新疆赛里木湖高白鲑性腺的组织学研究
2011-01-06曹玉洁齐亚银
曹玉洁,许 追,齐亚银,张 莉
(石河子大学动物科技学院,石河子832003)
新疆赛里木湖高白鲑性腺的组织学研究
曹玉洁,许 追,齐亚银,张 莉
(石河子大学动物科技学院,石河子832003)
通过光学显微镜对新疆赛里木湖高白鲑性腺的组织学结构进行研究。结果表明:高白鲑的精巢为小叶型,其精原细胞存在于精小叶内壁上,精母细胞和未成熟的精子细胞位于精小囊中,精子成熟后精小囊破裂释放精子进入小叶腔,完成发育过程,精小叶间存在大量的间质细胞、毛细血管和成纤维细胞。高白鲑卵巢是发育到第Ⅳ时期的卵巢,卵巢内存在不同发育时相的卵母细胞。
高白鲑;精巢;卵巢;组织学
高白鲑(Coregonus peled)属鲑科,白鲑属,为冷水性鱼类,自然分布于俄罗斯西伯利亚北纬50°以上的河流湖泊中,是俄罗斯重要的大面积养殖品种。高白鲑适应性强,生长速度快,肉味美,易捕捞,经济价值较高,又是典型的摄食浮游动物的鱼类。自1998年以来,新疆连续6年引进高白鲑发眼卵,经孵化后投放赛里木湖。高白鲑生殖器官成熟转化非常快,群体繁殖期非常短,这使其采卵的时机不易把握,因而人工繁殖时常会遇见难以同时解决功能性雄性和雌性亲鱼的问题,这给种苗生产造成很大困难[1]。吐尔逊等[2]对高白鲑的繁殖生物学特性进行了跟踪观测,对生殖器官外形发育的周年变化、繁殖力与体长体重的关系进行了详细的研究;张人铭等[3]在赛里木湖天然水域中直接捕捞高白鲑进行采卵和授精技术;蔡林钢等[4]指出高白鲑性腺的发育和成熟需要较低水温,雌鱼性腺发育到接近Ⅵ期末时,若水温能及时下降至0℃左右,则亲鱼能顺利产卵;反之,若水温不能及时下降,而保持3℃以上,则亲鱼产卵情况很差,如果保持较高水温,性腺将逐渐退化。目前,高白鲑的人工繁殖仍然处于摸索阶段,如何促进性腺发育成熟就成了生产上亟待解决的问题。因此,开展对高白鲑的生殖生物学的研究就显得非常迫切。本实验主要针对性成熟时期高白鲑的精巢和卵巢的显微结构进行观察,掌握该鱼的繁殖规律,以便在人工繁殖该鱼时,正确掌握催产的最佳时间,提高催产效果,旨在为克服目前的人工繁殖困难状况提供理论依据。
1 材料与方法
1.1 材料
2009年1月初,从新疆博州温泉县赛里木湖采集性成熟雌性和雄性高白鲑各10尾。
首先进行生物学测量并记录:雄性高白鲑全长37~40 c m,体长32~35 c m,头长5~7 c m,宽10.5~12 c m,总重568~654 g,然后解剖鱼体腹部,取出精巢,测量精巢大小并称重,计算其成熟系数;雌性高白鲑全长在38~41 c m,体长在33~34 c m,头长5~6 c m,宽10~12 c m,总重604~670 g,解剖取出卵巢,测量卵巢大小并称重,计算其成熟系数。
1.2 方法
将精巢和卵巢投入Bouin液中,固定12 h后修块,再投入Bouin液中固定12 h,共固定24 h,梯度乙醇脱水,二甲苯透明,石蜡包埋,切片6μm,APES粘片剂贴片,37℃烘片24 h,后经苏木精-伊红染色(HE染色),梯度乙醇脱水,二甲苯透明,中性树胶封片。在Oly mpus显微镜下观察并拍照。
2 结果与分析
2.1 精巢的外部形态
雄性高白鲑的精巢位于消化道的背方、鳔的腹面,左右各1条,以系膜与鳔相粘连,淡黑至乳白色,细长线状,中段稍粗,两端渐细(图1)。精巢长2.5~5.0 c m,最宽处直径0.25~0.30 c m。前段为生精部分,是精子发生的区域。后段为贮精囊,其长度约为生精部的1/3,在末端约1 c m的区域,向一侧长出薄膜状突起。高白鲑精巢平均成熟系数为14.5%。
2.2 精巢的组织学结构
光镜下观察性成熟个体精巢的纵切面和横切面,可以看到高白鲑同其它鲑形目鱼类一样,精巢分为外膜和实质两部分(图2)。外膜是一层结缔组织膜。实质部分由许多精小叶、小叶间质和输出管构成。
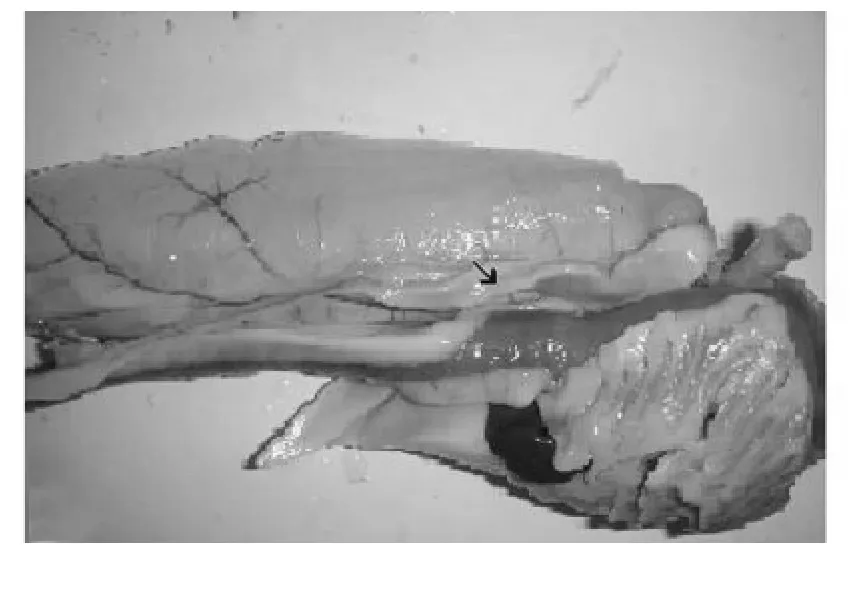
图1 高白鲑精巢外观Fig.1 The sper mary of Coregonus peled

图2 高白鲑精巢的横切面(10×10)Fig.2 The transverse section of sper mary in Coregonus peled.(10×10)
2.2.1 精小叶
精小叶呈管状,形状及分布不规则(图3)。在精小叶内壁分布有许多囊状结构,即精小囊。精小囊是支持细胞围成的囊状结构,形状不规则,除精原细胞外各期生精细胞均位于精小囊中。随着生精细胞的增殖和生长,精小囊的体积明显增大,精小囊壁则逐渐变薄。当精小囊中的生精细胞发育成熟,形成精子,精小囊就破裂,成熟的精子从精小囊中释放出来,进入小叶腔。图4可见到精小叶内缘的精原细胞呈圆形,体积较大,核大而清晰。生殖期高白鲑精巢中,精原细胞数量少,常位于小叶内靠近基膜的部位。精原细胞经有丝分裂后形成初级精母细胞,初级精母细胞呈圆球状,体积较精原细胞略小,核大且圆,嗜碱性较强,染色较深。大部分精小囊腔内有大量的精子细胞和成熟的精子。
2.2.2 小叶间质
精小叶之间由结缔组织分隔,称为小叶间质(图3)。小叶间质由纤维结缔组织、微血管和间质细胞构成。在血管外的间质组织中含有间质细胞、成纤维细胞和毛细血管等。间质细胞的细胞核形状不规则,强嗜碱性,细胞核较大。成纤维细胞胞体细长,细胞核为椭圆形。
2.2.3 小叶腔
小叶腔是精小叶管状结构中间的部分,Ⅳ期后期的精巢小叶腔变大,小叶壁变薄,生殖细胞减少,小叶边缘与中央的精子细胞群之间出现一环形空隙(图5)。Ⅴ期精子细胞经过变态成为成熟精子,体积小,被苏木素染成深蓝色,呈颗粒样,占据整个小叶腔。此时小叶腔变得更大更薄。
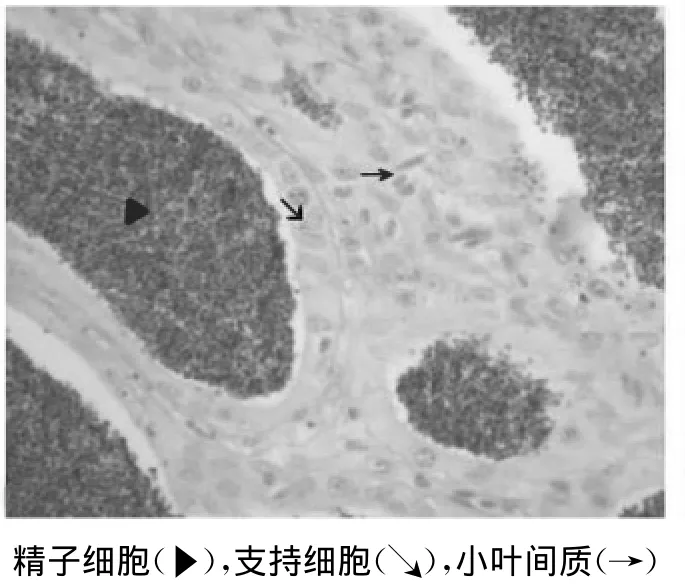
图3 精小囊及周围的小叶间质(40×10)Fig.3 Sper matogenic cyst and interlobular septum(40×10)
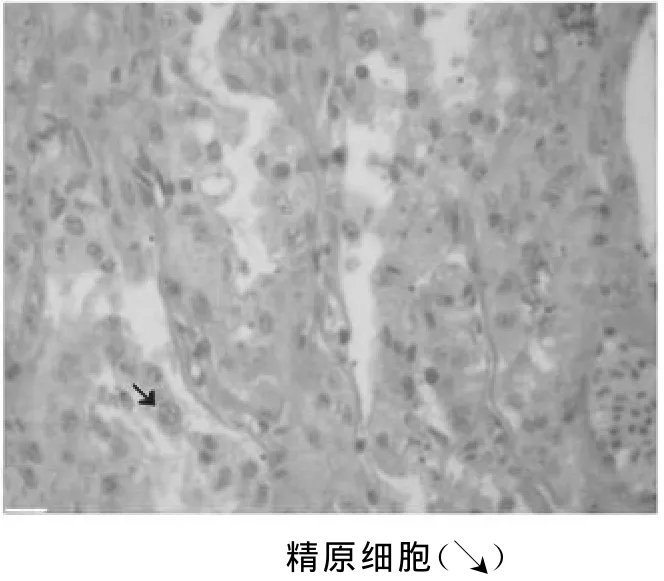
图4 精小叶生精细胞层(40×10)Fig.4 Sper matogenic cell layer of seminiferous lobula(40×10)
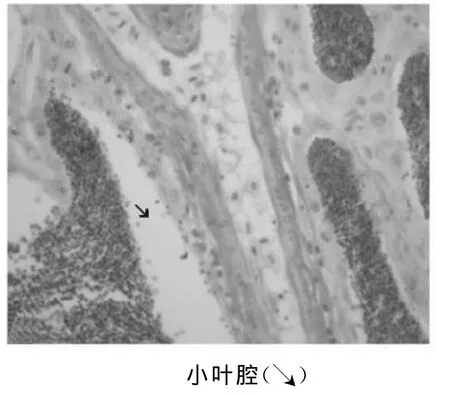
图5 精小叶及小叶腔(40×10)Fig.5 Seminiferous lobular and lobule lumen(40×10)
2.3 卵巢的外部形态
高白鲑的卵巢成对位于消化道背方,鳔上方的两侧,紧贴于肾脏腹面,外被卵巢膜。左右卵巢在腹腔后端相连,接口于一短的输卵管,呈“V”型,由泄殖孔开口于体外。高白鲑卵巢的平均成熟系数为16.7%。
2.4 卵巢的组织学结构
按照1939年Мейен提出的应用于一般硬骨鱼类中的标准,本实验所研究的高白鲑卵巢处于卵巢发育的第Ⅳ时期,卵巢内存在不同发育时期的卵母细胞以及少量成熟的卵粒。
第Ⅱ时相卵母细胞:处于初级卵母细胞的小生长期,大小不一,大多为不规则的多角圆形或椭圆形。细胞核较大,位于中央,占细胞体积的1/3~1/2,核膜明显,核仁较多,沿核膜内缘分布;细胞质为嗜碱性,H-E染色呈深蓝色,核呈浅紫色。第Ⅱ时相卵母细胞最明显的特征是胞质有明显的卵黄核,呈椭圆形,染色比胞质稍淡。卵母细胞外周有一薄层扁平滤泡细胞。第Ⅱ时相早期的卵母细胞呈圆形或椭圆形,胞质呈强嗜碱性,细胞质中出现着色较深的卵黄核(图6)。第Ⅱ时相晚期的卵母细胞,在细胞核外周出现围绕核的新月状的透明层。进一步发育,核膜局部破裂,部分核仁外排进入核周透明层中(图7)。

图6 第Ⅱ时相早期卵母细胞(↘)(40×10)Fig.6 The oocyte in the early phaseⅡ(↘)(40×10)
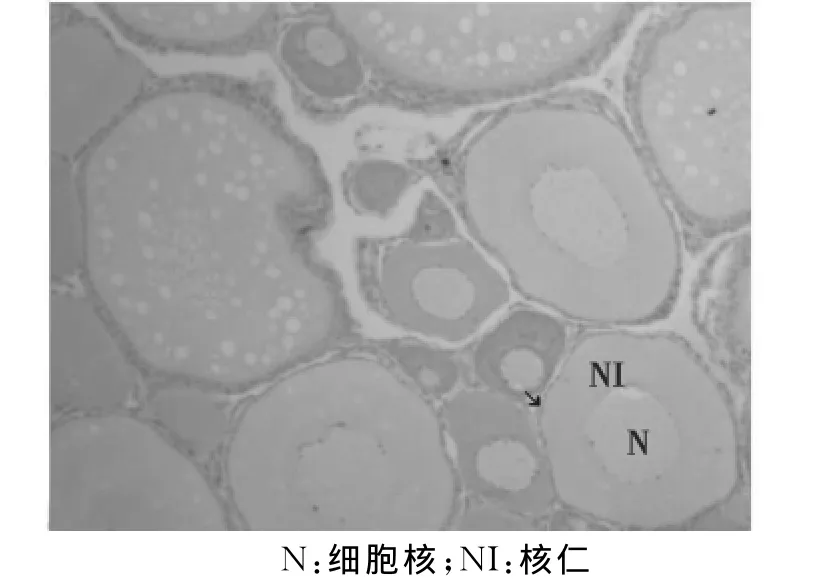
图7 第Ⅱ时相晚期卵母细胞(↘)(40×10)Fig.7 The oocyte in the late phaseⅡ(↘)(40×10)
第III时相卵母细胞:进入大生长期的初级卵母细胞,呈多角圆形,细胞和核的体积均进一步增大,细胞质为弱嗜碱性。在卵母细胞靠近卵膜周围的皮质层中出现2~4层大小不一的液泡,H-E染色不着色,在液泡之间有少量卵黄颗粒出现,滤泡细胞由单层发展为两层,同时在卵母细胞膜和滤泡细胞间出现一层非细胞结构的薄膜,称放射膜。第III时相早期的卵母细胞膜与滤泡细胞之间开始出现放射带,放射带外已有两层扁平的滤泡细胞,这时皮质层开始出现大小不等的液泡(图8)。随后液泡从皮层向中心移动,层数变为3~4层,卵母细胞体积也明显增大(图9)。
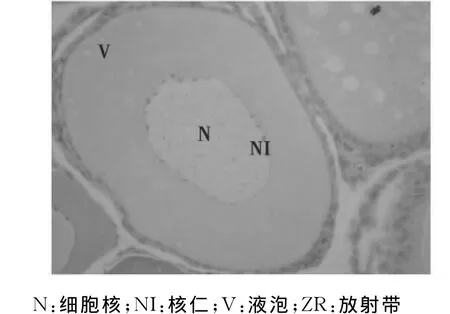
图8 第Ⅲ时相早期卵母细胞(40×10)Fig.8 The oocyte in the early phaseⅢ(40×10)

图9 第Ⅲ时相晚期卵母细胞(40×10)Fig.9 The oocyte in the late phaseⅢ(40×10)
第Ⅳ时相卵母细胞:此时相是初级卵母细胞卵黄颗粒充满的时期,其体积增大,多为椭圆形。此时,卵母细胞膜外的滤泡细胞仍为双层,放射膜增厚,清晰可见。第Ⅳ时相卵母细胞中,卵黄颗粒向内扩展至核膜的周围,几乎充满核外空间,只有在核的周围及靠近卵膜的边缘有较多的细胞质,为嗜碱性,液泡被挤压到卵母细胞周边的细胞质中。第Ⅳ时相早期卵母细胞胞质具有明显的分层现象,可分为内、中、外三层。内层,位于核膜外侧,无卵黄颗粒,液泡数量最多,大小不等,细胞质弱嗜碱性;中层,位于内、外层之间,含有大量的卵黄颗粒,其间夹有少量液泡;外层,即皮质部,含有少量液泡(图10)。第Ⅳ时相中期的卵母细胞几乎完全被卵黄颗粒所充满。卵黄颗粒可分为大、小两类,大卵黄颗粒多数呈圆球形或卵圆形(图11);此外,还可见一种棒状的卵黄颗粒(图12)。小卵黄颗粒呈圆球形,数量较多,不均匀的分布在大卵黄颗粒之间。放射带增厚呈强嗜酸性。到了晚期,胞质中开始出现油球,有的卵黄颗粒融合,呈均质红染,此时细胞核也开始由中央移向动物极,出现极化现象。
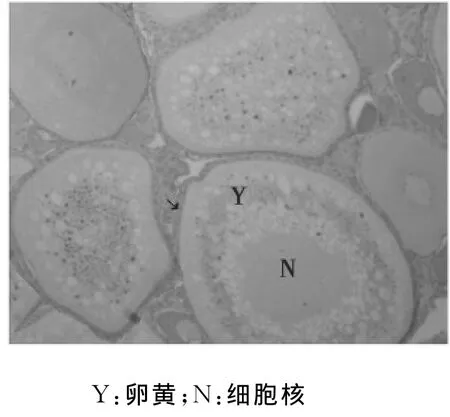
图10 第Ⅳ时相早期卵母细胞(↘)(40×10)Fig.10 The oocyte in the early phaseⅣ(↘)(40×10)

图11 卵黄颗粒呈圆球形或卵圆形(40×10)Fig.11 The yolk granules appeared spherical or oval(40×10)
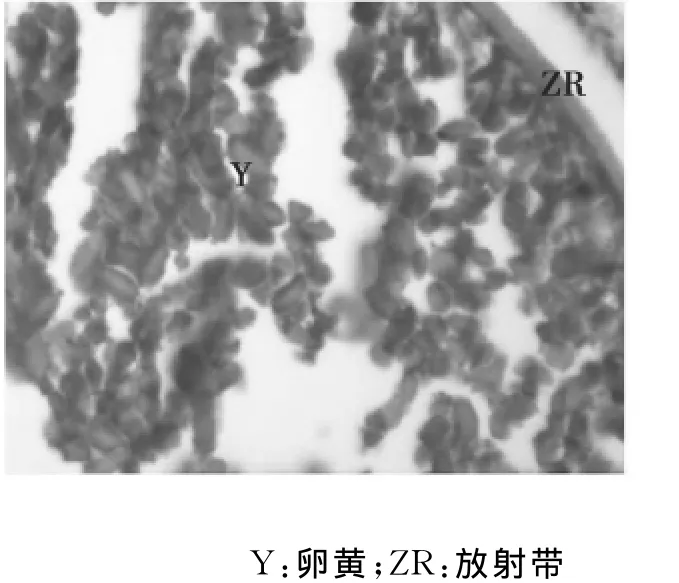
图12 卵黄颗粒呈棒状(40×10)Fig.12 The yolk granules appeared club-shaped(40×10)
第Ⅴ时相卵母细胞:这是由初级卵母细胞经过成熟分裂向次级卵母细胞过渡的阶段。细胞质中充满粗大的卵黄颗粒,卵黄颗粒大多数呈椭圆形,排列十分紧密。胞核已移动到动物极附近,核膜、核仁消失,仅见一团核质。
卵母细胞进入第2次成熟分裂中期(图13)。此时,卵细胞已生理成熟,与滤泡膜分离并游离于卵巢腔中。
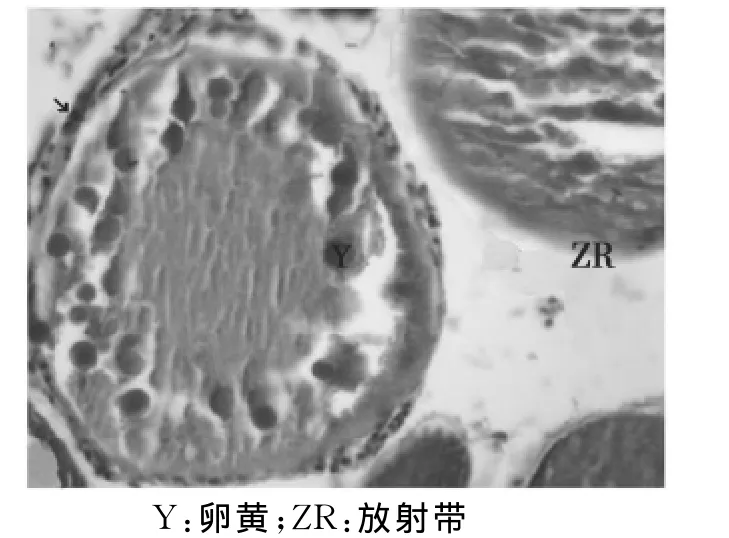
图13 第Ⅴ时相卵母细胞(↘)(40×10)Fig.13 The fifth period oocyte(↘)(40×10)
3 讨论
硬骨鱼类精巢的形态大多呈长囊状,且成对存在,但黄鳝(Monopterus albus)除外[5]。鲶形目鱼类如长吻鱼危(Leiocassis longirostris Günther)[6]、黄颡鱼(Pelteobagrus fulvidraco Richardson)[7]等的精巢则高度分支,南方鲇(Silur usmeridionalis Chen)[8]和革胡子鲇(Clarias leather)[9]的精巢也有少量较短的齿状突起。海鳗(Mureanesox cinereus)[10]精巢的形态在鱼类中较少见,由许多精巢叶片规则排列而成。中华乌塘鳢鱼精巢细长线状,生殖季节最粗时也只有3 mm。但贮精囊部分的膜状结构在生殖季节面积较大,大大增强了其分泌功能。一般硬骨鱼类精巢尾端合并,呈“Y”型。但高白鲑左右精巢在后端并不合并,而是分别开口于尿殖腔,此特点与金色小沙丁鱼(sardinella aurita)[11]相同。
关于硬骨鱼的精巢,一般根据生精细胞在精巢内分布的特点,分为小管型和小叶型。二者的主要区别在于小叶型精巢的精原细胞存在于小叶的边缘处,随着发育的进行,精原细胞和精母细胞形成小囊腔隙,精细胞和精子逐渐向小囊腔的中央移动;而小管型精巢没有壶腹结构,精原细胞只限于管的盲端[12]。切片显示,高白鲑的精巢与大多数硬骨鱼类的精巢相似,为小叶型,具有小叶型精巢的显著特点。
鱼类卵母细胞的成熟过程中有3种不同类型的卵黄,即皮质液泡(含碳水化合物卵黄)、蛋白卵黄颗粒和脂质卵黄滴[13]。高白鲑卵黄物质出现的次序是液泡-卵黄-油球,这与斑尾复虾虎鱼[14]、纹缟虾虎鱼[15]的卵黄出现次序一致。卵黄颗粒最先出现在胞质内靠近核膜处,然后逐渐向质膜方向移动,最后充满整个卵子,和鲻(Mugil cephalus)、圆口铜 鱼 (Coreius guichenoti)、长 江 刀 鲚 (Coilia ectenes)的卵黄颗粒出现方式不同[16-18]。鳞铲颌鱼(Varicorhinus macrolepis)和青海湖裸鲤(Gymnocypris przewalskii)的卵黄颗粒最先出现在卵母细胞核膜附近,并由内向周边扩展[19-20]。高白鲑第Ⅳ时相晚期卵母细胞开始出现一些小油滴,当卵接近成熟时,几个小油滴融合形成大的油球。
高白鲑样品于1月初采集,其中部分雌鱼轻压腹部有卵液流出,说明此时卵母细胞已经发育成熟;同时通过组织切片观察发现,大部分卵巢处于Ⅳ期,说明高白鲑卵巢的成熟期在1月或更晚,这与蔡林钢等[4]报道的繁殖旺季为每年的12月份或者第2年的1月初的结果相似。
[1]张人铭,蔡林钢,吐尔逊,等.赛里木湖高白鲑性腺发育观测[J].水产学杂志,2001,14(1):66-69.
[2]吐尔逊,蔡林钢,郭焱,等.赛里木湖高白鲑繁殖生物学特性研究[J].水产学杂志,2004,17(2):26-31.
[3]张人铭,蔡林钢,吐尔逊,等.赛里木湖高白鲑的人工繁殖[J].淡水渔业,2001,31(5):12-13.
[4]蔡林钢,吐尔逊,谢春刚,等.2008年度赛里木湖高白鲑人工采卵工作总结[J].新疆渔业科技,2008(2):18-21.
[5]张小雪,董元凯.黄鳝精巢显微和超微结构的研究[J].水利渔业,1994(4):9-11.
[6]张耀光,罗泉笙,钟明超.长吻鱼危精巢发育的分期及精子的发生和形成[J].动物学研究,1992,3(3):281-287.
[7]郑小真,林丹军,尤永隆.性成熟黄颡鱼精巢的年周期变化:Ⅰ.生精部的周年变化[J].福建师范大学学报:自然科学版,2000,16(2):97-101.
[8]张耀光,谢小军.南方鲇的繁殖生物学研究:性腺发育及周年变化[J].水生生物学报,1996,20(1):8-16.
[9]刘少军,姚占州.革胡子鲇成熟精巢超微结构的研究[J].湖南师范大学学报:自然科学学报,1992,15(3):252-256.
[10]张克俭,张饮江,郑东勇.海鳗性腺形态和发育的研究[J].水产学报,1999,23(1):13-20.
[11]王瑁,杨圣云,陈明茹.闽南台湾浅滩渔场金色小沙丁鱼精巢的发育[J].台湾海峡,2000,19(1):17-21.
[12]刘筠.中国养殖鱼类繁殖生理学[M].北京:农业出版社,1993:29-31.
[13]Mayer I,Shackley S E,Ryland J S.Aspect of the reproductive biology of the bass,Dicentrarchus l abax L.I.an histological and histochemical study of oocyte development[J].Fish Biol,1988,33:609-622.
[14]孙帼英,陈建国,吴引忠.斑尾复虾虎鱼的成熟与产卵[J].水产科技情报,1996,23(3):99-107.
[15]章龙珍,陈丽慧,庄平.长江口纹缟虾虎鱼繁殖季节的性腺组织学[J].水产学报,2009,33(2):245-252.
[16]方永强,林君卓,翁幼竹,等.池养鲻的卵巢发育和卵子发生过程[J].水产学报,2004,28(4):353-359.
[17]张贤芳,张耀光,甘光明,等.圆口铜鱼卵巢发育及卵子发生的初步研究[J].西南农业大学学报,2005,27(6):892-901.
[18]陈文银,李家乐,练青平.长江刀鲚性腺发育的组织学研究[J].水产学报,2006,30(6):773-777.
[19]唐洪玉,陈大庆,史建全,等.青海湖裸鲤性腺发育的组织学研究[J].水生生物学报,2006,30(2):166-172.
[20]宋卉,王树迎,彭克美,等.多鳞铲颌鱼性腺分化发育的组织学研究[J].中国水产科学,2006,13(5):723-729.
Gonad Histological Structure of Coregonus Peled in Sayram Lake in Xinjiang
CAO Yujie,XU Zhui,QI Yayin,ZHANG Li
(College of Animal Science and Technology,Shihezi University,Shihezi 832003,China)
Gonad histological structure of Coregonus peled in Xinjiang Sayram Lake was investigated by light microscope.The results indicated that the sper mar y of Coregonus peled belongs to the lobular type,its spermatogonium resides in the endothecia of seminiferous lobula,sper matocyte and immature spermatid located in the spermatogenic cysts.As sper matogenic cysts rupture and release the spermintolu men of seminiferous lobula after sper matozoons have matured,develop ment process of spermatozoon would be co mpleted.There are copious leydig cells,fibroblast cells and blood capillaries among seminiferous lobulas.The ovary of Coregonus peled belongs to the fourth period of ovarian development.The ovary has several periods of oocytes in ovarian development.
Coregonus peled;spermary;ovary;histology
S965.232
A
1007-7383(2011)05-0570-06
2011-06-21
曹玉洁(1986-),女,硕士生,专业方向为动物生理与生物化学;e-mail:1457839598@qq.co m。
张莉(1982-),女,副教授,从事动物生殖生物学研究;e-mail:zhangli@shzu.edu.cn。
