犁头鳅属鱼类物种地理分化过程
2010-12-25唐琼英杨金权刘焕章
唐琼英,俞 丹,杨金权,刘焕章,*
(1. 中国科学院水生生物研究所 水生生物多样性与保护重点实验室,湖北 武汉 430072;2. 上海海洋大学 鱼类研究室,上海 201306)
犁头鳅属鱼类物种地理分化过程
唐琼英1,俞 丹1,杨金权2,刘焕章1,*
(1. 中国科学院水生生物研究所 水生生物多样性与保护重点实验室,湖北 武汉 430072;2. 上海海洋大学 鱼类研究室,上海 201306)
犁头鳅属为中国的特有属,包括犁头鳅和长鳍犁头鳅两个物种。犁头鳅广泛分布于长江中上游,而长鳍犁头鳅则局限分布于闽江水系。该研究对采自长江中上游的犁头鳅20个个体和闽江水系的长鳍犁头鳅8个个体的线粒体细胞色素b(cytb)基因序列进行分析,以期对犁头鳅属鱼类的物种地理分化过程进行研究。结果显示,犁头鳅不同单倍型之间的遗传变异水平为0.1%~1.8%,平均为0.7%;长鳍犁头鳅各单倍型之间的遗传变异水平为0.6%~1.3%,平均为0.7%;犁头鳅和长鳍犁头鳅单倍型之间的遗传差异也较小,仅为0.2%~1.8%,平均为0.9%。采用邻接(NJ)法和贝叶斯(BI)法构建的分子系统发育树都一致显示,该研究中的犁头鳅属鱼类构成一个单系;所有长鳍犁头鳅样本构成一个单系,位于系统发育树的顶部位置;犁头鳅的样本不构成单系,而是形成并系。从形态上看,犁头鳅和长鳍犁头鳅均为有效种。由此推测,在第四纪冰期,长江中下游的犁头鳅沿东海大陆架向南扩散到东南沿海水系,长鳍犁头鳅可能是犁头鳅的一个种群扩散到闽江水系后,由于适应新的环境条件而分化成的一个新物种,剩下的长江流域的犁头鳅种群则构成一个并系类群。该文对于类似的地理物种形成方式也进行了讨论。
犁头鳅属;物种地理分化;东海大陆架;长江水系;闽江水系
物种分化是指一个物种产生分化而形成新物种的进化过程。物种分化是物种多样性形成的基本过程,是生物多样性起源的重要组成部分。对物种分化的研究是进化生物学最热门的研究课题之一。2009年是达尔文诞辰200周年和《物种起源》发表150周年纪念,因此,关于物种分化的研究再次成为最活跃最热门的研究课题。
引起物种分化的因素有很多,其中隔离是一个重要的影响因素。在物种分化过程中,隔离(包括地理隔离和生殖隔离)起着十分关键的作用。隔离是物种形成的一个极为重要的条件,隔离导致遗传物质交流中断,使群体歧化不断加深,直至新种形成。隔离(主要是环境隔离)既是物种形成的重要条件,隔离(主要是生殖隔离)同时又是物种形成的重要标志(Xia,2005)。关于物种的分化过程,Avise(2000)认为要经历复系、并系、单系三个阶段,即在进化初期由一个祖先分化出A种群和B种群,它们分别分化出很多分支。隔离的出现使这些分支随机分布在不同的生境,此时这两个种群间存在基因交流,相互构成复系关系;在后来的进化过程中,由于自然选择、瓶颈效应等各种因素的作用,种群B成为一个单系群,而种群A 则可能成为一个并系群。再后来,随着时间的推移,种群A也会进化为一个单系群。 这样A和B分别构成各自的单系群,成为两个分化完全的物种。根据这一假说,一个物种为单系,其祖先种为并系,这一状态成为物种处于早期分化阶段的一个标记。
研究物种分化过程的方法有很多,谱系生物地理学(phylogeography)是近年来随着分子生物学技术的发展而兴起的一门学科,它是采用分子系统发育分析的方法,探讨近缘物种或种内族群间的演化与地质历史事件之间的关系,分析种间或种内支系现有地理分布格局的形成过程(Avise, 2000; Zhang, 2003),进而探讨物种的分化机制。近年来,利用线粒体DNA(mtDNA)作为分子标记对动植物进行谱系生物地理学研究已成为热点,也取得了不少成果,尤其在淡水鱼类中。这是因为淡水鱼类的分布受水系的严格限制,地理隔离广泛存在。
犁头鳅属(Lepturichthys)隶属于鲤形目(Cypriniformes)鳅超科(Cobitoidea)平鳍鳅科(Balitoridae)平鳍鳅亚科(Balitorinae),是中国的特有属。Regan(1911)以Homaloptera fimbriataGünther为模式建立了新属——犁头鳅属(Lepturichthys),该属以具细长的尾柄而显著区别于Homaloptera属。后来,Hora (1932)认为采自四川岷江和湖南洞庭湖的犁头鳅标本不同于采自湖北宜昌的犁头鳅模式标本,并根据胸鳍不分支鳍条数目、背鳍最长鳍条的长度、全身是否具疣状刺突、体色体斑等特征分别将采自四川岷江、湖南洞庭湖的标本命名为两个新种:Lepturichthys guntheri和Lepturichthys nicholsi,但当时描述新种时采用的标本分别仅2尾。陈宜瑜(Chen, 1978)认为L. guntheri和L. nicholsi都应是犁头鳅L. fimbriata的同物异名。Dai(1985)描述了采自福建南平的一个新种——长鳍犁头鳅(Lepturichthys dolichopterus)。因此,到目前为止,犁头鳅属鱼类共包括两个有效种——犁头鳅和长鳍犁头鳅。其中,犁头鳅广泛分布于长江中上游,而长鳍犁头鳅则局限分布于闽江水系(Chen & Tang,2000)。
与大多数鲤科鱼类相比,犁头鳅属鱼类的分布范围较窄,两个种呈现间断分布,为典型的东洋区淡水鱼类类群。犁头鳅属鱼类目前的分布格局是如何形成的;属内两个物种的分化过程如何等问题的研究目前还未见报道。
快速进化的线粒体DNA(mtDNA)在进行淡水鱼类的谱系生物地理学研究时可以作为一个很好的分子标记。其中细胞色素b(cytb)基因为蛋白质编码基因,目前已广泛地运用于各淡水鱼类类群的物种分化研究(如:Perdice et al, 2004; Xia et al, 2005; Yang & He, 2008; Zhang et al, 2008; Yang et al, 2009)。本研究拟采用线粒体cytb基因对犁头鳅属鱼类的物种地理分化过程进行研究,讨论其现今分布格局的形成原因,以探讨物种分化的规律和机制。
1 材料与方法
1.1 研究材料
本研究共包括33个样本,其中采自长江中上游的犁头鳅共20个个体,闽江水系的长鳍犁头鳅共8个个体,并选择同属于平鳍鳅亚科的四川华吸鳅(Sinogastromyzon szechuanensis)、下司华吸鳅(S. hsiashiensis)、台湾间吸鳅(Hemimyzon formosanum)、台东间吸鳅(H. taitungensis)及广西华平鳅(Sinohomaloptera kwangsiens)作为外类群进行比较。实验所用标本均为95%酒精固定样本,保存于中国科学院水生生物研究所淡水鱼类博物馆。标本名称及采集地等详细信息见表1,犁头鳅属鱼类的采集地点如图1所示。
1.2 基因组DNA的提取、PCR扩增及测序
基因组DNA的提取采用高盐抽提法(Tang et al, 2008)。基因序列片段通过PCR扩增所得。扩增所用引物为L14724和H15915(Xiao et al, 2001)。PCR反应的总体积为60 µL,用大约50 ng的基因组DNA作为模板,反应体系包括:10×Buffer 6 µL,dNTPs0.75 µL(10 mmol/L),引物各1.5 µL(10 µmol/L),TaqDNA聚合酶1.5 U,加灭菌双蒸水至60 µL。PCR反应条件为:94℃ 预变性3 min;94℃ 变性45 s,52~58℃退火45 s,72℃延伸1 min,共35个循环;最后在72℃延伸8 min。将获得的PCR产物送基因公司进行纯化回收,采用扩增所用引物对回收产物进行测序。序列递交GenBank,获得的序列号见表1。
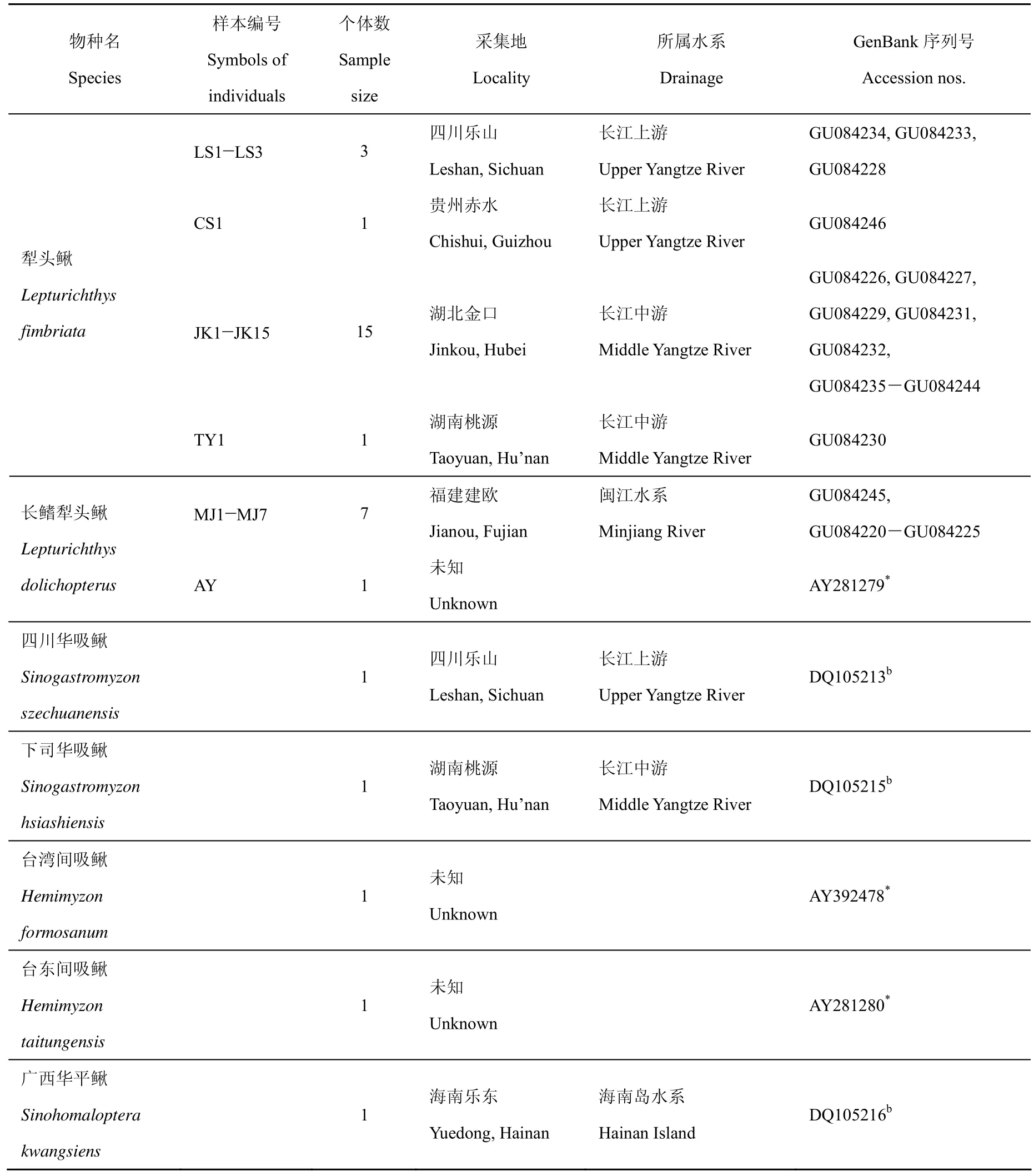
表 1 研究所用样本的物种名、编号、个体数、采集地及GenBank序列号等信息Tab. 1 Species, symbols, sample size, sample locations and GenBank accession numbers in the present study
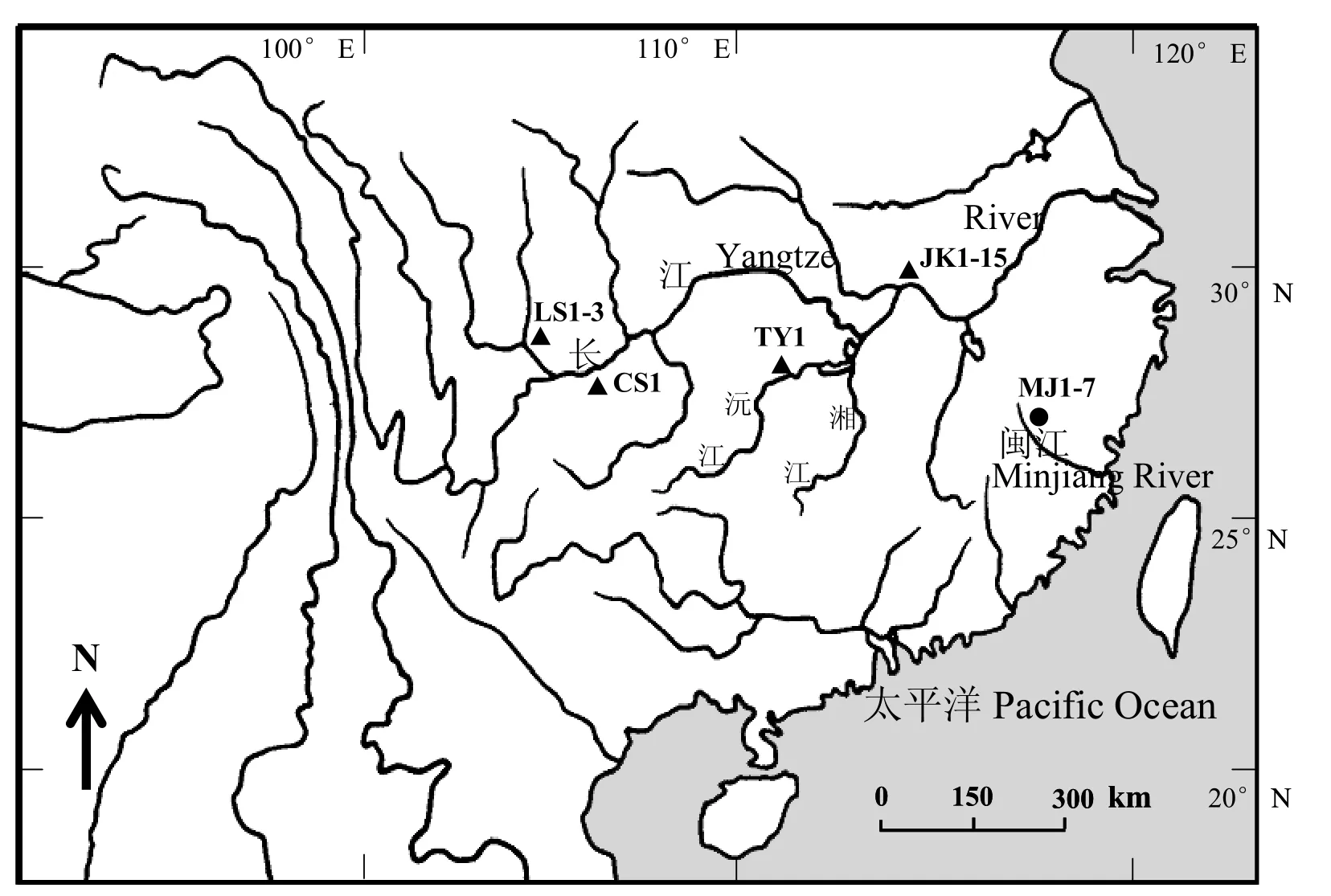
图 1 本研究中犁头鳅属鱼类的采集地点Fig. 1 Sample locations for the genus Lepturichthys in the present study
1.3 数据处理及分析
DNA序列的排列使用Clustal X软件,并在SEAVIEW程序中对序列进行手工调整。单倍型多样性和核苷酸多样性的统计采用DNAsp4.9软件(Rozas et al, 2003)。单倍型之间成对的遗传距离、各类群内及类群间的平均遗传差异情况在Mega4软件(Tamura et al, 2007)中采用Kimura双参数模型进行计算;序列中各碱基组成、碱基变异情况在PAUP软件中进行统计分析。序列的核苷酸替代饱和性分析是通过对转换(transitions, Ts)、颠换(transversions, Tv)的绝对数目和遗传距离进行作图分析(替代的绝对数目和遗传距离均在PAUP软件中进行统计)。分子系统树的构建采用邻接(neighbor joining, NJ)法和贝叶斯法(Bayesian analysis)。邻接法采用Mega4软件,各分支的置信度都采用1000次自展分析(Bootstrap analysis)进行重复检验。贝叶斯法采用MrBayes 3.0b软件(Huelsenbeck & Ronquist, 2001),各分支的置信度以后验概率(Posterior probability)来表示。
2 结 果
2.1 犁头鳅属鱼类cyt b基因序列分析
对测序获得的犁头鳅属鱼类cytb基因序列进行比对分析后,删去首尾测序信号较差的碱基,最终用于分析的cytb基因序列长为1 074 bp。20个犁头鳅个体中共检测到12个单倍型,单倍型多样性的平均值为0.937,核苷酸多样性的平均值为0.00 468;8个长鳍犁头鳅个体中有4个单倍型,单倍型多样性的平均值为0.643,核苷酸多样性的平均值为0.00349。犁头鳅属鱼类单倍型的分布情况见表2。犁头鳅样本中,采自四川乐山与湖北金口的个体存在共享单倍型(LS2和JK2共享单被型Hap1,LS1、LS3、JK8及JK11共享单倍型Hap4),采自湖南桃源的个体与湖北金口的个体存在共享单倍型(TY1和JK14共享单倍型Hap3),湖北金口的个体间也存在共享单倍型(Hap7, Hap9),剩下的7个单倍型为独有,其中1个为贵州赤水的个体拥有,其它6个为湖北金口的个体拥有。8个长鳍犁头鳅样本中,有5个个体共享一个单倍型hap1,其余3个个体拥有独有的单倍型。
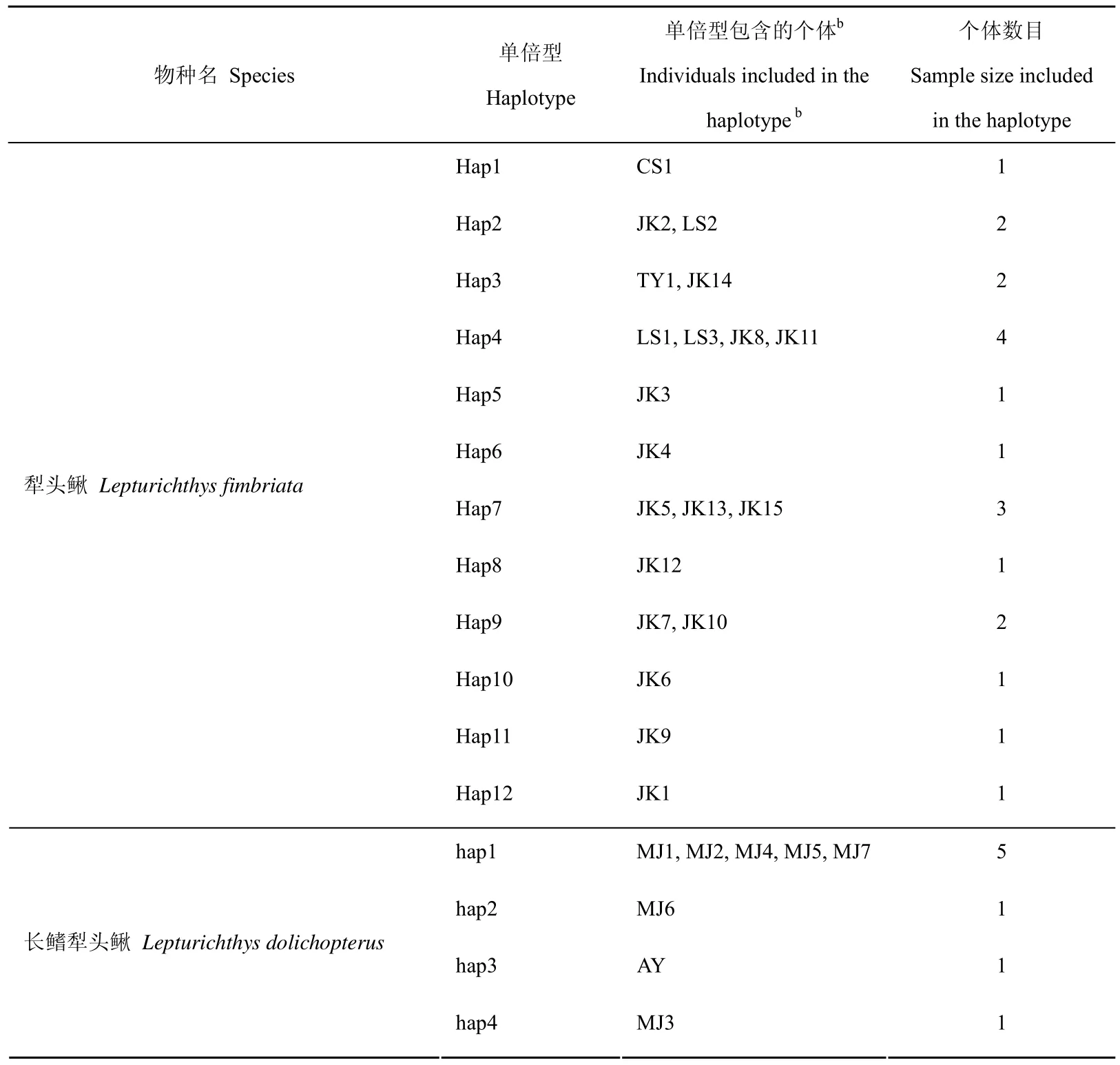
表 2 犁头鳅属鱼类单倍型的分布情况Tab. 2 The information of haplotypes of the genus Lepturichthys species
对本研究中获得的所有单倍型及外类群进行序列分析,结果显示,该基因序列片段中A、C、G、T的平均含量分别为28.0%、29.6%、14.4%和28.0%,A+T的含量(56.0%)明显高于C+G的含量(44.0%),同时表现出强烈的低G碱基含量的碱基组成偏向性,尤其在第三位点表现更为明显,G的含量仅为3.4%。所有序列转换颠换比(Ts/Tv)的平均值为5.21。在1074个位点中,变异位点有217个,其中简约信息位点有108个。序列中核苷酸替代的饱和性分析结果表明,转换和颠换均未达饱和(图2),因此,所有碱基均可用于系统发育分析。
基于Kimura双参数模型计算的犁头鳅属鱼类单倍型之间的遗传变异情况表明,犁头鳅不同单倍型之间的遗传变异水平为0.1%~1.8%,平均为0.7%,长鳍犁头鳅各单倍型之间的遗传变异水平为0.6%~1.3%,平均为0.7%,犁头鳅和长鳍犁头鳅单倍型之间的遗传差异也较小,仅为0.2%~1.8%,平均为0.9%,这和犁头鳅及长鳍犁头鳅种内单倍型之间的差异类似。犁头鳅属鱼类和外类群华吸鳅属、间吸鳅属及华平鳅间的平均差异较大,分别为5.4%、8.9%和9.5%。
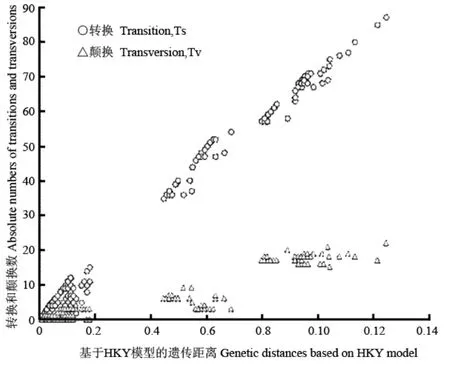
图 2 Cyt b基因序列中核苷酸替代的饱和性分析图Fig. 2 Plot of transition and transversion substitutions versus the HKY distance for all positions of the cytochrome b gene sequences
2.2 分子系统发育分析
采用邻接法和贝叶斯法构建的犁头鳅属鱼类单倍型之间的系统发育关系如图3。邻接(NJ)树和贝叶斯(BI)树的拓扑结构大致类似,只在单倍型的分支顺序上存在差异。这两棵系统发育树都一致显示,该研究中的犁头鳅属鱼类构成一个单系,且得到较强的支持(NJ树中自展值为100,BI树中的后验概率为1.00)。分布于闽江水系的长鳍犁头鳅构成一个单系,位于系统发育树的顶部位置,但在NJ树中支持率不高,仅为49,在BI树中支持率为1.00。所有犁头鳅的样本不构成单系,而是形成并系,其中采自湖北金口的两个个体(JK9和JK1)位于犁头鳅属鱼类的最基部位置,其它犁头鳅个体位于系统发育树的中间位置,犁头鳅的这种系统发育关系在NJ树和BI树中几乎是一致的。
3 讨 论
3.1 犁头鳅属鱼类的物种有效性
本研究结果显示,犁头鳅属鱼类中犁头鳅和长鳍犁头鳅这两个物种间的遗传差异较小,几乎和物种内不同个体间的遗传变异相当。分子系统发育分析的结果表明,长鳍犁头鳅的个体可以形成单系,但犁头鳅的样本不能聚为单系,犁头鳅的部分样本与长鳍犁头鳅具有更近的亲缘关系。这两个物种是否均为有效种,仅用遗传差异的大小是不能界定物种的,因为不同类群的物种间及不同基因间遗传差异变异的范围很大。另外,物种的划分也不能简单地以分支关系来处理(Xia et al, 2005)。长鳍犁头鳅和犁头鳅在形态上存在显著的区别特征:长鳍犁头鳅偶鳍均较长,体长为胸鳍长的4.1~4.8倍(VS. 6.5~7.9倍),为腹鳍长的4.8~5.7倍(VS. 6.5~8.3倍);腹鳍末端接近或稍为超过肛门(VS. 腹鳍末端伸至腹鳍腋部至臀鳍起点间的中点);肛门约在腹鳍腋部至臀鳍起点间的2/3稍后(VS.约在腹鳍腋部至臀鳍起点间的2/3处)(Chen & Tang, 2000)。另外,我们对所研究样本的体色体斑进行了检视,发现长鳍犁头鳅样本较为单一,而犁头鳅样本变异较大。长鳍犁头鳅样本的体色均为棕黑色,横跨体背部具7~8个椭圆形的大斑块,被窄小的浅黄色条纹隔开;犁头鳅样本的体色从浅黄色到棕黑色,背部斑纹有的为褐色的椭圆形斑块,有的呈圆形眼斑,有的为不规则斑纹,有的斑纹不明显。这两个物种在形态上存在显著的区别特征,分布区也不重叠,因此我们认为这两个物种均为有效种。目前的分子数据表明,这两个物种间的遗传差异极小,我们推测,长鳍犁头鳅可能是新近分化出来的一个物种。环境的改变导致犁头鳅的一个地理种群在形态上发生迅速改变,但在遗传上还没有足够长的时间来积累其遗传差异。鱼类中还有很多在形态上存在差异但在遗传上差异却不明显的例子,尤其在非洲湖泊的丽鱼中这种现象更为普遍,尽管这些鱼类之间的遗传差异较小,但都是有效种(Meyer et al, 1990; Wilson et al, 2000)。
3.2 犁头鳅属鱼类的物种分化过程
犁头鳅广泛分布于长江中上游水域,之前所记录的分布范围是长江水系的湖南洞庭湖至四川境内。郭治之和刘瑞兰(Guo & Liu, 1995)、张鹗等(Zhang et al, 1996)发现在江西鄱阳湖水系的信江中也有犁头鳅的分布。长鳍犁头鳅局限分布于福建闽江水系。信江发源于浙赣两省交界的怀玉山南的玉山水和武夷山北麓的丰溪水,闽江水系发源于武夷山东南。因此,武夷山是长江中下游鄱阳湖水系与闽江水系的天然分水岭。本研究中犁头鳅和长鳍犁头鳅单倍型之间的遗传差异为0.2%~1.8%,按cytb基因在淡水鱼类中进化速率为每百万年1%计算(Durand et al, 2002),犁头鳅和长鳍犁头鳅发生分歧的时间在0.9~0.1百万年前,属于更新世。根据犁头鳅属鱼类目前的分布模式,其分布格局的形成有两种可能的途径:一种途径为信江和闽江曾经相连接,犁头鳅属鱼类曾经广布,武夷山的隆升才造成它们的隔离;另一种途径为,第四纪冰期海平面下降后,长江流域的犁头鳅顺长江口的水系进入闽江。由于犁头鳅和长鳍犁头鳅的遗传分化很小,说明其隔离的时间很晚。武夷山的隆升相对久远,造成的隔离应该比较大(Liu, 1984),第四纪冰期的发生则较为晚近。因此,对于犁头鳅属鱼类的分布,第四纪冰期的扩散更有可能。
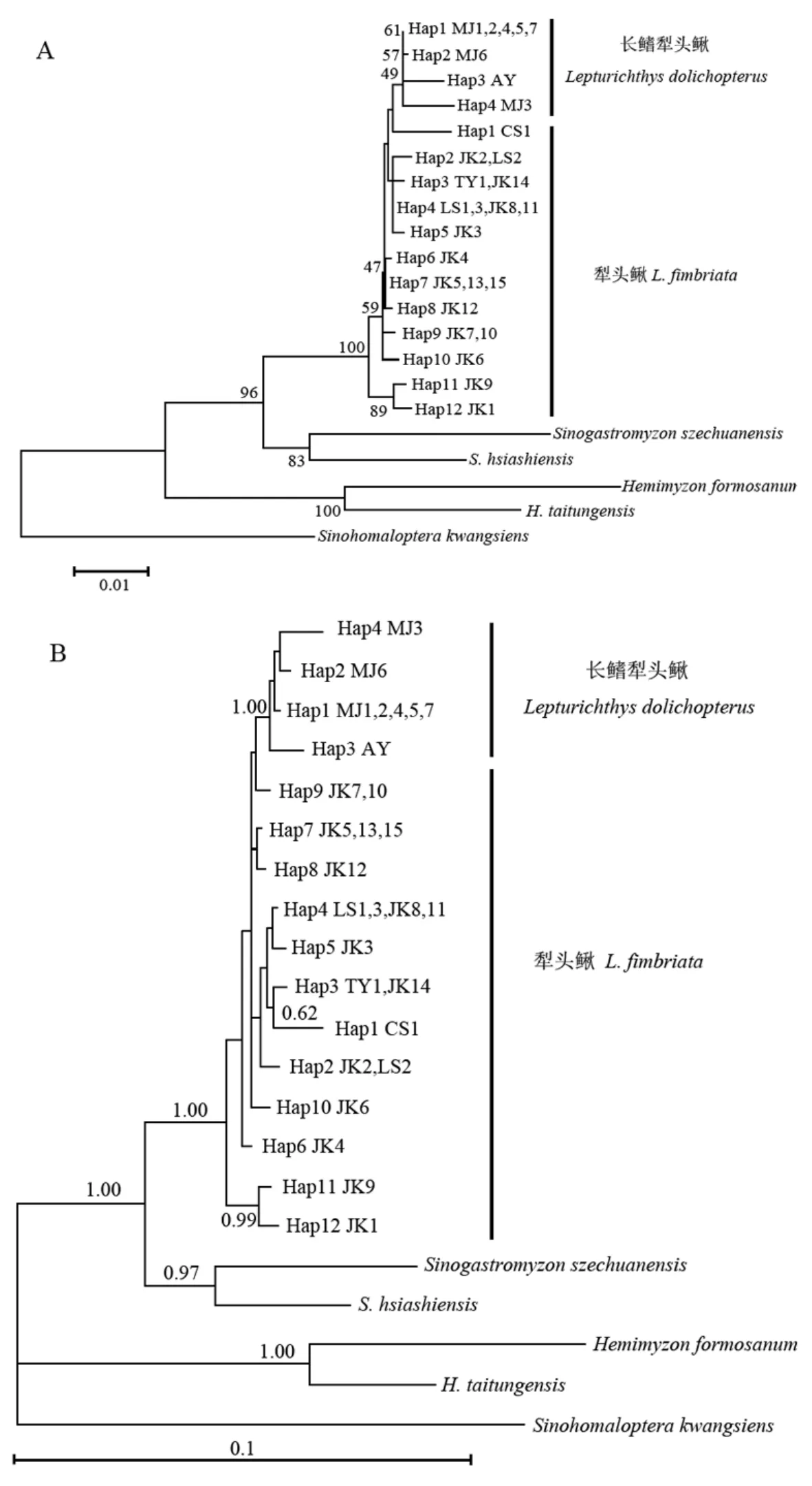
图 3 基于cyt b基因构建的犁头鳅属鱼类单倍型间的分子系统发育关系Fig. 3 The molecular phylogenetic trees of the haplotypes identified from the genus Lepturichthys species
综合本文的研究结果以及历史资料,本文对犁头鳅属鱼类的物种分化过程作如下推测:青藏高原的隆升及东亚季风气候的形成,使犁头鳅属这类适应激流环境的鱼类衍生出来,广泛分布于长江水系。在第四纪冰期全球气温下降,海平面降低,较现在下降超过100 m,东海大陆架露出海面成为陆地,部分水系可能相互贯通(Li & Fang, 1990)。分布于长江中下游的犁头鳅逐渐扩散至东南沿海各水系。在间冰期,全球气候变暖,海平面上升,大陆架被海水淹没,各沿海水系被隔离。犁头鳅属于产漂流性卵的鱼类,在产卵场的下游需要有足够长的河道才能满足受精卵漂流孵化的时间要求。因此,犁头鳅只在少部分水系中被保留下来,如闽江水系。由于闽江水系环境的差异,扩散到闽江水系的犁头鳅快速地发生一些分化以适应新的环境,从而分化形成长鳍犁头鳅。相对说来,长鳍犁头鳅表现出较低的遗传多样性,而分布广泛的犁头鳅则具有更为丰富的遗传多样性,且形成一个并系。
在本研究中,我们推测犁头鳅的扩散路径是从长江中下游经沿海大陆架向南进行扩散的。采用沿海大陆架作为迁移路径的例子在其他动物中也有报道,如长江下游四大家鱼向珠江流域、海南岛等地的扩散(Li & Fang, 1990),淮河过渡动物群的南迁(Cai, 2002)等。在第四纪大冰期期间,各物种向南迁移到“避难所”的现象是较为普遍的,而沿海大陆架是物种向南迁移的一个主要路径。
关于物种分化过程,Avise (2000)认为一般需要经历从复系到并系再到单系3个发展阶段。Zhang et al (2008)研究了采自长江的华鳈(Sarcocheilichthys sinensis sinensis) 和闽江的福建华鳈 (Sarcocheilich-thys sinensis fukiensis)这两个亚种的物种分化,发现这两个亚种都不能分别构成单系,而是相互混杂聚在一起形成复系。因此,推测这两个亚种可能还处于物种分化的早期阶段,这也正支持了两者的亚种地位。另外,Zhang et al (2008)对克氏鳈(Sarcocheilichthys czerskii)和黑鳍鳈(S. nigripinnis)的物种分化过程也进行了研究,发现黑鳍鳈的一个北方种群与克氏鳈有更近的谱系关系。由此他们推测,分布于黑龙江水系的克氏鳈可能是黑鳍鳈北方种群的一个分支向中国东北扩散后分化出来的一个物种,剩下的黑鳍鳈则形成一个并系类群。Shields et al(2000)指出,北极熊由棕熊的一个种群分化成种后,剩下的棕熊种群也是形成一个并系类群。
本研究中长鳍犁头鳅可以构成一个单系,但犁头鳅为一并系。犁头鳅和长鳍犁头鳅的物种分化过程与克氏鳈和黑鳍鳈及北极熊和棕熊的物种分化过程极为相似,即一个地理种群为适应新的环境而演化成另一个物种,从而使得原来物种的其他地理种群形成一个并系,它们在遗传上还没有完全分化成两个独立的物种,还处于物种分化的并系阶段。而上面所提及的华鳈和福建华鳈则还处于复系这一物种分化的起始阶段。因此,本文的研究结果进一步证实和丰富了物种形成的地理隔离过程假说。
致谢:感谢浙江大学的周卓诚先生、武汉中国地质大学的黄伟先生、中科院水生生物研究所的曾燏博士和熊玉宇博士在收集样本上给予的帮助。另外,本所的刘思情和周健同学在实验上也提供了大力支持,在此一并表示衷心的感谢。
Avise JC. 2000. Phylogeography, the History and Formation of Species [M]. England: Harvard University.
Cai BQ. 2002. The evidence of dry land of the Taiwan straits during late Würm glaciation [J].Mar Sci, 26(6): 51-54. [蔡保全. 2002. 晚玉木冰期台湾海峡成陆的证据. 海洋科学, 26(6): 51-54.]
Chen YY, Tang WQ. 2000. Homalopteridae [M]//Yue PQ. Fauna Sinica. Osteichthyes. Cypriniformes III. Beijing: Sciences Press, 438-567. [陈宜瑜, 唐文乔. 2000. 平鳍鳅科 [M]// 乐佩琦. 中国动物志(硬骨鱼纲·鲤形目·下卷). 北京: 科学技术出版社, 438-567.]
Chen YY. 1978. Systematic studies on the Fishes of the family Homalopteridae of China I. classification of the fishes of the subfamily Homalopterinae [J].Acta Hydrobiol Sin, 6(3): 331-348. [陈宜瑜. 1978.中国平鳍鳅科鱼类系统分类的研究I. 平鳍鳅亚科鱼类的分类. 水生生物学集刊, 6 (3): 331-348.]
Dai DY. 1985. A new species of the genusLepturichthysfrom China (Cypriniformes: Homalopteridae) [J].Acta Zootax Sin, 10 (2): 221-224. [戴定远. 1985. 中国犁头鳅属一新种. 动物分类学报, 10 (2): 221-224.]
Durand J, Tsigenopoulos C, Ünlü E, Berrebi P. 2002. Phylogeny and biogeography of the family Cyprinidae in the Middle East inferred from cytochromebDNA-evolutionary significance of this region [J].Mol Phylogenet Evol, 22: 91-100.
Guo ZZ, Liu RL. 1995. The fishes of Jiangxi Province [J].J Nanchang Univ:Nat Sci, 19(3): 222-232. [郭治之, 刘瑞兰. 1995. 江西鱼类的研究.南昌大学学报: 理科版, 19(3): 222-232.]
Huelsenbeck JP, Ronquist F. 2001. MrBayes: Bayesian inference of phylogeny [J].Bioinformatics, 17: 754-755.
Li SZ, Fang F. 1990. On the geographical distribution of the four kinds of pond-cultured carps in China [J].Acta Zool Sin, 36(3): 244-250. [李思忠, 方 芳. 1990. 鲢、鳙、青、草鱼地理分布的研究. 动物学报, 36(3): 244-250.]
Liu ZZ. 1984. The origin and morphological characteristics of the Wuyi Mountain, Fujian Province [J].J Nanjing Univ:Nat Sci, 3: 567-576. [刘振中. 1984. 武夷山的形成与地貌发育特征. 南京大学学报: 自然科学版, 3: 567-576.]
Meyer A, Kocher TD, Basasibwaki P, Wilson AC. 1990. Monophyletic origin of Lake Victoria cichlid fishes suggested by mitochondrial DNA sequences [J].Nature, 347: 550-553.
Perdices A, Cunha C, Coelho MM. 2004. Phylogenetic structure ofZacco platypus(Teleostei, Cyprinidae) populations on the upper and middle Chang Jiang (= Yangtze) drainage inferred from cytochromebsequences [J].Mol Phylogenet Evol, 31(1): 192-203.
Rozas J, Sanchez-DelBarrio JC, Messeguer X. 2003. DNASP, DNA polymorphism analyses by the coalescent and other methods [J].Bioinformatics, 19: 2496-2497.
Shields GF, Adams D, Garner G, Labelle M, Pietsch J, Ramsay M, Schwartz C, Titus K, Williamson S. 2000. Phylogeography of mitochondrial DNA variation in brown bears and polar bears [J].Mol Phylogent Evol, 15 (2): 319-326.
Tamura K, Dudley J, Nei M, Kumar S. 2007. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0 [J].Mol Biol Evol, 24: 1596-1599.
Tang Q, Freyhof J, Xiong B, Liu H. 2008. Multiple invasions of Europe by East Asian cobitid loaches (Teleostei: Cobitidae) [J].Hydrobiologia, 605: 17-28.
Tang Q, Liu H, Mayden RL, Xiong B. 2006. Comparasion of evolution rates in the mitochondrial DNA cytochromebgene and control region and their implications for the phylogeny of the Cobitoidea [J].Mol Phylogent Evol, 39: 347-357.
Wilson AB, Noack-Kunnmannn K, Meyer A. 2000. Incipient speciation in sympatric Nicaraguan crater lake cichlid fished: sexual selection versus ecological diversification [J].Proc R Soc Lond B, 267: 2133-2141.
Xia X, Zhang X, Liu H. 2005. Molecular biogeography ofPseudogobio vaillanti(Cypriniformes, Cyprinidae, Gobioninae) [J].Prog Nat Sci, 15(9): 1064-1069. [夏曦中, 张训蒲, 刘焕章. 2000. 似鮈的分子生物地理学分析. 自然科学进展, 15(9): 1064-1069.]
Xia XZ. 2005. Phylogeography and speciation of generaRhinogobioandPseudogobioin China [D]. Master’s Degree Dissertation, Huazhong Agricultural University, Wuhan. [夏曦中. 2005. 吻鮈属和似鮈属鱼类物种分化的比较. 硕士学位论文, 华中农业大学, 武汉.]
Xiao WH, Zhang YP, Liu HZ. 2001. Molecular systematics of Xenocyprinae (Teleostei: Cyprinidae): Taxonomy, biogeography, and coevolution of a special group restricted in East Asia [J].Mol Phylogenet Evol, 18: 163-173.
Yang L, He S. 2008. Phylogeography of the freshwater catfishHemibagrus guttatus(Siluriformes, Bagridae): Implications for South China biogeography and influence of sea-level changes [J].Mol Phylogenet Evol, 49: 393-398.
Yang L, Mayden RL, He S. 2009. Population genetic structure and geographical differentiation of the Chinese catfishHemibagrus macropterus(Siluriformes, Bagride): evidence for altered drainage patterns [J].Mol Phylogenet Evol, 51: 405-411.
Zhang DX. 2003. Advances in historical biogeography: a brief perspective [J].J Trop Subtrop Bot, 11(3): 283-289. [张奠湘. 2003. 历史生物地理学的进展. 热带亚热带植物学报, 11(3): 283-289.]
Zhang E, Liu HZ, He CC. 1996. Fish fauna in northeastern Jiangxi Province [J].Chn J Zool, 31(6): 3-12. [张 鹗, 刘焕章, 何长才. 2003. 赣东北地区鱼类区系的研究. 动物学杂志, 31(6): 3-12.]
Zhang L, Tang QY, Liu HZ. 2008. Phylogeny and speciation process of the eastern Asian cyprinid genusSarcocheilichthys[J].J Fish Biol, 72: 1122-1137.
Geographical Speciation Process of the GenusLepturichthys(Cypriniformes: Balitoridae)
TANG Qiong-Ying1, YU Dan1, YANG Jin-Quan2, LIU Huan-Zhang1,*
(1.The Key Laboratory of Aquatic Biodiversity and Conservation, Institute of Hydrobiology, the Chinese Academy of Sciences, Wuhan430072,China; 2.Laboratory of Fishes, Shanghai Ocean University, Shanghai200090,China)
The genusLepturichthysis endemic in China, including two valid species,L. fimbriataandL. dolichopterus. The former is widely distributed in the Upper and Middle Yangtze River, while the latter only in Minjiang River. In the present study, the mitochondrial cytochromebgene sequences were analyzed for 20 individuals ofL. fimbriatacollected from the Upper and Middle Yangtze River and eight individuals ofL. dolichopteruscollected from Minjiang River. This paper aimed to investigate the geographical speciation process of the genusLepturichthys. The results showed that the genetic variations among different haplotypes ofL. fimbriataranged from 0.1% to 1.8%, with an average of 0.7%, and from 0.6% to 1.3%, with an average of 0.7% among different haplotypes ofL. dolichopterus. The variations betweenL. fimbriataandL. dolichopteruswere also very low, ranging from 0.2% to 1.8%, with an average of 0.9%. Two methods, Neighbor Joining (NJ) and Bayesian analysis (BI), were used for constructing the molecular phylogenetic trees. Both NJ and BI trees indicated that allLepturichthysspecimens formed as a monophyletic group; theL. dolichopterussamples also formed as monophyletic group, having a derived position in the phylogenetic trees, while theL. fimbriatasamples formed as a paraphyletic group. BothL. fimbriataandL. dolichopterusare valid species in morphology. Based on molecular and morphological data, we presumed that the geographical speciation process of the genusLepturichthyscould be as following: during the Quaternary Glacial period,L. fimbriatain the Middle and Lower Yangtze River dispersed to the drainages of Southeast China along the Continental Shelf of East China Sea, one population spread to the Minjiang River and envolved as a new species (L. dolichopterus) because of considerable environmental difference, while the remainingL. fimbriatapopulations in the Yangtze River formed as a paraphyly. The other similar geographical speciation patterns are discussed in the present study as well.
book=3,ebook=136
Lepturichthys; Geographical speciation; Continental shelf of East China Sea; Yangtze River; Minjiang River
Q959.483; Q349
A
0254-5853-(2010)03-0230-09
10.3724/SP.J.1141.2010.03230
2009-10-10;接受日期:2010-03-01
国家自然科学基金资助项目(30700072;40432003);美国自然科学基金资助项目CToL∗
Corresponding author),Tel:027-68780776,E-mail:hzliu@ihb.ac.cn
