] MASHEK D G,BEEDE D K.Peripartum responses of dairy cow s to partial substitution of corn silage w ith corn grain in diets fed during the late dry period[J].Journal of Dairy Science,2000,83(10):2310-2318.
[12] KEADY TW J,MAYNEC S,FITZPATRICK D A,et al.Effectof concentrate feed level in late gestation on subsequentm ilk yield,m ilk composition,and fertility o f dairy cows[J].Journal of Dairy Science,2001,84(6):1468-1479.
[13] HOLCOMB C S,VAN HORN H H,HEAD H H,et al.Effects of prepartum dry matter intake and forage percentage on postpartum performance of lactating dairy cow s[J].Journal o f Dairy Science,2002,84(9):2051-2058.
[14] DOEPEL L,LAPIERRE H,KENNELLY J J.Peripartum performance and m etabolism of dairy cow s in response to prepartum energy and p rotein intake[J].Journal of Dairy Science,2002,85(9):2315-2334.
[15] DANN H M,VARGA G A,PUTNAM D E.Im proving energy supp ly to late gestation and early postpartum dairy cow s[J].Journal o f Dairy Science,1999,82(8):1765-1778.
[16] ORDWAY R S,ISHLER V A,VARGA G A.Effects of sucrose supplementation on dry matter intake,m ilk yield,and blood metabolites of periparturient Ho lstein dairy cow s[J].Journal of Dairy Science,2002,85(4):879-888.
[17] PENNER G B,OBA M.Increasing dietary sugar concentration may imp rove dry matter intake,rum inal fermentation,and productivity of dairy cow s in the postpartum phase of the transition period[J].Journal of Dairy Science,2009,92(7):3341-3353.
[18] SM ITH K L,WALDRON M R,OVERTON T R,et al.Performance of dairy cows as affected by prepartum carbohydrate source and supp lementation with chrom ium throughout the periparturient period[J].Journal of Dairy Science,2002,85(Suppl.1):23.
[19] STOKESSR,GOFF JP.Evaluation of calcium propionate and propylene glyco l adm inistered into the esophagus at calving[J].Professional Animal Science,2001,17(3):115-122.
[20] PICKETTM M,CASSIDY T W,TOZER P R,et al.Effect of prepartum dietary carbohydrate source and m onensin on d ry matter intake,m ilk production and blood m etabolites of transition dairy cow s[J].Journal of Dairy Science,2003,86(Suppl.1):10.
[21] BURHANSW S,BELL A W,NADEAU R,et al.Factors associated with transition cow ketosis incidence in selected New England herds[J].Journal o f Dairy Science,2003,86(Suppl.1):247.
[22] MANDEBVU P,BALLARD C S,SN IFFEN C J,et al.Effect of feeding an energy supp lementprepartum and postpartum on m ilk yield and com position,and incidence o f ketosis in dairy cow s[J].Animal Feed Science and Technology,2003,105(8):81-93.
[23] KRONFELD D S.M ajor metabo lic determ inants of m ilk volume,mammary efficiency,and spontaneous ketosis in dairy cows[J].Journal of Dairy Science,1982,65(11):2204-2212.
[24] DOUGLAS G N,DRACKLEY J K,OVERTON T R,et al.Lipid metabolismand production by Holstein cow s fed control or high fatdiets at restricted or ad libitum intakes during the dry period[J].Journal of Dairy Science,1998,81(Suppl.1):295.
[25] BERTICSS J,GRUMMER R R.Effects of fat and methionine hydroxy analog on prevention or alleviation of fatty liver induced by feed restriction[J].Journal of Dairy Science,1999,82(12):2731-2736.
[26] GRUM D E,DRACKLEY JK,YOUNKER R S,et al.Nutrition during the dry period and hepatic lipid metabolism of periparturient dairy cows[J].Journal of Dairy Science,1996,79(10):1850-1864.
[27] DOEPEL L,LAPIERRE H,KENNELLY JJ.Peripartum performance and metabolism of dairy cows in response to p repartum energy and protein intake[J].Journal of Dairy Science,2002,85(9):2315-2334.
[28] BAUMGARD L H,SANGSTER JK,BAUMAN D E.M ilk fat synthesis in dairy cow s is progressively reduced by increasing supp lemental amounts of trans-10,cis-12 conjugated linoleic acid(CLA)[J].The Journalo f Nutrition,2001,131(10):1764-1769.
[29] BAUMAN D E,GRIINARIJM.Nutritional regulation ofm ilk fat synthesis[J].Annual Review of Nutrition,2003,23(2):203-227.
[30] GIESY JG,VISWANADHA S,HANSON TW,et al.Effects of calcium salts of conjugated linoleic acid(CLA)on estimated energy balance in Holstein cow s early in lactation[J].Journal of Dairy Science,1999,82(Supp l.1):74.
[31] BERNAL-SANTOSG,PERFIELD JW,BARBANO D M,etal.Production responses o f dairy cow s to dietary supplementation with conjugated linoleic acid(CLA)during the transition period and early lactation[J].Journal o f Dairy Science,2003,86(10):3218-3228.
[32] SELBERG K T,STAPLESCR,BAD INGA L.Production and metabolic responses to dietary conjugated linoleic acid(CLA)and trans-octadecenoic acid isomers in periparturient dairy cow s[J].Journal o f Dairy Science,2002,85(Supp l.1):19.
[33] GRUMMER R R.Etiology of lipid-related m etabo lic disorders in periparturient dairy cows[J].Journal of Dairy Science,1993,76(12):3882-3896.
[34] YAO Z M,VANCE D E.Reduction in VLDL,but not HDL,in plasma of rats deficient in choline[J].Biochem istry and Cell Biology-biochim ie et Bio logie Cellu laire,1990,68(2):552-558.
[35] HARTWELL JR,CECAVA J,DONKIN S S.Impacto f dietary rumen undegradable protein and rumenprotected choline on intake,peripartum liver triacylglyceride,plasma metabolites,and m ilk p roduction in transition dairy cows[J].Journal of Dairy Science,2000,83(12):2907-2917.
[36] ZAHRA L C,DUFFIELD T F,LESLIE K E,et al.Effects o f rumen p rotected choline and monensin on m ilk production andmetabolism of periparturientdairy cows[J].Journal of Dariy Science,2006,89(12):4808-4818.
[37] SCHEER W A,LUCY M C,KERLEY M S,et al.Effects of feeding soybeans and rumen-p rotected choline during late gestation and early lactation on performance of dairy cows[J].Journal of Dairy Science,2002,85(Suppl.1):276.
[38] PINOTTIL,BALDI A,POLITIS I,et al.Rumenprotected choline adm inistration to transition cows:effects on m ilk production and vitam in E status[J].Journalof Veterinarian Medicine,2003,50(1):18-21.
[39] WU X,SHANG A,JIANG H,et al.Demonstration of biphasic effects of docosahexaenoic acid on apolipoprotein B secretion in HepG 2 cells[J].A rteriosclerosis Thrombosis Vascular Biology,1997,17(11):3347-3355.
[40] MASHEK D G,BERTICSS J,GRUMMER R R.Metabolic fate of long-chain unsaturated fatty acids and their effects on palm itic acid metabolism and gluconeogenesis in bovine hepatocy tes[J].Journal of Dairy Science,2002,85(9):2283-2289.
