寄生养殖大菱鲆的海洋盾纤类纤毛虫(弗州拟尾丝虫相似种)的形态学和18S rDNA序列分析
2011-01-12李天保叶海斌盖春蕾朱安成
许 拉, 李天保, 叶海斌, 盖春蕾, 朱安成, 张 伟
(山东省海水养殖研究所 山东省海水养殖病害防治重点实验室, 山东 青岛 266002)
寄生养殖大菱鲆的海洋盾纤类纤毛虫(弗州拟尾丝虫相似种)的形态学和18S rDNA序列分析
许 拉, 李天保, 叶海斌, 盖春蕾, 朱安成, 张 伟
(山东省海水养殖研究所 山东省海水养殖病害防治重点实验室, 山东 青岛 266002)
从养殖大菱鲆(Scophthalmus maximus)患病鱼体中分离出一种寄生纤毛虫, 通过活体观察、碳酸银法染色对其形态学特征进行分析, 初步鉴定为弗州拟尾丝虫相似种(Parauronemacf.virginianumThompson, 1967)。同时对其18S rDNA序列进行了测定和分子系统发育学分析, 结果显示该纤毛虫序列与弗州拟尾丝虫(Parauronema virginianum)存在8个碱基的差异, 序列相似性为99%, 表明二者的高度相似性。在对本纤毛虫的18S rDNA序列和GenBank中报道的14种相关盾纤类纤毛虫相应序列相似性和进化距离分析的基础上, 作者构建了包括迄今所有相关、已知种类的系统进化树。本研究为海水鱼类相关病害的病原鉴定提供了一份新的数据和资讯。
大菱鲆(Scophthalmus maximus); 盾纤虫; 弗州拟尾丝虫(Parauronema virginianum); 碳酸银染色; 18S rDNA
大菱鲆(Scophthalmus maximus)自1992年引入中国以来, 取得了良好的经济效益。然而近几年来, 除细菌和病毒性疾病外, 寄生虫病的危害也越来越突出, 其中由嗜组织的盾纤类纤毛虫所引起的危害十分突出, 产生了严重的经济损失[1-6]。
据文献报道, 迄今国内从养殖鲆鲽类病鱼体表溃烂组织中分离出的该类纤毛虫中, 已鉴定出有贪食迈阿密虫(Miamiensis avidus), 蟹栖异阿脑虫(Mesanophrys carcini), 指状拟舟虫(Paralembus digitiformis) 以及水滴伪康纤虫(Pseudocohnilembus persalinus)[2-6]。
作者新近从大菱鲆病鱼组织中分离出一种纤毛虫, 形态学分析, 初步鉴定为弗州拟尾丝虫相似种;并测定了该纤毛虫的18S rDNA基因序列, 分析比较了与之相关的14种盾纤毛虫相应序列的同源性, 构建了系统发生树, 从分子序列方面进一步确定了该纤毛虫的系统分类地位[7]。
1 材料与方法
1.1 盾纤毛虫的培养
2007年8月至2008年6月, 从山东烟台、威海、青岛等地患病养殖大菱鲆取样, 自体表、鳃和内脏分离出病原纤毛虫, 过滤海水为培养液, 加新鲜大菱鲆鱼肉, 室温培养2~3 d。
1.2 纤毛虫的形态学与分类鉴定
形态学研究方法包括: 光学显微镜活体观察、碳酸银染色法[8-9]。本文采用的分类系统参照Corliss(1994)。
1.3 DNA的提取
虫体经福尔马林固定后, 3 000~3 500 r/min离心富集, 取富集虫体培养液50 µL, 加入450 µL裂解缓冲液(10 mmol Tris-HCl, pH 8.3; 50 mmol KCl; 2.5 mmol MgCl2; 0.6%Tween 20; 0.6%Nonidet P40; 60 mg/L Proteinase K), 56°C水浴1~2 h, 常规酚氯仿法抽提, 用50 µL灭菌双蒸水溶解DNA沉淀, -20℃保存备用[10]。
1.4 PCR扩增18S rDNA基因序列
选择 Elwood等[11]报道的通用引物 16S-like-F:5’-AACCTGGTTGATCCTGCCAGT-3’, 16S-like-R:5’-TGATCCTTCTGCAGGTTCACCTAC-3’进行扩增,该引物能扩增大多数盾纤类纤毛虫近乎全长的小亚基 rRNA基因[12], 由上海生工生物有限公司合成。50 µL的PCR反应体系中包含: 1× PCR缓冲液, 1.5 mmol/L MgCl2, 200 µmol/L dNTP 混合物, 1 µmol/L引物16S-like-F和16S-like-R, 1µL DNA模板。PCR反应条件为: 94℃预变性5 min后, 加入Taq DNA聚合酶, 初始的5个循环为94°C变性1 min, 56°C退火2 min, 72°C延伸2 min, 后35个循环将退火温度提升为62℃, 最后72°C延伸5 min, 于4°C结束反应。用1%的琼脂糖电泳, Genefinder染色后, 在紫外下观察条带, DL2000用做DNA分子量标准。
1.5 基因的克隆与测序
采用北京博日胶回收试剂盒纯化回收PCR产物,利用T-A连接, 按pMD18-T载体说明书将纯化后适量的PCR产物直接连接至pMD18-T载体中, 按常规方法转化到Escherichi coli感受态细胞, 使用选择性LB琼脂平板筛选含重组质粒的白色克隆, 接于含有Amp的LB培养基过夜培养, 通过PCR验证后送大连宝生物有限公司进行DNA双向测序。
1.6 DNA序列分析及数据处理
将测序所得到的纤毛虫18S小亚基 rDNA序列在 GenBank数据库中 BLAST查找, 收集相关的 14种盾纤目纤毛虫的小亚基 rRNA基因序列, 采用ClustalW1.83软件进行多序列比对(Multiple Alignment), 用系统进化分析软件 MEGA4.1进行统计和聚类分析。用Kimura双参数模型计算各序列分化距离, 缺少和不确定位点在计算中被省略, 采用邻接法(neighbor-Joining method)获得系统发生树, 并通过自举分析(bootstrap)进行置信度检测, 自举数据集为1000次。
2 结果
2.1 形态学
由于种种原因, 我们在对所获虫体的观察和研究中, 虽然没有得到详细的口器结构等全面信息,但根据宋微波[13]对发现自濒死幼虾体内的弗州拟尾丝虫(Parauronema.virginianum)的描述, 本研究的样本与该种在大小、主要形态特征上均相似。考虑到该类群普遍具有的兼性寄生的特性, 可以认定本寄生纤毛虫应为弗州拟尾丝虫的相似种。其主要形态特征如下:
活体约 36µm×16µm(长×宽), 外形稳定, 呈瓜子型, 皮膜无缺刻, 体前部具多个晶体, 内质无色,中央区为较透明的大核。虫体顶端略尖, 呈小的截面,为裸毛区(AP)。口区约占体长的 1/2, 体纤毛长约6~8 µm, 尾纤毛约 18 µm。伸缩泡直径可达 5 µm, 位于体后端。运动呈旋转式快速前进。虫体喜聚集在营养丰富的基质上, 成极高的密度, 并可通过横二分裂快速繁殖(图1 A-C)。
银染结果显示, 本种体动基列为10-12列。虫体近顶端的小膜 1(M1)呈短三角形, 由数个毛基粒组成, 小膜 2(M2)为相互平行的 2排纵行的毛基粒列,小膜3为斜向的2排毛基粒列。口侧膜(PM)起始于M2中部, 至 M3前端为一单列毛基粒, 后变为双动基列构造, 并至胞口(Cs)后。大、小核各一个, 位于体中部。大核(Ma)为不规则的椭圆形, 长 10.6 µm,小核(Mi)近球形, 长 3 µm, 紧位于大核侧边(图1 D-F)。
2.2 18S rDNA基因PCR克隆
以提取的DNA为模板, 16S-like-F和16S-like-R为引物, 扩增出 1条约为 1.7 kb的目的片段。PCR产物回收纯化后与pMD18-T载体连接、转化, 筛选得到阳性克隆, 将得到的重组质粒进行 PCR扩增鉴定, 结果如图2。
挑选阳性克隆经DNA双向测序, 得到1条长度为1758 bp的近乎全长的18S rDNA基因片段, GC含量为 42.4%, 片段大小和组分与相近属种的已知序列相近。所得序列已提交 GenBank, 登录号为FJ595488。
2.3 序列比较与系统进化分析
将测定的该纤毛虫 18S rDNA基因序列与GenBank内登录的14种不同种属的盾纤类纤毛虫相应序列进行比对, 序列间的同源性和进化距离分析见表2。如表所示, 实验纤毛虫与 14种纤毛虫的序列相似度在 79%~99%之间, 而与Parauronema. virginianum(AY392128)相似度最高, 达到99%, 有7个碱基的差异, 与其遗传距离为 0.01。采用的 MEGA 4.1软件构建的分子进化树显示新测序列(FJ595488)与Parauronema.virginianum(AY392128)以最高置信值(100%)聚为一枝, 之后与Uronema marinum(DQ867072)相聚(100%), 而与其他种类相距较远(图3)。

图1 弗州拟尾丝虫相似种的形态学特征Fig. 1 Morphological characteristics of Parauronema cf. virginianum derived from cultured turbot

表1 弗州拟尾丝虫相似种的统计学特征(自碳酸银制片标本, 测量单位: µm)Tab. 1 Biometrical characterrization of Parauronema cf. virginianum (Data based on specimens prepared by silver carbonated staining)
3 讨论
目前国内外已有大菱鲆纤毛虫病的报道, 证明了感染大菱鲆的病原纤毛虫不止一种, 并且无明显的宿主选择性。西班牙学者Iglesias等鉴定感染大菱鲆的病原为Philasterides dicentrarchi。而后, Song和Wilbert[3]等根据 Iglesias提供的详细资料, 将其更正为贪食迈阿密虫(Miamiensis avidusThompson, 1964),国内王印庚等[4]报道了大菱鲆盾线虫病是由一种嗜组织的蟹栖异阿脑虫(Mesanophrys carcini)寄生而引起的, 并在病原纤毛虫流行病学, 病原学、组织病理学及防治方法等方面做了较详细的研究。
本文所涉的病原纤毛虫与徐奎栋、宋微波等[13-14]分离自贝类中的弗州拟尾丝虫形态特征有较大的相似性, 但在口器小膜1(M1)和小膜3(M3)略有不同。结合分子信息, 我们可以认定此危害种为一与弗州拟尾丝虫高度相似的拟尾丝虫, 由其引起养殖大菱鲆盾纤虫病, 在国内属首次报道。此也表明了感染大菱鲆的病原纤毛虫不止一种。
在对其18S rDNA序列比较分析中, 本文克隆测序所得序列(FJ595488)与弗州拟尾丝虫AY392128序列同源性为 99%, 遗传距离为 0.005, 在构建的分子系统发育树中高置信度地聚为一支, 而与之前学者报道的感染大菱鲆的贪食迈阿密虫(Miamiensisavidus) AY550080、蟹栖异阿脑虫(Mesanophrys carcini)AY103189的相似性分别为 91%、92%, 遗传距离分别为0.09、0.10, 与之前报道的感染牙鲆的指状拟舟虫(Paralembus digitiformis)AY297715、水滴伪康纤虫(Pseudocohnilembus persalinus)AY835669的相似性分别为91%、79%, 遗传距离分别为0.10、0.25。在构建的分子系统发育树中上述 4种纤毛虫18S rDNA序列分别聚类在4个不同分支上, 与本文测定的纤毛虫18S rDNA序列(FJ595488)不在一个类群分支上。通过分析比较18S rDNA序列进一步验证本文研究的纤毛虫与弗州拟尾丝虫亲缘关系最近, 而与之前报道的寄生类纤毛虫有明显的差异。鉴于对本种的分类学信息仍欠完备, 此妨碍了种类鉴定的进一步开展, 我们期待在后期的工作中完成对此寄生虫的廓清工作。

图2 18S rDNA基因PCR扩增产物、回收纯化及重组质粒PCR扩增结果Fig. 2 The results of 18S rDNA gene PCR amplification,recovery, purification and recombinant plasmid PCR amplification

表2 15种相近盾纤类纤毛虫18S rDNA基因序列相似性和进化距离Tab. 2 Sequence similarity and evolutionary distance of 18S rDNA gene of 15 ciliates
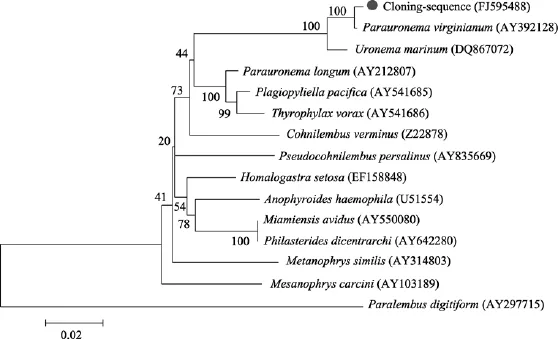
图3 以18S rDNA基因序列构建的相关类群的系统发育树Fig. 3 Phylogenetic relationship of related ciliates based on 18S rDNA gene sequences
[1]Yoshinaga T, Nakazoe J. Effects of light and rotation culture on thein vitrogrowth of a ciliate causing the scuticociliatosis of Japanese flounder [J]. Fish Pathol,1997, 32: 227-228.
[2]Iglesias R, Paramá A, Alvarez M F, et al.Philasterides dicentrarchi(Ciliophora, Scuticociliatida) as the causative agent of scuticociliatosis in farmed turbotScophthalmus maximusin Galicia(NW Spain) [J]. Dis Aquat Org, 2001, 46: 47-55.
[3]Song W, Wilbert N. Redefinition and redescription of some marine scuticociliates from China, with report of a new speciesMetanophrys sinensisnov. spec. (Ciliophora, Scuticociliatida) [J]. Zoologisches Anzeiger,2000, 239: 45-74.
[4]王印庚, 陈洁君, 秦蕾.养殖大菱鲆蟹栖异阿脑虫感染及其危害[J].中国水产科学, 2005, 12: 594-601.
[5]周丽, 徐奎栋, 战文斌, 等.一种纤毛虫的分类及形态研究——寄生于牙鲆体表溃烂组织中的指状拟舟虫[J].青岛海洋大学学报, 2001, 31(2): 190-194.
[6]张立坤, 王玉梅, 肖国华, 等.牙鲆体表溃烂组织中的水滴伪康纤虫[J].河北渔业, 2007, 10: 42-43.
[7]孙春燕, 赵元莙.DNA序列分析在黏孢子虫系统学研究和病原检测中的应用[J].中国水产科学, 2006,13(1): 159-164.
[8]宋微波, 徐奎栋.纤毛虫原生动物形态学研究的常用方法[J].海洋科学, 1994, 6: 6-8.
[9]宋微波.原生动物学专论[M].青岛: 青岛海洋大学出版社, 1999.
[10]陈子桂, 容丽, 宋微波.海洋尾丝虫(Uronema marinum)的DNA微量提取研究[J].动物学报, 2001, 47(专刊): 6-8.
[11]Elwood H J, Olsen G J, Sogin M L. The small-subunit ribosomal RNA gene sequences from the hypotrichous ciliatesOxytricha novaandStylonychia pustulata.[J].Mol Biol Evol, 1985, 2: 399-410.
[12]Schlegel M, Elwood H J, Sogin M L. Molecular evolution in hypotrichous ciliates: sequence of the small subunit ribosomal RNA genes fromOnychodromus quadricornutusandOxytricha granulifera(Oxytrichi-dae, Hypotrichida, Ciliophora)[J]. J Mol Evol, 1991, 32:64-69.
[13]宋微波.海水养殖中危害性原生动物[M].北京: 科学出版社, 2003.
[14]徐奎栋, 宋微波.海洋贝类几种危害性纤毛虫的研究[J].中国水产科学, 1999, 6(2): 41-45.
Morphological studies and 18S rDNA sequence analysis of the marine scuticociliateParauronemacf.virginianum(Protozoa,Ciliophora) isolated fromScophthalmus maximus
XU La, LI Tian-bao, YE Hai-bin, GAI Chun-lei, ZHU An-cheng, ZHANG Wei
(Shandong Mariculture Research Institute, Key Laboratory for Mariculture Disease Treatment of Shandong Province, Qingdao 266002, China)
Dec., 23, 2010
Scophthalmus maximus; Scuticociliatosis;Parauronemacf. virginianum; silver carbonate staining; 18S rDNA
A parasitic ciliate isolated from the cultured flatfishScophthalmus maximuswas analyzed in the present paper. Morphological investigations were performed using living observation and silver carbonate staining method.The data of the body shape, size, structure of buccal apparatus, macro- and micronucleus, as well as the somatic ciliature showed that the parasitic ciliate was similar toParauronema virginianum. The molecular phylogenetic characters were analyzed to further determine the classification of the parasitic ciliate. The 18S rDNA of the ciliate was sequenced (GenBank accession number is FJ595488), and it only had 8 different bases compared with those ofP. virginianum. They shared 99% similarity insequence. The 18S rDNA of the ciliate was also compared with those of 14 related ciliates. Phylogenetic dendrogram was constructed based on the genetic distance analysis, which revealed that this organism andP. virginianumclustered into one branch, with their genetic distance of 0.005. Based on the obtained results, the isolated ciliate was identified as aParauronema-similar taxon, that is,P.cf. virginianum.
S941 Q959 文献标识码: A 文章编号: 1000-3096(2011)12-0036-06
2010-12-23;
2011-04-19
山东省科技攻关项目(2007GG10005006)
许拉(1981-), 男, 湖南湘潭人, 硕士, 主要从事水产动物病害防治学研究, E-mail: gene05@163.com; 李天保, 通信作者, 研究员,主要从事水产养殖病害防治, E-mail; ltb1601@126.com
张培新)
