六线鱼科鱼类特殊体色与繁殖特性
2011-01-12于道德张少春高凤翔胡发文
郭 文, 于道德, 潘 雷, 张少春, 高凤翔, 胡发文
(山东省海水养殖研究所, 山东 青岛 266002)
六线鱼科鱼类特殊体色与繁殖特性
Special coloartion changes and reproductive characteristics of greenlings (family Hexagrammidae)
郭 文, 于道德, 潘 雷, 张少春, 高凤翔, 胡发文
(山东省海水养殖研究所, 山东 青岛 266002)
六线鱼科(Hexagrammidae)隶属鲉形目(Scorpaeniformes), 六线鱼亚目(Hexagrammoidei)。六线鱼属(Hexagrammos)最大的特点就是身体具有多条侧线, 由于其中的斑头鱼仅仅具有一条侧线而被另外立新属, 斑头鱼属(agrammus), 但是更多的学者还是习惯于将斑头鱼归为六线鱼属。因此六线鱼科鱼类就是包括六线鱼属在内共5属12种(或6属12种)。六线鱼科的鱼类统称的英文名为 greenling,类似于石斑鱼的grouper以及鲆鲽类的flatfish。如果严格区分, 在文献中具有英文名称 greenling的六线鱼科鱼类仅仅有9种(表1)。中国分布的六线鱼科鱼类仅有4种, 分别为斑头鱼、大泷六线鱼、长线六线鱼、叉线六线鱼[1]。关于六线鱼科鱼类中英文、拉丁文、分布及主要特征详见表1。
六线鱼科主要分布于西北太平洋水域的岩礁近岸水域, 包括阿拉斯加海域和阿留申群岛, 其中斯氏六线鱼(H. Stelleri)为最北品种, 在北极圈内也有分布。除了纬度上的区别外, 在垂直分布上, 不同六线鱼差异也很大, 例如斑头鱼和叉线六线鱼栖息在相对较浅的藻礁, 而大泷六线鱼栖息于较深的礁石水域[2-3]。六线鱼科鱼类无鳔, 除Ophiodon elongatus为圆鳞外, 六线鱼科的其他鱼类都为栉鳞(ctenoid scale)。六线鱼科中的六线鱼属和多线鱼属(Pleurogrammus)都具有多条侧线, 在系统分枝学上,多侧线性状为两个属的共源性状(synapomorphy)。其中, 六线鱼属的大泷六线鱼与叉线六线鱼都具有 5条侧线, 而且第4条都很短, 各个侧线的相对位置也类似, 形态上区别就是尾鳍的形状和羽状(皮质)突起的数目, 大泷六线鱼在眼后缘和枕骨上缘各具有2个羽状突起, 尾鳍截形; 而叉线六线鱼仅在眼后缘有1个羽状突起, 尾鳍圆形[2,4]。
叶青[5]研究了青岛近海大泷六线鱼的生长和食性, 发现幼鱼饵料中端足类、等足类栅幼蟹等常见,成鱼以鱼虾蟹为主, 但均属底栖动物食性。因此, 认为大泷六线鱼成鱼和幼鱼之间食性转换不明显[6]。
叶青在大泷六线鱼消化器官结构与功能适应性上做了很好的解释, 如上颌略长于下颌的特点有利于其摄食底栖动物; 上下颌的外行圆锥齿及犁骨的绒毛齿等, 适于捕食鱼类和头足类等游泳动物。另外咽齿的存在, 适于压碎贝螺等具硬壳的食物。鳃耙内侧具一列小刺, 适于滤食端足类等小个体生物[5]。
国内外学者对六线鱼科鱼类的形态习性、地理分布和繁殖习性进行了较为深入的研究, 而有关六线鱼科鱼类体色和繁殖特性的研究则鲜有报道, 作者就此进行了综述。
1 体色变化
在长期的自然选择和性选择的基础上, 鱼类适应外界环境条件, 尤其是群体间的社会交流, 会形成特定的体色模式以及伴随的相关行为学, 包括领地竞争、捕食以及防止被捕食、繁殖等[7-9]。六线鱼的体色变化丰富, 模式多样。涉及整个生活史的模式变化、种类间以及同种雌雄间的差异, 即性成熟后雌雄间体色二态性: 体色的日常变化和婚姻色等。
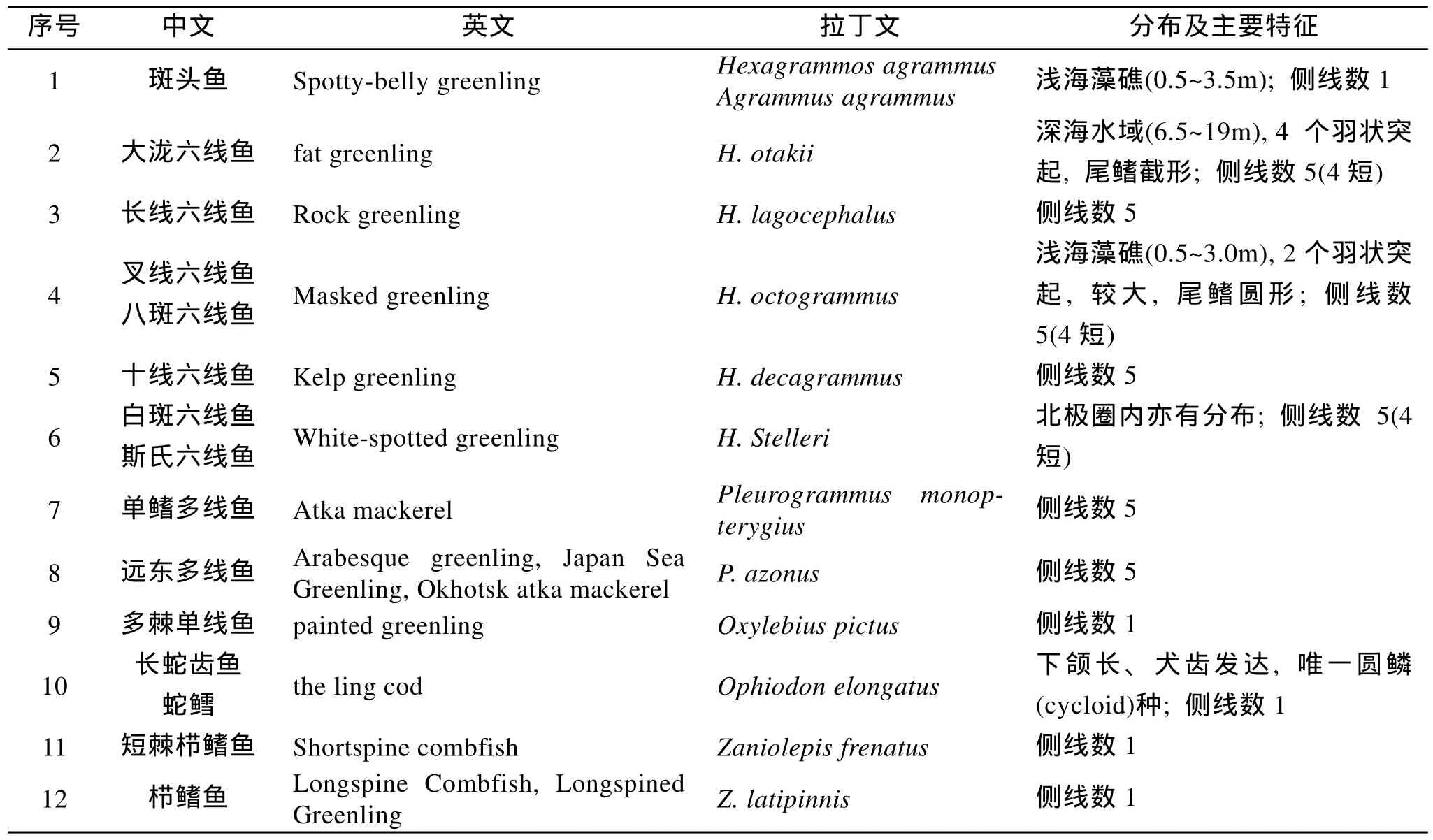
表1 六线鱼科鱼类的中英文、拉丁文和主要特征
1.1 体色的发育过程
英文 greenling的含义就是绿色小鱼的意思, 六线鱼鱼类在早期发育过程中, 包括卵本身就具有不同的颜色。据报道, 卵粒会呈灰白、黄橙、棕红、灰绿、墨绿等颜色[10]。在大泷六线鱼胚胎发育的后期,胚体开始呈现出蓝绿色体色, 而后随着发育的进行,整个仔鱼和稚鱼时期, 身体主要呈现为淡绿色, 直到变态阶段, 体色才逐渐转变为正常的黄色(图1)。有的六线鱼, 例如Oxylebius pictus和十线六线鱼,在性成熟后, 雌雄出现体色的性二态性(Sexual dimorphism)会持续一生[11], 这也难怪最初的学者将六线鱼的雌雄划分为两个物种[12], 甚至将幼鱼阶段和性成熟阶段划分为两个物种[13]。

图1 大泷六线鱼体色发育
1.2 体色的日常变化
大部分六线鱼一般营固定的底栖生活, 但是单鳍多线鱼(Pleurogrammus monopterygius)在每天的活动中出现一次垂直迁徙的过程[14], 这种迁徙严格与光照强度相关(白昼的时程), 在 150 m 水深光照强度大于0.004 lx就会生发垂直迁徙, 同时伴随着体色的变化。在白昼, 成群的单鳍多线鱼上升到水表面, 其身体的背部表面分布有黑色斑块, 而躯干其他为垂直的黑色条纹, 这样下面表现为灰色。在夜晚, 鱼群下降到海底藻床, 身体变为浅灰色, 更加不易为发现。这种体色变化的目的就是为了躲避其他鸟类、鱼类等的捕食[15-16],而其垂直迁徙的目的就是为了日常摄食需求, 单鳍多线鱼为典型的浮游动物食性(zooplanktivorous), 主要摄食磷虾类、挠足类和水蚤类[17]。在饱食后, 很多鱼群在下午或傍晚时分提前下降到海底藻床[15]。
1.3 婚姻色(nuptial coloration)
在繁殖季节出现的特定的体色变化(雌雄差异明显), 用以繁殖过程, 包括求偶、交配和护巢行为等的体色变化被称为婚姻色[18]。类似于其他硬骨鱼类, 大部分六线鱼(除了lingcodOphiodon elongatus)在繁殖季节也会出现婚姻色, 雄性的表现更为明显[19]。而且,六线鱼雄性的婚姻色的出现与护巢行为密切相关[20]。例如, 斑头鱼和叉线六线鱼雄性腹鳍和臀鳍变为黑色, 大泷六线鱼和单鳍多线鱼的雄性变为亮黄色[20-21]。一般来说, 婚姻色属于季节性体色变化, 会持续整个繁殖季节。更为特殊的种类——单鳍多线鱼,不仅具有至少两种不同的雄性婚姻色, 而且这种不同的雄性婚姻色与其特别殊的繁殖行为相关(包括求偶、繁殖和护巢过程), 持续时间较短, 类似于其晨昏垂直迁徙的体色变化[8]。
2 繁殖习性
六线鱼性成熟较早, 雌性六线鱼属于多次产卵类型, 以大泷六线鱼为例, 一般在2~3龄, 且雄鱼早于雌鱼。六线鱼怀卵量较少, 一般在万粒之内。产黏性卵, 卵刚产出时不黏, 后显黏性, 卵与卵之间互相黏着成团块, 并附着于海底岩礁、砾石、贝壳或海藻[22-23]。孵化时间较长(大泷六线鱼20 d, 单鳍多线鱼40~45 d)[15]。
在繁殖季节, 雄性六线鱼相互竞争, 在岩礁区域建立自己的繁殖领地, 雌性六线鱼到雄性六线鱼的领地产卵, 在单鳍多线鱼(通过雄鱼特殊的婚姻色)中, 这个时期在可以延长到7个月[16,24]。雌鱼产卵后,完成交配, 雄性则开始进行护巢直到仔鱼孵化(巢穴的数量从一个到10个不等, 范围可以扩大到1.9 m),且往往是几个雌鱼所产的卵[24]。同样, 在一个繁殖季节, 对于每个六线鱼雌性来说, 也不会仅仅在一个雄鱼的领地产卵。简言之, 六线鱼属于具有护巢行为的“多夫多妻制”繁殖模式[25], 类似的繁殖模式在其他硬骨鱼类中也较为常见[26]。在已知道的 422科硬骨鱼类中, 约有 21%的鱼类具有护巢行为, 护巢行为在进化上最大的意义就是大大提高仔鱼的成活率,大部分护巢行为由雄鱼来承担。
首先, 栖息地和护巢行为密切相关, 六线鱼属于浅海近岸种类, 类似与淡水水域; 具有水体较小,不开阔, 容易遭遇敌害生物等不利因素, 因此, 近岸种类和淡水鱼类一般都产黏性卵, 这样可以避免漂浮卵子被摄食, 护巢行为更是将这不利因素进一步缩小。
以单鳍多线鱼为例, 在繁殖季节存在两种类型的雄性, 其中一种为无自己巢穴(即无领地)的雄性,涉及寄生繁殖行为, 较为复杂[16]。从遗传的角度来考虑, 寄生繁殖雄性的存在已经使得六线鱼的“多夫多妻制”繁殖模式更加普遍和复杂化, 更利于种群的繁衍和多样化, 但是六线鱼却将此模式继续扩大至属间的自然杂交。
3 杂交情况
鱼类在自然界的杂交情况并不少见, 在很多鱼类中都存在杂交地带[27], 研究杂交地带有利于人们了解鱼类关于繁殖隔离和物种形成的进化过程[5],鱼类杂交地带的研究主要集中于淡水种类[28], 对于海洋鱼类人们知之甚少[29]。一般来说, 杂交地带在海洋中要比陆地宽广, 这就是为何海洋种群高度扩散的原因[30]。
在日本北海道南部海域和东北地区, 存在六线鱼属3种鱼类的自然杂交现象[2,20], 包括斑头鱼和叉线六线鱼[31], 大泷六线鱼与叉线六线鱼的杂交[32]。由于3种六线鱼都是产黏性卵, 需要附着基, 一般为苔藓或其他藻类或礁石等。人为破坏生态环境, 例如防浪堤(水泥块)的建设, 在浅海水域形成一个缓坡。这样, 很容易将本来隔离的较浅水域的繁殖地和较深水域的繁殖地联系起来, 形成镶嵌式的繁殖地和生态环境。通过人为的模拟实验同样证实了这一点[33]。
有趣的是, 两种自然杂交组合, 都是以叉线六线鱼为母本, 而且所有的杂交后代都表现为雌性[2],这说明了这类杂交具有典型的性联锁的不亲和性以及性联锁的后裔群现象; 其实类似的现象在其他硬骨鱼类中也有报道, 例如在淡水中, 利用具有天然雌核发育的方正银鲫为母本, 以兴国红鲤为父本形成的异育银鲫, 就是在充分考虑外源精子能影响到银鲫雌核发育子代的性能, 如生长、性比和体色等而形成的新品种[34]。虽然可以把这两种六线鱼的杂交组合称之为异育叉线六线鱼, 是否具有异育银鲫的效果还需要进一步研究和探讨。从另一个角度考虑,利用杂交组合来生产六线鱼的新品种, 因为这样的新品种不仅不影响亲本的优良性状, 更能为六线鱼的种质资源的保护以及六线鱼的人工繁育和养殖带来新的契机, 最后, 也能为鱼类天然雌核发育的生物学原理增添新的佐证, 进一步丰富鱼类繁殖理论和性别决定模式的基础理论。
4 展望
无论是环境因素, 还是人为造成的, 不可否认,中国沿海很多有经济价值的鱼类资源都已枯竭, 包括六线鱼类。随着人们对于渔业资源的合理利用, 一系列围绕渔业资源保护的措施正如火如荼的开展起来。对于衰退的鱼类资源来说, 较为重要的手段之一就是开展人工繁育工作。山东省海水养殖研究所从1998年开始就开展了六线鱼的人工养殖技术研究[10]。经过两代人的努力, 目前在该所鳌山卫养殖基地突破了六线鱼的人工繁殖技术难关, 解决了受精率和孵化率低的问题, 首次大规模培育6 cm以上大规格苗种 15.5万尾。受精率 97%, 孵化率 92%, 苗种成活率为43.2%。相关繁殖生物学、早期发育过程等基础研究已经相继开展。除此之外, 关于其他3种六线鱼的生物学数据很少, 相关养殖方面的报道更是缺乏。因此, 作者建议相关科研单位联合开展其他3种六线鱼的相关研究。
作者研究大泷六线鱼早期发育过程中发现其蓝绿色体色并非是皮肤体表的颜色, 可能是来源于体内骨骼的颜色, 因为鱼类本身很少含有蓝绿色素细胞, 类似于其他鱼类的骨骼体色成因。如扁颌针鱼(Ablennes anastomella)的绿色、鲣鱼(Katsuwonus pelamis)的蓝绿色, 以及其他种类包括杜父鱼属(Cottus), 绵鳚属(Zoarces)等, 由于骨内含有胆绿素(biliverdin)而呈现出蓝绿色的体色[35-36]。大泷六线鱼早期发育是否也是由于存在胆绿素还需要进一步证实。更为重要的, 为何六线鱼仅仅在早期发育过程中出现这种体色, 这种色素在体内的作用, 以及其在进化和生态上的意义, 更是值得人们深思。
[1]金鑫波. 中国动物志 硬骨鱼纲-鲉形目[M]//金鑫波.北京: 北京科学出版社, 2006: 550-561.
[2]Crow K D, Munehara H, Kanamoto Z,et al.Maintenance of species boundaries despite rampant hybridization between three species of reef fishes(Hexagrammidae): implications for the role of selection[J].Biological Journal of the Linnean Society,2007, 91(1): 135-147.
[3]Crow K D, Munehara H, Bernardi G. Sympatric speciation in a genus of marine reef fishes[J].Molecular Ecology, 2010, 19(10): 2089-2105.
[4]Wonsettler A L,Webb J F. Morphology and development of the multiple lateral line canals on the trunk in two species ofHexagrammos(Scorpaeniformes, Hexagrammidae)[J].Journal of Morphology,1997, 233(3): 195-214.
[5]叶青. 青岛近海欧氏六线鱼食性的研究[J].海洋湖沼通报, 1992, 14(4): 50-55.
[6]叶青,郑家声. 青岛近海欧式六线鱼(Hexagrammos otakiiJ &S)年龄和生长的研究[J].青岛海洋大学学报, 1993, 23(2): 59-68.
[7]Mills M G,Patterson L B. Not just black and white:Pigment pattern development and evolution in vertebrates[J].Seminars in Cell & Developmental Biology, 2009, 20(1): 72-81.
[8]Leclercq E, Taylor J F,Migaud H. Morphological skin colour changes in teleosts[J].Fish and Fisheries, 2010,11(2): 159-193.
[9]Hubbard J K, Uy J A C, Hauber M E,et al. Vertebrate pigmentation: from underlying genes to adaptive function[J].Trends in Genetics, 2010, 26(5): 231-239.
[10]庄虔增, 于鸿仙, 刘岗, 等. 六线鱼苗种生产技术的研究[J].中国水产科学, 1999, 6(1): 103-106.
[11]DeMartini E E. Social behavior and coloration changes in painted greenling,Oxylebius pictus(Pisces:Hexagrammidae)[J].Copeia, 1985, 4: 966-975.
[12]Mecklenburg C W, Eschmeyer W N. Family Hexagrammidae Gill 1889 - greenlings[J].Calif Acad Sci Annotated Checklists of Fishes, 2003, 3(11): 1-11.
[13]Shinohara G, Amaoka K.Stellistius katsukiiJordan &Tanaka, 1927, a junior synonym ofPleurogrammus azonusJordan & Metz, 1913 (Scorpaeniformes:Hexagrammidae)[J].Japanese Journal of Ichthyology,1994, 40(4): 487-490.
[14]Orlov A M. On the feeding of Atka mackerelPleurogrammus monopterygiusin the Pacific waters of the northern Kuril Islands[J].J Ichthyol, 1997, 337(3):226-231.
[15]Nichol D G, Somerton D A. Diurnal vertical migration of the Atka mackerelPleurogrammus monopterygiusas shown by archival tags[J].Marine Ecology Progress Series, 2002, 239: 193-207.
[16]Lauth R R, Guthridge J L, Cooper D W,et al.Behavioral ecology of color patterns in Atka Mackerel[J].Marine and Coastal Fisheries: Dynamics,Management, and Ecosystem Science, 2010, 2:399-411.
[17]Yang M. The trophic role of Atka mackerel,Pleurogrammus monopterygius, in the Aleutian Islands area[J].Fish Bull, 1999, 97: 1047-1057.
[18]Kodric-Brown A. Sexual dichromatism and temporary color changes in reproduction of fishes[J].American Zoologist, 1998, 38: 70-81.
[19]Lauth R R, Guthridge J, Nichol D G,et al. Timing of nesting, spawning, and brooding periods of Atka mackerel,Pleurogrammus monopterygius, in the North Pacific Ocean[J].US National Marine Fisheries Service Fishery Bulletin, 2007, 105: 560-570.
[20]Munehara H, Kanamoto Z, Miura T. Spawning behavior and interspecific breeding in three Japanese greenlings(hexagrammidae)[J].Ichthyological Research, 2000,47(3): 287-292.
[21]Zolotov O G. Notes on the reproductive biology ofPleurogrammus monopterygiusin Kamchatkan waters[J].Journal of ichthyology, 1993, 33(4): 25-37.
[22]冯昭信, 韩华. 大泷六线鱼资源合理利用的研究[J].大连水产学院学报, 1998, 13(2): 24-28.
[23]刘奇. 大泷六线鱼(Hexagrammos otakii)生物学特征与遗传多样性研究[D]. 青岛:中国海洋大学, 2010:5-15.
[24]Crow K D, Powers D A,Bernardi G. Evidence for multiple maternal contributors in nests of kelp greenling (Hexagrammos decagrammus, Hexagrammidae)[J].Copeia, 1997, 1: 9-15.
[25]King J R,Withler R E. Male nest site fidelity and female serial polyandry in lingcod (Ophiodon elongatus,Hexagrammidae)[J].Molecular Ecology, 2005, 14(2):653-660.
[26]DeWoody J A, Avise J C. Genetic perspectives on the natural history of fish mating systems[J].Journal of Heredity, 2001, 92(2): 167-172.
[27]Arnold M L. Natural Hybridization and Evolution[M].New York: Oxford University Press, 1997.
[28]Bleckmann H, Role of the lateral line in fish behaviour,in Behaviour of Teleost Fishes[M]. Chapman & Hall:London, 1993: 201-246.
[29]Suquet M, Dorange G, Omnes M H,et al. Composition of the seminal Fluid and ultrastructure of the spermatozoon of turbot (Scophthalmus maximus)[J].Journal of Fish Biology, 1993, 42(4): 509-516.
[30]van Herwerden L,Doherty P J. Contrasting genetic structures across 2 hybrid zones of a tropical reef fish,Acanthochromis polyacanthus(Bleeker 1855)[J].J Evol Biol, 2006, 19: 239-252.
[31]Balanov A A, Antonenko D V. First finding ofHexagramos agrammus×H. octogrammushybrids and new data about occurrence ofH. agrammus(Hexagrammidae) in Peter the Great Bay (The Sea of Japan)[J].Journal of Ichthyology, 1999(39): 149-156.
[32]Balanov A A, Markevich A I, Antonenko D V,et al. The fi rst occurrence of hybrids ofHexagrammos otakii×H.octogrammusand description ofH. otakiifrom Peter the Great Bay (The Sea of Japan)[J].Journal of Ichthyology, 2001, 41: 728-738.
[33]Kimura M, Munehara H. The disruption of habitat isolation among threeHexagrammosspecies by artificial habitat alterations that create mosaic-habitat[J].Ecological Research, 2011, 25(1):41-50.
[34]蒋一珪, 梁绍昌, 陈本德, 等. 异源精子在银鲫雌核发育子代中的生物学效应[J].水生生物学集刊, 1983,8(1): 1-11.
[35]Fox D, Millott N. A biliverdin-like pigment in the skull and vertebrae of the ocean skipjack,Katsuwonus pelamis(Linnaeus)[J].Cellular and Molecular Life Sciences, 1954, 10(4): 185-187.
[36]Cornelius C E. Bile pigments in fishes: A review[J].Vet Clin Pathol, 1991, 20: 106-115.
S917.4 文献标识码: A 文章编号: 1000-3096(2011)12-0132-05
2009-11-22;
2010-12-15
山东省资源修复计划“六线鱼苗种大规模人工繁育技术研究”; 山东省科技发展计划项目(2011GGB12008)
郭文(1963-), 男, 山东寿光人, 研究员, 主要从事海洋鱼类生物学研究, 电话: 0532-86513001, E-mail: yzszsjd@126.com; 于道德, 通信作者, 主要从事海洋鱼类生物学研究, E-mail:wensentte@163.com
梁德海)
