植物乳杆菌素抑菌机理的研究
2011-01-08程建军李想郭明若
程建军,李想,郭明若
(1.东北农业大学,哈尔滨 150030;2.双城雀巢有限公司,哈尔滨 150100;3.佛蒙特大学,美国伯灵顿 05403)
植物乳杆菌素抑菌机理的研究
程建军1,李想2,郭明若3
(1.东北农业大学,哈尔滨 150030;2.双城雀巢有限公司,哈尔滨 150100;3.佛蒙特大学,美国伯灵顿 05403)
从植物乳杆菌B-28的代谢产物中分离纯化植物乳杆菌素,以蜡样芽胞杆菌为指示菌,通过测定指示菌细胞结构和细胞中APT、离子量的变化,分析探讨植物乳杆菌素的抑菌机理。研究结果表明:当植物乳杆菌素作用于蜡样芽胞杆菌时,胞内ATP呈现迅速下降趋势;并迅速释放出K+和Na+,而且植物乳杆菌素的质量分数与K+和Na+释放量基本呈现线性正相关。随着植物乳杆菌素添加量的增加,蜡样芽胞杆菌细胞裂解率在逐渐提高。而且电镜观察发现:植物乳杆菌素作用的蜡样芽胞杆菌的细胞壁边缘模糊,菌体细胞变形,部分细胞壁缺失,可看到胞内物质的损失。
植物乳杆菌素;抑菌;机理
0 引言
乳酸菌可产生对食品中微生物具有抑制作用的酸性物质,主要是乳酸菌的代谢终产物及中间产物[1,2]。而且有些乳酸菌还能代谢产生具有抑菌活性的乳酸菌素,这类细菌素主要是Nisin和Ⅱa类细菌素,其抗菌机理的研究结果表明细菌素作用的靶目标是细胞膜。
不同的乳酸菌素由于结构不同,其活性和对细胞膜的作用方式也不同。Nisin的抑菌作用最初被认为是一种表面活性剂;亦有人认为Nisin的作用是由于其中的两个脱水氨基酸(脱水丙氨酸和脱水丁氨酸)与细菌细胞中酶的巯基发生反应;目前认为除了以上两种原因外,还存在孔道形成机制[3,4]。Ⅱa类细菌素的抗菌机理在近几年才逐渐为人们所研究,其主要是由于细菌素吸附在细胞膜上,并形成孔道,使得细胞膜的通透性增加从而引起细胞内各种离子的渗漏和能量物质的消耗,导致细胞解体死亡[5]。
1 材料和方法
1.1 菌株
植物乳杆菌B-28(Lactobacillus plantarumB-28);从保加利亚传统发酵小麦饮料中分离得到。蜡样芽胞杆菌(Bacillus cereus)。
1.2 主要试剂
XRA细菌细胞释放剂,萤火虫荧光素。
1.3 主要仪器设备
电子显微镜,BIO-680全自动酶标仪,BHP9505微量光度计,PE-300原子吸收分光光度计,SP-722分光光度计。
1.4 方法
1.4.1 ATP测定
ATP测定由张东声、Sirugusa和Naghmouchi方法修正后进行测定[6-8]:对数生长期的蜡样芽胞杆菌(Bacillus cereus),1 480 g离心15 min,收集菌体,洗涤后的悬浊液溶于浓度为50 mmol/L(pH值为6.0)的MOPS缓冲溶液中,625 nm的吸光值为0.02。
添加葡萄糖到菌悬液中,终点的浓度达到0.5 mmol/L,5 min后取100 μL样品加入100 μL缓冲溶液,混合后立即用0.45 μL微量滤器过滤,滤液用于细胞外ATP测定,滤器上的细胞用于细胞内ATP测定,测定结果用作对照。
6 min时,分别添加0.5 mL和1 mL植物乳杆菌素到细胞悬浊液中,每隔5 min,取出100 μL样品过滤,滤器上的细胞中加入50 μL的细胞裂解液和50 μL的荧光素,迅速搅动3次后,置于微型荧光计中,10 s后的相对光单位(Relative Light Units,RLU)为胞内ATP值。
取50 μL滤液,加入50 μL的荧光素,迅速搅动3次后,置于微型荧光计中,相对光单位RLU为胞外ATP值。以时间为横坐标,以相对光单位RLU为纵坐标,绘制ATP变化曲线。
1.4.2 细胞裂解率的测定
细胞裂解率测定由Naghmouchi(2006)方法修正后测定[8]:10 mL培养6h在4℃下,10 000 g离心10 min,洗涤后的悬浊液溶于5 mL浓度为20 mmol/L(pH值为6.0)的磷酸盐缓冲溶液中。
将细胞悬浮液125 μL和25,50,75,100,125 μL的植物乳杆菌素的磷酸盐缓冲溶液混合,总量保持在250 μL,37℃下培养,测定630 nm的吸光值。细胞裂解率为
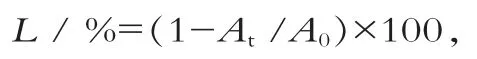
式中:At为9 h或12 h的吸光值;A0为开始时的吸光值。以植物乳杆菌素添加量为横坐标,以细胞裂解率为纵坐标作图。
1.4.3 细胞离子流出的测定
对数生长期的蜡样芽胞杆菌(Bacillus cereus),4℃下,10 000 g离心10min,菌体细胞用浓度为50 mmol/ L(pH值为6.0)的MOPS缓冲溶液洗涤两次,溶于质量浓度为2 g/L葡萄糖的MOPS缓冲溶液中,分别添加0.5 mL和1 mL植物乳杆菌素,30℃下培养,每隔5 min,0.45 μL微量滤器过滤后,采用原子吸收分光光度计,根据GB/T 5009.124-2003的方法,测定K+和Na+的总量。以时间为横坐标,以K+和Na+的质量分数为纵坐标,绘制离子流出曲线。
1.4.4 电镜观察细胞的变化
根据杨向科、Beatriz Gonzalez和Naghmouchi方法[8-10],分别添加植物乳杆菌素500 μL和1 mL的对数生长期的蜡样芽胞杆菌(Bacillus cereus),4℃下,10 000 g离心10 min,收集菌体细胞,4℃下0.05%戊二醛和溶于pH值为7.2磷酸盐缓冲溶液的2.5%多聚甲醛中固定2 h,样品每5 min冲洗1次;4次后,用质量浓度为30 g/L琼脂糖微包埋,4℃下含质量浓度为1 g/L的四氧化锇的磷酸盐缓冲溶液固定2 h,洗涤4次后,乙醇梯度脱水、包埋、60℃聚合48 h。切片,磷酸盐缓冲溶液清洗,乙酸双氧铀和柠檬酸铅染色,电镜观察细胞结构变化。
2 结果
2.1 ATP的测定
以时间为横坐标,以相对光单位RLU为纵坐标,绘制ATP(Adenosine-Triphosphate)释放变化曲线,结果如图1所示。
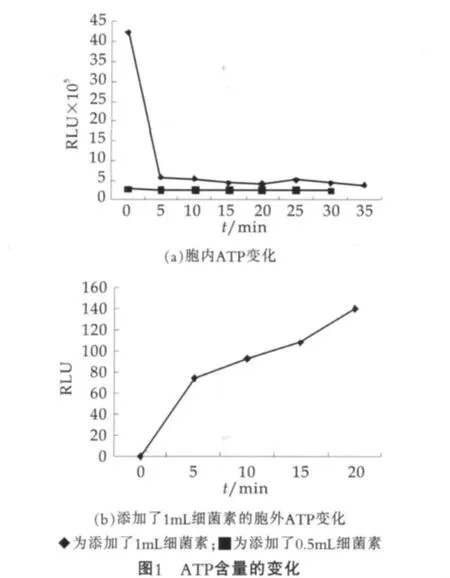
由图1可以看出:加入了植物乳杆菌素的蜡样芽胞杆菌胞内和胞外ATP都发生了变化,添加了1mL植物乳杆菌素的胞内ATP在5 min添加后迅速下降,然后呈现缓慢下降趋势,并趋于平稳;胞外ATP迅速增加;而添加了0.5 mL植物乳杆菌素胞内ATP虽然出现缓慢下降趋势,但变化幅度明显小于1 mL植物乳杆菌素添加量时ATP的变化。
这说明了植物乳杆菌素破坏了蜡样芽胞杆菌的细胞壁,完整细胞内的ATP释放出来,这个结果与Waite等、Naghmouchi等的研究结果[8,11]是一致的,这可能是由于植物乳杆菌素抑制了依赖于磷酸转移酶体系的磷酸烯醇式丙酮酸(PEP)的合成,从而造成了胞内ATP缺失。
2.2 细胞裂解率的测定
以植物乳杆菌素添加量为横坐标、细胞裂解率为纵坐标绘制的植物乳杆菌素在不同作用时间的蜡样芽胞杆菌细胞裂解率,结果如图2所示。
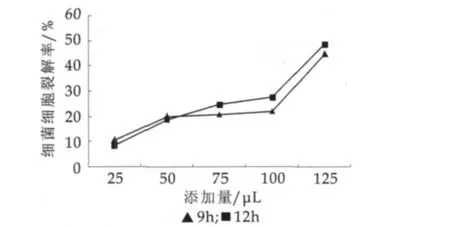
图2 蜡样芽胞杆菌细胞裂解率
由图2可以看出,随着植物乳杆菌素添加量的增加,蜡样芽胞杆菌细胞裂解率在逐渐提高,作用9 h和12 h裂解率的变化趋势是一致的。开始时,细菌裂解率增加;然后添加量50~100 μL间保持相对稳定状态;在100 μL植物乳杆菌素的添加量时,细胞裂解率出现了突越,植物乳杆菌素添加量为125 μL(相当于0.625 mg)时达到了最高,为48.52%。这说明了植物乳杆菌素抑制或杀灭蜡样芽胞杆菌需要有效的浓度,只有当浓度达到最适时,抑菌或杀菌效果才会达到最佳。
2.3 细胞内钾、钠离子流出的测定
以时间为横坐标,以从蜡样芽胞杆菌中释放出的K+和Na+质量分数为纵坐标,绘制离子流出曲线,结果如图3所示。
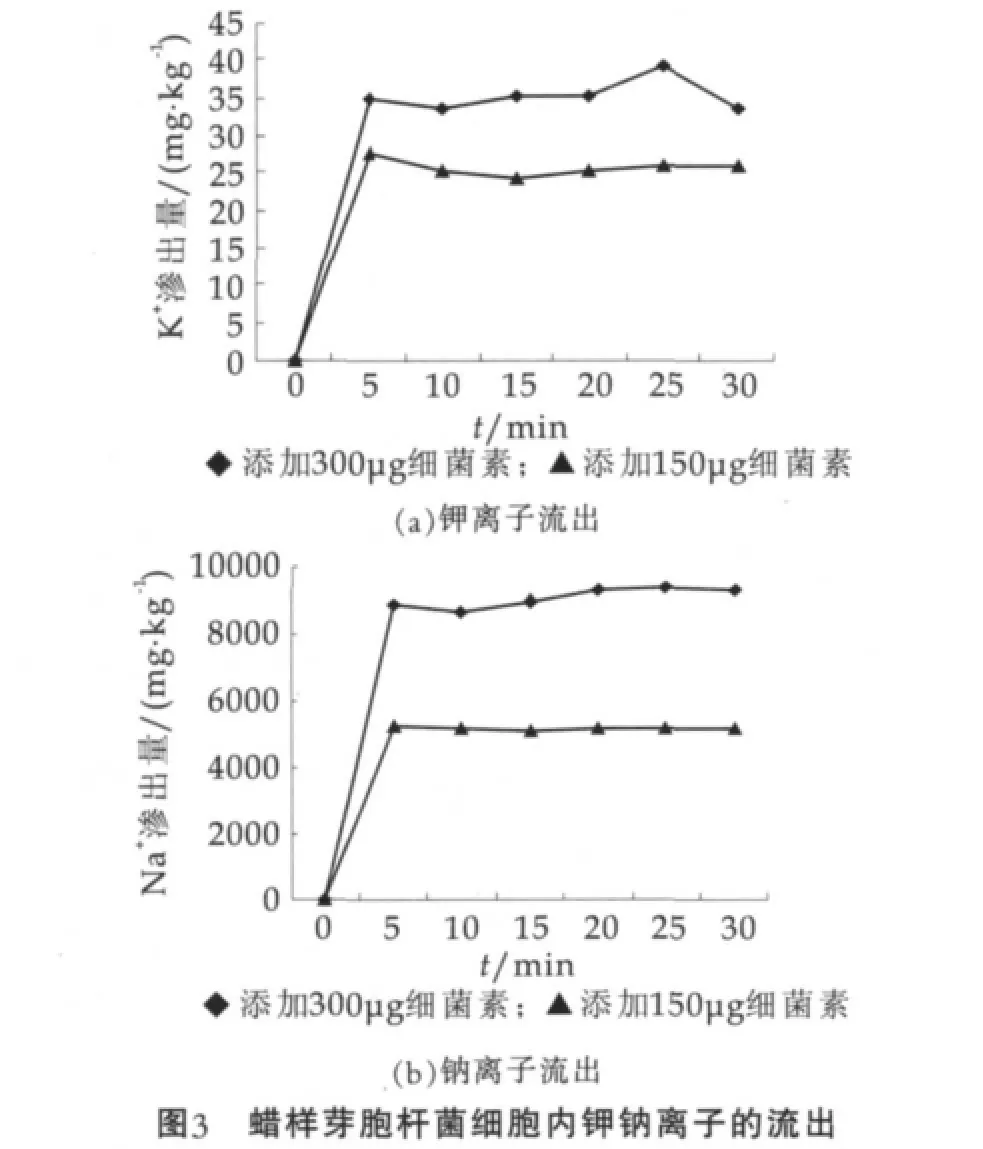
由图3可以看出,因植物乳杆菌素作用而释放出K+和Na+质量分数在开始时,迅速释放,5 min后达到稳定状态;300 μg和150 μg植物乳杆菌素处理的蜡样芽胞杆菌的K+和Na+释放量基本呈现倍数,表明了植物乳杆菌素的质量分数与K+和Na+释放量基本呈现正相关。
图3中,钠离子的释放量高于钾离子,最高达到了9203 mg/kg,而钾离子最高只有39.24 mg/kg。这可能是由于蜡样芽胞杆菌对NaCl具有较强的适应能力,在营养肉汤培养基中就含有质量分数为5%的NaCl,细胞本身就含有较高钠离子的缘故。植物乳杆菌素可能是通过穿透指示菌的细胞膜来杀死细菌的,当两肽细菌素相互作用形成两性的α-螺旋结构而插入指示菌的细胞膜中,形成了钾离子的选择性通道[12]。
2.4 电镜观察细胞的变化
植物乳杆菌素对蜡样芽胞杆菌作用的形态结构电镜观察结果如图4所示。由图4可以看出,图4(a)和(d)的蜡样芽胞杆菌细胞壁连续、完整、光滑,结构紧密,细胞质均匀。图4(b)、(c)、(e)和(f)植物乳杆菌素作用的蜡样芽胞杆菌细胞壁边缘模糊,细胞壁上有凹陷,可看到胞内物质的损失,部分细胞壁缺失,菌体细胞变形,有些细胞质甚至解体出现空腔。而且随着植物乳杆菌素浓度的增加,蜡样芽胞杆菌细胞壁、细胞膜和细胞质的变化越来越明显。说明了植物乳杆菌素溶解了细胞壁,对腊样芽胞杆菌的生长起到了抑制作用。
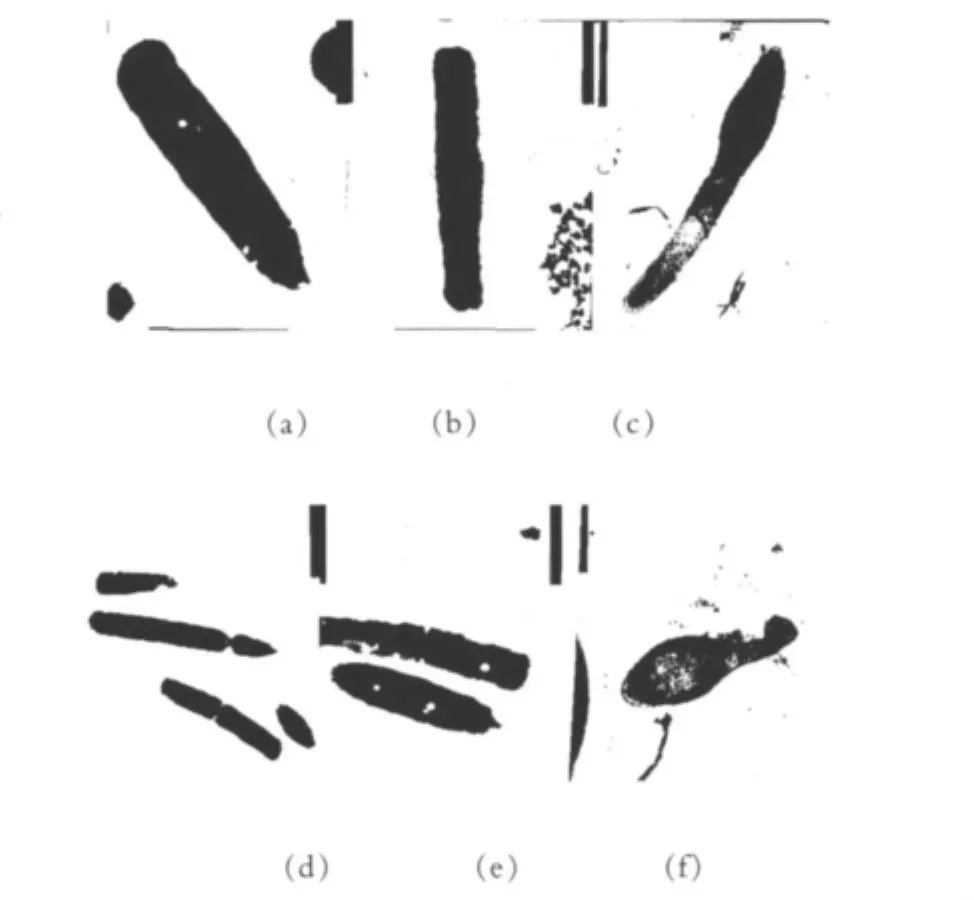
图4 电镜观察结果
图4中,(a)、(d)为对照;(b)、(e)为500 μL植物乳杆菌素处理;(c)、(f)为1 000 μL植物乳杆菌素处理。
3 结论
加入了植物乳杆菌素的蜡样芽胞杆菌胞内ATP呈现迅速下降趋势,并逐渐趋于平稳;胞外ATP值迅速增加;而添加了0.5 mL植物乳杆菌素胞内ATP值虽然出现缓慢下降趋势,但变化幅度明显小于1mL植物乳杆菌素添加量ATP值的变化。
植物乳杆菌素作用而释放出K+和Na+值在开始时,迅速释放,5 min后释放趋于稳定状态;而且植物乳杆菌素的浓度与K+、Na+释放量基本呈现线性正相关。
随着植物乳杆菌素添加量的增加,蜡样芽胞杆菌细胞裂解率在逐渐提高,125 μL达到了最高,为48.52%。9 h和12 h裂解率的变化趋势是一致的。
电镜观察蜡样芽胞杆菌细胞,与对照比较,细胞壁边缘模糊,可看到胞内物质的损失,部分细胞壁缺失、细胞膜破裂,菌体细胞变形;有些细胞质甚至解体出现空腔。而且随着蜡样芽胞杆菌培养时间的延长,细胞壁、细胞膜和细胞质的变化越来越明显。
[1]李铁军,李爱云,张晓峰.乳酸菌抗菌机理研究进展[J].微生物学通报. 2002,29(5):81-85.
[2]郭本恒.细菌素在肉品保藏中的应用[J].食品研究与开发.2003,22(1):49-50.
[3]侯瑞,饶贤才,胡福泉.羊毛硫抗生素研究进展[J].微生物学杂志. 2006,26(6):69-73.
[4]MCAULIFFEO,PAULROSSRC.Lantibiotics:Structure, Biosynthesis and Mode of Action[J].Microbiology Reviews,2001,25: 285-308.
[5]ENNAHARS,TOSHIHIROS,KENJIS,etal.ClassIIa Bacteriocins:Biosynthesis,Structure and Activity[J].Microbiology Reviews,2000,24:85-106.
[6]张东声,王春生,杨俊毅,等.荧光法测定微型生物细胞内的ATP的技术[J].海洋学研究,2006,24(1):73-81.
[7]SIRAGUSA G,GUTTER,DORSA C N,et al.MUse of a Rapid Microbial ATP Bioluminesescence Assay to Detect Contamination of Beef and Pork Carcasses[J].Journal of Food Protection,1995,58,770-775.
[8]NAGHMOUCHI K,DRIDER D,FLISS I.Action of Divergicin M35, a Class IIa Bacteriocin,on Liposomes and Listeria[J].Journal of Applied Microbiology,2006,9:1-10.
[9]杨向科,邹艳丽,孙谧,等.海洋微生物溶菌酶的抑菌作用及抑菌机理初步研究[J].海洋水产研究,2005,26(5):62-68.
[10]Beatriz Gonzalez,Glaasker Erwin,Edmund R.S.Kunji,et al.Bactericidal Mode of Action of Plantaricin C[J].Applied and Environmental Microbiology.1996,8:2701~2709
[11]WAITE B L,SIRAGUSA G R,HUTKINS R W.Bacteriocins Inhibit Glucose PEP:PTS Activity in Listeria monocytogenes by Induced Efflux of Intracellular Metabolites[J].Journal of Applied Microbiology,1998,85:287-292.
[12]ANDERSSEN ERLEND L,DIEP D B,INGOLF F N,et al.Antagonistic Activity of Lactobacillus plantarum C11:Two New Two-Peptide Bacteriocins,Plantaricins EF and JK,and the Induction Factor Plantaricin A[J].Applied and Environmental Microbiology,1998,6: 2269-2272.
Antibacterial mechanism of plantaricin by Lactobacillus plantarum B-28
CHENG Jian-jun1,LI Xiang2,GUO Ming-ruo3
(1.Northeast Agricultural University,Harbin 150030,China;2.Nestle Shuangcheng Limited Company,Harbin 150100,China;3.University of Vermont,Burlington,VT 05403,USA)
The plantarcin was screened byLactobacillus plantarumB-28.The Bacillus cereus was the indicator bacteria.The antibacterial mechanism of the plantarcin was investigated through the changes of ATP level,ionic content and structure of cell.The results showed:As plantarcin which was screened byLactobacillus plantarumB-28 was added,intracellular ATP levels in energized cells of theBacillus cereusdropped quickly and then tended to be stable,however,extracellular ATP level increased during incubation in the present of plantaricin.The indictor’s cell lysis rate increased when the content of plantaricin was increased.After plantarcin was added,intracellar K+and Na+released quickly.It seemed there was a linear correlation between total amount of ions released and the concentration of plantaricin.Bacillus cereuswere treated by plantaricin,it was clearly seen that its cells wall appeared irregular at the edges by electron microscopy.At that time,their cells were cytomorphosis,cell walls were partially damaged,and intracellular materials were lost.
plantaricin;antibacterial;mechanism.
Q93-33
A
1001-2230(2011)05-0031-03
2011-02-21
程建军(1969-),男,博士,研究方向为农产品加工。
