4个地理群体魁蚶(Scapharca broughtonii)的形态差异与判别分析
2011-01-12杨爱国刘志鸿周丽青
梁 超, 杨爱国, 刘志鸿, 周丽青, 吴 彪
(1. 农业部海洋渔业资源可持续利用重点开放实验室, 中国水产科学研究院 黄海水产研究所, 山东 青岛266071; 2. 上海海洋大学 水产与生命学院, 上海 200090)
4个地理群体魁蚶(Scapharca broughtonii)的形态差异与判别分析
梁 超1,2, 杨爱国1, 刘志鸿1, 周丽青1, 吴 彪1
(1. 农业部海洋渔业资源可持续利用重点开放实验室, 中国水产科学研究院 黄海水产研究所, 山东 青岛266071; 2. 上海海洋大学 水产与生命学院, 上海 200090)
运用多变量形态度量学分析方法对韩国统营、山东黄岛、山东蓬莱、江苏前三岛4个地理区域的魁蚶(Scapharca broughtoniiSchrenck)种群间的形态变异进行了研究。单因素方差分析(ANOVA)和Tukey检验表明, 魁蚶不同地理种群表现出显著形态变异(P<0.05)。聚类分析结果表明, 黄岛和前三岛两个种群形态最为接近, 首先聚为一类, 然后与韩国种群聚类, 再与蓬莱种群聚类; 主成分分析构建了3个主成分, 其贡献率PC1为33.366%,PC2为20.407%, PC3为15.422%, 3个主成分累计方差贡献率为69.195%; 建立了魁蚶4个种群的判别函数, 其判别准确率分别为韩国通营96.4%, 山东蓬莱100%,山东黄岛84.3%, 江苏前三岛95.6%。这些数据可为魁蚶不同地理种群的识别、种质资源的可持续利用及保护提供依据。
魁蚶Scapharca broughtonii; 形态变异; 方差分析; 主成分分析; 聚类分析; 判别分析
魁蚶(Scapharca broughtoniiSchrenck)隶属于软体动物门(Mollusca),半鳃纲(Lamellibranchia), 列齿目(Taxodonta), 蚶科(Arcidae), 俗称赤贝、血贝、大毛蛤, 是一种大型冷温性蚶类, 广泛分布于太平洋西部沿岸、日本北海道以南、朝鲜、菲律宾、俄罗斯东南部等地区。在中国主要分布在辽东半岛东南部、山东半岛北部和东部等海区[1]。
魁蚶成体个大体肥, 肉质鲜美, 经济价值很高,近年来由于市场和出口大量需求, 自然资源远远不能满足市场需求, 人工养殖得到大力发展。在魁蚶养殖生产实践中, 中国魁蚶在生长速度、成活率等生物学指标方面与韩国魁蚶有一定的差异, 韩国魁蚶生长速度较快, 成活率较高。养殖单位比较认可韩国的魁蚶苗种, 许多苗种生产单位每年从韩国引进魁蚶种贝进行苗种生产。目前魁蚶人工育苗、繁殖与生长和生理生化等方面的研究已有较多报道[2-7], 尚未见魁蚶地理种群间形态特征变异的报道。为进一步丰富魁蚶形态学方面的基础资料, 本文采用多变量形态度量学的方法对一个韩国魁蚶自然群体和 3个中国魁蚶自然群体进行地理种群间的形态变异研究,目的在于探讨魁蚶不同地理种群的形态特征变异特点、分化水平以及进行多变量分析的可行性, 建立魁蚶不同地理种群的判别方法, 为魁蚶不同地理种群的识别、种质资源的可持续利用及保护提供可靠依据。
1 材料与方法
1.1 材料
4个群体的魁蚶于2007年8月至10月份分别随机采自于山东蓬莱(SP)、山东黄岛(SHD)、江苏前三岛(JQ)、韩国统营(Korea)4个沿海海区, 均为自然群体(图1)。各群体数目分别为31、51、45、28。
1.2 形态指标及测定
根据魁蚶的外部形态, 测量各个体的形态指标。用游标卡尺(精度 0.01 mm)常规测量壳长(L)、壳宽(W)、壳高(H)、壳顶宽(w)、外韧带长(l)、背缘长(x)和放射肋宽y(左壳中部最宽放射肋的腹缘处宽度),然后吸干贝壳表面水分, 用电子天平(精度0.0001 g)称壳湿质量(z)。统计放射肋数量(R)。测量部位及参数见图1。

图1 魁蚶形态测定Fig. 1 Morphological measurements of S.broughtonii
1.3 数据处理
将各性状的度量值与其壳长(L)的回归剩余值作为个体大小的矫正值(壳湿质量经 ln转化), 以消除个体大小对形态特征的影响, 得到 7个比例性状和一个计数性状, 运用 SPSS11.5分析软件进行以下统计分析[8-9]:
单因素方差分析(ANOVA):进行单因素方差分析, 比较魁蚶不同地理群体间不同性状的差异。
聚类分析(CA):求出各个群体中 7个比例性状和一个计数性状平均值, 采用欧氏距离的最短距离平方法进行系统聚类分析, 绘出反映形态相似性的树形图。
主成分分析(PCA):获得多个形态指标的综合指标, 主成分贡献率和累计贡献率, 绘出所有样本的主成分散布图。
判别分析(DA):用逐步判别法进行分析, 采用对4个群体形态差异贡献较大的参数建立4个群体的判别函数。
2 结果
2.1 形态差异比较
4个地理群体魁蚶形态特征的描述性统计值见表1。4个地理种群魁蚶的度量性状矫正值和计数性状经单因子方差分析结果表明:韩国群体与蓬莱群体在H、S、w、l、y、R上有显著差异(P<0.05), 韩国群体与黄岛群体在w、y、z、R上有显著差异(P<0.05), 韩国群体与前三岛群体在W、w、l、x、y、z、R上有显著差异(P<0.05); 蓬莱群体与黄岛群体在H、W、W、l、x、y、z、R上有显著差异(P<0.05), 蓬莱群体与前三岛群体在H、w、x、z、R上有显著差异(P<0.05); 黄岛群体与前三岛群体在W、w、l、x、y上有显著差异(P<0.05)。Tukey分析结果见表2。
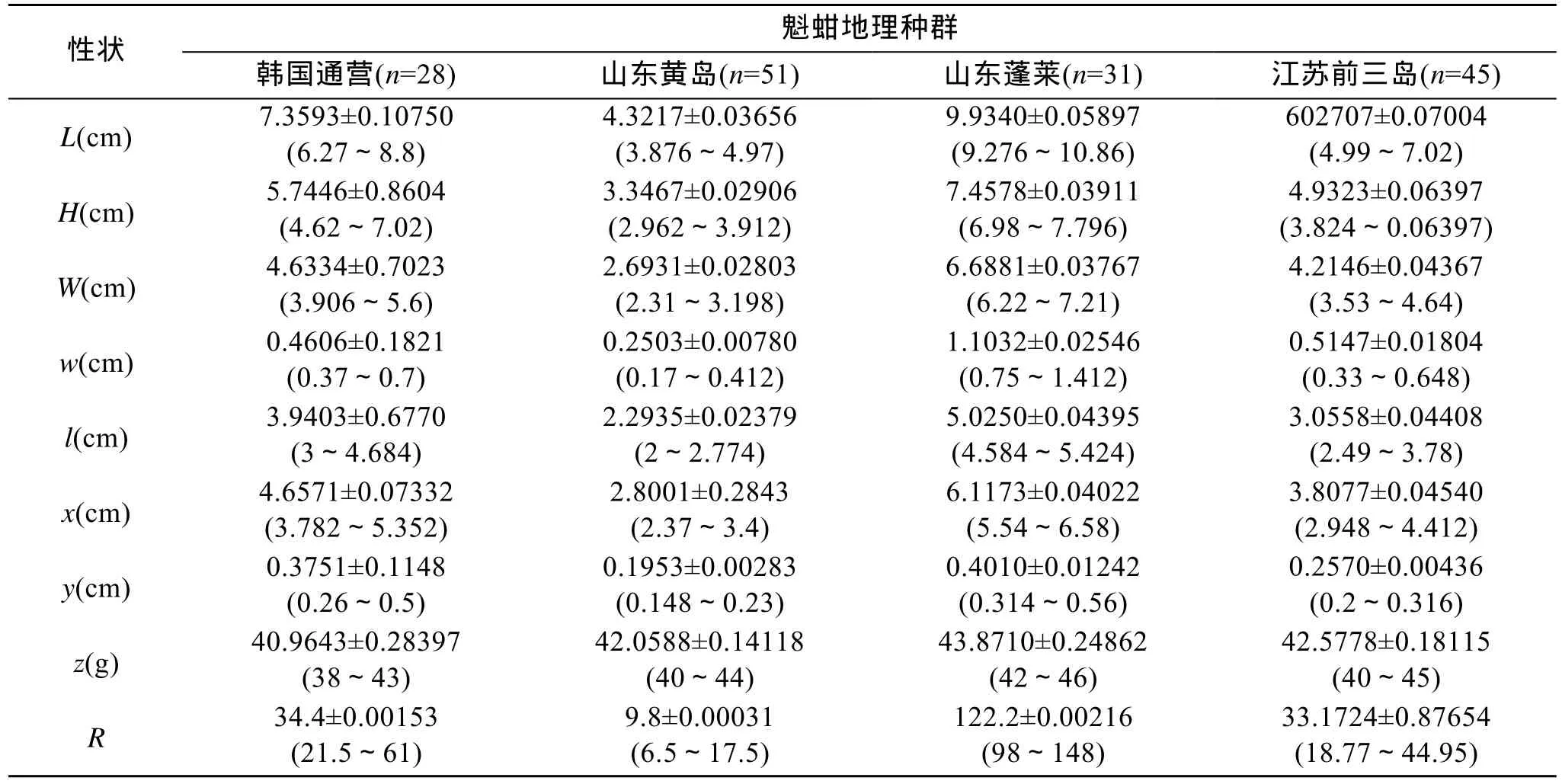
表1 不同地理种群魁蚶形态特征的描述性统计值Tab. 1 Descriptive statistics on the morphological characteristics of S.broughtonii from diferent geographical populations

表2 魁蚶不同地理种群8个性状的Tukey多重比较Tab. 2 Turkey multiple comparisons for five characters of different regional populations of S.broughtonii
2.2 主成分分析
通过对经标准化后的所有样本的 8个性状数据进行主成分分析, 得到了 PC1、PC2和PC3 三个主成分, 8个性状对3个主成分的负荷值和累计贡献率见表3。结果表明, 3个主成分解释形态变异的方差贡献率为:PC1为33.366%, PC2为20.407%, PC3为15.422%, 累计方差贡献率为 69.195%; 在第一主成分中, 形态学指标w/L、RR有较高的负荷值, 第二主成分中,l/L、x/L具有较高的负荷值, 第三主成分中,H/L、lnz/L具有较高的负荷值。4个魁蚶地理种群的主成分散布图见图2。通过不同群体的重叠区域可以看出, 韩国群体、黄岛群体在 PC2轴上重叠区域较多, 韩国通营群体与前三岛群体在PC3轴距离较远。前三岛群体与黄岛群体在PC1轴、PC2轴上各有部分重叠, 而在PC3轴上差别较大。
2.3 聚类分析
对4个群体的8个形态校正值进行聚类分析, 结果如图3。黄岛和前三岛两个种群形态最为接近, 首先聚为一类, 然后与韩国种群聚类, 再与蓬莱种群聚类。
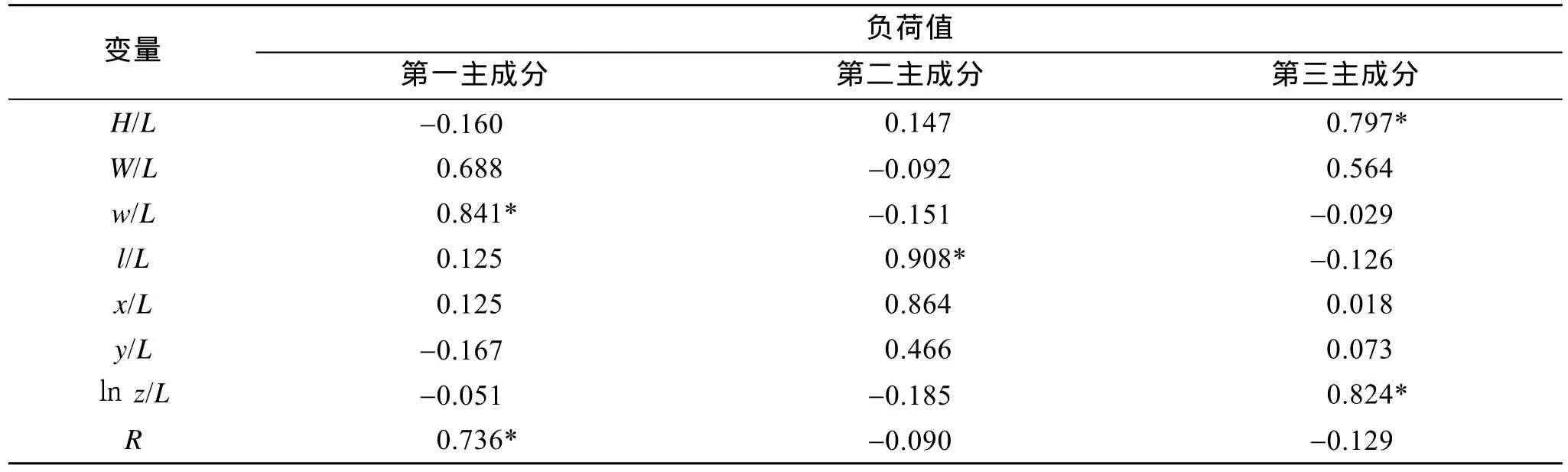
表3 魁蚶形态特征主成分的负荷值和贡献率Tab. 3 Contributive proportion and loading of principal components on the morphological characteristics of S.broughtonii
2.4 判别分析
通过逐步判别分析, 用筛选的 6个比例性状与一个计数性状的特征值建立了魁蚶 4个地理种群魁蚶的判别函数, 判别公式如下:


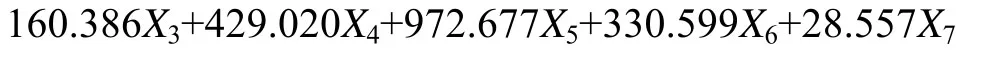
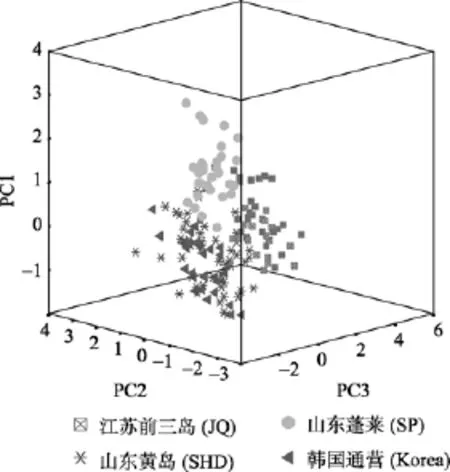
图2 魁蚶不同地理种群的3个主成分散布图Fig. 2 Scatter diagram for PC1, PC2 and PC3 of S.broughtonii from different geographical populations

图3 魁蚶不同地理种群的聚类分析图Fig. 3 Clustering dendrogram of S.broughtonni from different geographical populations
式中X1、X2、X3、X4、X5、X6、X7分别代表了SW/SL、UW/SL、OLL/SL、DBL/SL、RRW/SL、(lnSMW)/SL、RR。
将随机个体的6个比例性状值与一个计数性状值分别代入4个公式, 计算出4个函数值, 将函数值最大的判别函数所对应的种群名称作为该个体的地理种群名。按上述判别方法对所有的观测样本进行预测分类,判别结果表明总判别准确率为92.9%, 其中韩国通营、山东蓬莱、山东黄岛、江苏前三岛 4个种群的判别准确率分别为96.4%、100%、84.3%、95.6%。
3 讨论
3.1 多变量形态度量学分析方法在魁蚶形态差异判别上的应用
在贝类分类与鉴定上, 形态描述只是通过壳长、壳高、壳宽等少数几个形态数量性状和一些质量性状如贝壳颜色、纹理、形状等来进行简单归类。但对于一些形态相似的种类和种群, 这些方法不能达到很好的鉴别目的, 多变量形态度量学在贝类形态差异分析中已得到广泛应用[10-15]。本研究采用聚类分析、主成分分析和判别分析三种多变量形态学分析方法对 4个魁蚶不同地理种群进行研究, 分析了魁蚶的9个形态学参数的7个比例性状和一个计数性状, 在形态上较好地区分了这 4个地理种群的形态学差异。聚类分析是一种探索性的分析方法, 从样本数据出发, 将形态相近的种群进行初步归类, 并可以量化种群间的差异程度, 分析种群间的相似程度[16-17]; 主成分分析是将原始的变量信息进行重新组构, 将多个形态比例性状综合成可以反映原始资料大部分信息的少数几个因子, 从而得出不同种群的差异大小, 本文把4个魁蚶地理种群7个比例性状和一个计数性状综合成 3个能较好代表魁蚶各群体特点的因子, 来进行不同地理群体形态差异的比较得到了比较好的效果; 判别分析通过建立判别函数来进行种和种群的判别, 逐步判别是从已知的因子中选出必要、最佳组合的因子来建立判别函数[9], 通过逐步判别分析, 用筛选的 6个比例性状与一个计数性状的特征值建立了魁蚶 4个地理种群魁蚶的判别函数, 判别效果比较理想。
3.2 魁蚶不同地理种群形态差异比较分析
魁蚶 4个地理种群的单因素方差分析表明, 在进行比较的性状上, 4个种群不同的性状之间有显著差异。聚类分析结果与群体间的地理距离基本一致,黄岛和前三岛群体的形态差异较小, 韩国与黄岛、前三岛群体的形态差异较大, 蓬莱与黄岛群体、前三岛群体、韩国群体的形态差异最大。但是需要指出的是蓬莱群体与黄岛群体、前三岛群体之间的距离比韩国群体与黄岛群体、前三岛群体之间的距离要大,这与群体间的地理距离有出入, 么宗利[15]对青蛤(Cyclina sinensis)不同地理群体进行形态学分析时也有这样的问题, 并提出仅采用形态学指标分析并不能完全准确反映其地理分布特性, 要结合其他的生物学证据, 比如遗传学数据, 这样或许能够更好说明不同地理群体间的关系[18]。Laudien等[19]在对南非海岸的斧蛤(Donax serra)进行形态和遗传变异研究时提出, 形态学上的差异是由地理环境特性决定的, 而群体间的遗传相似性是由基因交流选择和进化习惯造成的。
本研究通过主成分分析, 得到了 3个能较好说明魁蚶种群形态特征的因子。根据PC1、PC2、PC3作出魁蚶不同种群的主成分散布图, PC1中w/L、R的负荷值较高, PC2中l/L、x/L的负荷值较高, PC3中H/L、(lnz)/L的负荷值较高。主成分散布图上的重叠区域说明了魁蚶不同种群形态特征的相似性,非重叠区域说明了魁蚶不同种群形态差异。通过不同群体的重叠区域可以看出, 韩国、黄岛群体在PC2轴上重叠区域较多, 韩国通营与前三岛群体在 PC3轴距离较远。前三岛与黄岛群体在PC1轴、PC2轴上各有部分重叠, 而在PC3轴上差别较大, 因为PC3受H/L、W/L、(lnz)/L影响较大, 而黄岛与前三岛群体在W、z有显著差异(P<0.05), 蓬莱群体独自成区,这在聚类分析中可以看到同样的结果, 蓬莱群体与其他 3个群体的距离最远。张永普[13]对泥蚶(Tegillarca granosa)不同地理种群、钱荣华[10]对中国五大湖三角帆蚌(Hyriopsi cumingii)的不同种群、冯建彬[14]对中国 4个海区不同群体文蛤(Meretrix meretrix)等所进行的主成分分析都较好地说明了不同群体间的形态差异。这种差异可能是由栖息环境不同造成的, 如水温、盐度、饵料等, 魁蚶为底栖动物, 其形状在一定程度上还与底质的理化性质有关。
从判别分析结果来看, 韩国、蓬莱、前三岛群体的判别率较高, 建立的判别函数可较好地反映各自群体的形态特征, 可以正确地区分 4个魁蚶不同地理种群。黄岛判别率较低, 可能是因为它与前三岛的形态差异较小, 导致判别准确率下降。
形态学表型差异建立在一定遗传基础之上并与生存环境密切相关[20-21]。形态特征是由遗传因子决定, 但是受到环境因子的影响, 地理屏障使同种的不同种群在一定程度上相互隔离, 受到不同栖息环境的影响, 如果环境因子差别较大并长期作用于种群, 就会导致不同地理种群在形态上产生较大的差异, 甚至在遗传上形成一定的差异。本研究表明, 韩国统营群体与中国魁蚶群体有较大差异, 这一现象值得我们思考:魁蚶地理种群之间的形态变异主要受栖息环境中哪些因子的影响, 相关环境因子与形态变异相关程度有多大, 这些差异又有多少是反映在遗传物质的变异上呢?导致韩国魁蚶和中国魁蚶在养殖过程中的差异是否是由遗传因子造成的呢?如果是, 能否利用这种遗传差异来改良魁蚶品种以提高经济性状, 这些问题需要继续开展生态学和分子遗传方面的研究, 并结合生产实践的验证才能给予解释。
[1]刘世禄, 杨爱国. 中国主要海产贝类健康养殖技术[M]. 北京:海洋出版社, 2005:176-177.
[2]王子臣, 张国范, 高勉, 等.温度和饵料对魁蚶性腺发育的影响[J]. 大连水产学院学报, 1987,8(2):1-9.
[3]杨爱国.魁蚶人工育苗的研究[J]. 海洋水产研究,1990, 32:31-35.
[4]唐启升, 邱显寅, 王俊, 等. 应用生态学报[J]. 山东近海魁蚶资源增殖研究, 应用生态学报, 1994, 5(4):396-402.
[5]邱显寅, 王俊, 郭学武, 等. 魁蚶中间育成实验研究[J]. 海洋水产研究, 1994, 15:87-95.
[6]王俊. 魁蚶食性及摄食季节变化初步研究[J]. 海洋水产研究, 1994, 15:65-69.
[7]殷邦忠, 腾瑜, 王家林, 等. 魁蚶低温保活方法研究[J]. 中国水产科学, 1994, 1(2):40-46.
[8]张尧庭, 方开秦. 多元统计分析引论[M]. 北京:科学出版社, 1982.
[9]余建英, 何旭宏. 数据统计分析与SPSS应用[M]. 北京:人民邮电出版社, 2003.
[10]钱荣华, 李家乐, 董志国, 等. 中国五大湖三角帆蚌形态差异分析[J]. 海洋与湖沼, 2003, 34(4):436-443.
[11]周晓农, 洪青标, 孙乐平, 等. 中国钉螺螺壳的聚类分析[J]. 动物学杂志, 1997, 3(25):4-7.
[12]魏开建, 熊邦喜, 赵小红, 等. 五种蚌的形态变异与判别分析[J]. 水产学报, 2003, 2(71):13-18.
[13]张永普, 林志华, 应雪萍. 不同地理群体泥蚶的形态差异与判别分析[J]. 水产学报, 2004, 28(3):339-342.
[14]冯建彬, 李家乐, 王美珍, 等.我国四海区不同群体文蛤形态差异与判别分析[J]. 浙江海洋学院学报(自然科学版), 2005, 24(4):318-323.
[15]么宗利, 王慧, 周凯, 等. 我国 5个青蛤地理群体形态差异分析[J]. 海洋水产研究, 2007, 28(2):63-70.
[16]蔡庆华. 武汉东湖浮游植物水华的多元分析[J]. 水生生物学报, 1990, 14(1):22-31.
[17]李勤生, 蔡庆华, 华俐, 等.东湖异养细菌群落的分类结构和聚类分析[J].水生生物学报, 1991, 15(3):242-253.
[18]Murta A G. Morphological variation of horse mackerel(Trachurus trachurus) in the Iberian and North African Atlantic:Implications for stock identification[J].ICES Journal of Marine Science, 2000, 57(4):1240-1248.
[19]Laudien J, Flint N S, Bank F H, et al.Genetic and morphological variation in four populations of the surf clamDonax seltafrom southern African sandy beaches[J]. Biochemical Systematics and Ecology,2003, 31(7):751-772.
[20]Salini J P, Milton D A, Rahman M J, et al.Allozyme and morphological variation throughout the geographic range of the tropical shad, hilsaTenualosa ilisha[J].Fisheries Research, 2004, 66(1):53-69.
[21]Susanna P, Jorma P. Morphological differentiation among local trout(Salmo trutta) populations[J]. Biological Journal of the Linnean Society, 2001, 72(2):231-239.
Morphological variations and discriminant analysis of different geographical populations ofScapharca broughtoniiSchrenck
LIANG Chao1,2, YANG Ai-guo1,LIU Zhi-hong1, ZHOU Li-qing1, WU Biao1
(1. Key Laboratory for Sustainable Utilization of Marine Fisheries Resources, Ministry of Agriculture, Yellow Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Qingdao 266071, China; 2. College of Fisheries and Life Science, Shanghai Ocean University, Shanghai 200090, China)
Apr., 18, 2010
Scapharca broughtonii; morphological variation; ANOVA; principal component analysis; cluster analysis; discriminant analysis
The multivariate morphometrics based on 9 morphological characters were used to investigate the morphological variations among the four different geographical populations ofS.brougghtoniifrom Kroea and China (Shandong and Jiangsu). ANOVA and Tukey test indicated that these four populations showed significant morphological variations (P<0.05). The result of cluster analysis showed thatS.broughtoniifrom Huangdao, Shandong and Qiansandao, Jiangsu are similar and clustered first, then clustered withS.broughtoniifrom Korea, and clustered withS.broughtoniifrom Penglai Shandong last. Three principal components were constructed in the principal component (PC) analysis by factor loading; the first principal component was mostly affected by umbo width and radial rib width; the second principal component was mostly affected by outer ligament length and dorsal brim length; and the third principal component was mostly affected by shell height and shell moisture weight. The contributory ratios of three principal components were 33.366%, 20.407%and 15.422%, respectively; and the cumulative contributory ratio was 69.195%. The discriminant functions of four populations were established and the dicriminant accuracy was 100% for theS.broughoniifrom Penglai Shandong, 96.4%, 84.3%, and 95.6% forS.broughtoniifrom Korea, Huangdao (Shandong), and Qiansandao(Jiangsu). The result of this research can be used in discrimination of different geographical populations and sustainable utilization and protection of the breed ofS.broughtonii.
S968.3 文献标识码:A 文章编号:1000-3096(2011)11-0108-06
2010-04-18;
2010-07-12
国家 863计划项目(2006AA10A408); 国家科技支撑计划项目(2006BAD01A13-6); 中央级公益性科研院所基本科研业务费项目(2007-qn-14); 国家科技基础条件平台建设项目(2006DKA30470-006)
梁超(1980-), 男, 硕士, 主要从事贝类遗传育种研究,E-mail:lcvshxq@163.com; 杨爱国, 通信作者, 电话:0532-85811982,E-mail:yangag@ysfri.ac.cn
张培新)
