滇南勐腊地区威氏小鼷鹿种群生境利用
2010-12-25陈明勇
曹 明,周 伟,*,白 冰,张 庆,王 斌,陈明勇
(1. 西南林业大学 保护生物学学院, 云南省森林灾害预警与控制重点实验室, 云南 昆明 650224; 2. 西双版纳国家级自然保护区管理局,云南 景洪 666100)
滇南勐腊地区威氏小鼷鹿种群生境利用
曹 明1,周 伟1,*,白 冰1,张 庆1,王 斌2,陈明勇2
(1.西南林业大学 保护生物学学院,云南省森林灾害预警与控制重点实验室,云南 昆明650224; 2.西双版纳国家级自然保护区管理局,云南 景洪666100)
以样线与样方相结合,2007年6—12月在滇南勐腊地区的勐满和龙门片收集威氏小鼷鹿(Tragulus williamsoni)足迹链坐标位点,用GIS软件生成威氏小鼷鹿的分布格局图。在样方中测量11个生态因子。结果显示,威氏小鼷鹿对低海拔的河谷浅滩地带有明显的选择偏好,主要聚集在勐满片的三岔河地区和龙门片的大榕树地区。比较利用和可利用样方的定性因子,分析结果表明,威氏小鼷鹿偏好在中、下坡位活动,对粗叶榕(Ficus hirta)、木奶果(Baccaurea samiflora)、长柄山姜(Alpinia kwangsiensis)和柊叶(Phrynium capitatum)等植物有选择偏好。这些植物多沿河谷的两岸集中分布,是影响威氏小鼷鹿分布格局的重要原因之一。比较利用样方和可利用样方的定量因子,分析结果表明,乔木盖度、灌木盖度、落叶层盖度和距水源距离等4个因子差异显著,并且是区分利用样方和可利用样方的关键因子,也是影响小鼷鹿生境利用的主要因子。高密度的灌木和高大乔木形成双重的隐蔽条件,大大提高了安全性。由于距水源较近,乔木层及林下灌层生长状况较高海拔地区更为茂密,这是小鼷鹿在勐腊地区形成沿海拔较低的河岸集中分布格局的重要原因之一。
威氏小鼷鹿;生境利用;分布格局;云南南部
由于鼷鹿类为夜行性动物,常在茂密的灌木丛中活动,难以发现,生态学研究大都局限于笼养状况(Barrette, 1987; Dubost 1975, 1978; Nowak, 1991)。虽然笼养状态观察到的鼷鹿类习性不能完全等同于野生状态的习性,但对研究野生鼷鹿类的习性提供了参考和借鉴。对自然栖息地小鼷鹿研究较有影响的一项是对马来半岛的爪哇小鼷鹿(Tragulus javanicus)密度、种群大小的研究和自动摄像记录(Miura et al, 1997; Miura & Idris, 1999);另一项是通过佩带项圈,以无线电跟踪方法,调查婆罗洲Kabili-Sepilok保护区内野生爪哇小鼷鹿的家域及生境利用(Matsubayashi et al, 2003)。研究结果显示,在白天爪哇小鼷鹿从一隐蔽物向另一隐蔽物移动和在林下觅食;如果白天静栖的话,则是在隐蔽物下;夜晚静息时间比白天长,且在森林的开阔区;雌性的日活动范围和家域均大于雄性的。过去一直认为爪哇小鼷鹿是夜行性的,但结果大相径庭。
Peng et al (1962) 首次在云南省西双版纳的勐腊地区发现爪哇鼷鹿威氏亚种(Tragulus javanicus williamsoni),证明了小鼷鹿在中国的分布,这也是国内惟一的分布区。直到20世纪90年代末,国内才有对该物种生境选择的初步研究。在不同的生境类型中设置长250 m、宽4 m,面积为0.1 hm2的调查样方63个,结果表明,影响爪哇鼷鹿威氏亚种的生境因子主要是下木郁闭度、地表覆盖度、坡度和植被类型。凭借这4个生境因子的组合,将生境划分为最佳生境地、良好生境地、一般生境地和劣等生境地(Luo et al, 1999)。
Meijaard & Groves (2004)对东南亚地区的鼷鹿分类作了修订,把Tragulus javanicus williamsoni从爪哇小鼷鹿(Tragulus javanicus)中分离出来作为独立种,即:威氏小鼷鹿(Tragulus williamsoni)。这一物种仅分布于泰国北部、老挝北部和云南南部。鉴于此,可以说Luo et al (1999) 的工作是目前对威氏小鼷鹿(Tragulus williamsoni)的惟一专项生态学研究,但该项研究仅是比较宏观的生境选择研究,并未回答威氏小鼷鹿对微生境因子的选择性。2005—2007年西双版纳国家级自然保护区与西南林学院(西南林业大学的前身) 合作,系统调查了勐腊地区威氏小鼷鹿的资源现状。在此基础上,本项工作深化研究内容,希望了解威氏小鼷鹿对微生境因子的选择性和探究它在云南南部的分布现状和成因,探讨威氏小鼷鹿分布格局和栖息环境之间的关系,为保护区划定威氏小鼷鹿保护的关键区域和制定有效保护措施提供基础资料。
1 方法
1.1 研究地概况
西双版纳保护区的尚勇片区位于勐腊县南部(21°13′30″~21°24′48″N, 101°22′48″101°37′30″E),总面积31 184 hm2。保护区南部边界恰是中国与老挝接壤的国境线,其长约15 km。该片区以中山、低中山为主,为切割较深的中山峡谷型地貌区;地势以南部及西南部较高,东部与北部较低,海拔610~1 691 m。低海拔地区植被类型为热带雨林和季雨林,高海拔地区以南亚热带常绿阔叶林为主(Guo et al, 1987)。根据2005年12月—2006年3月在尚勇片区的走访结果,于2006年8月—2007年12月进行威氏小鼷鹿种群数量调查(结果另文报道)。与此同时,2007年6—12月在威氏小鼷鹿种群数量较多,植被完整的勐满(21°21′8.1″~21°16′48.7″N,101°22′51.2″~101°26′19.1″E)和龙门(21°18′17.1″~21°13′34.1″N,101°26′45.1″~101°32′34.6″E)开展分布格局和生境选择利用的研究。
1.2 样线设置及足迹链坐标收集
根据走访调查得知的威氏小鼷鹿的分布信息,结合文献资料报道的生态习性,威氏小鼷鹿主要栖于热带、亚热带海拔1 000 m 以下的森林、灌丛,经常活动和隐蔽于近水潮湿地带的灌草丛或树脚(Yang & Zhang, 1987)。参考1∶50 000地形图的河流分布和走向,从谷底至山顶将山坡划分为上坡位(海拔大于950 m)、中坡位(海拔800~950 m)和下坡位(海拔小于800 m)等3部分。在坡位的每一部分各设置2~3条样线,以500 m为间隔,设置30 m ×30 m样方一个。样线行走时间为9:00—17:00,在样方中仔细寻找威氏小鼷鹿活动痕迹或足迹链。利用GPS(美洲豹GARMIN 72)实时测定每个足迹链的地理坐标。如果个别样点由于地形、植被、天气等因素导致无法接收卫星信号,则根据相邻样方在1∶50 000图上的坐标位置,再根据地形或河谷走向推算坐标数据。
1.3 植被样方设置与数据收集
在上述样方中发现威氏小鼷鹿足迹链时,即为其利用样方(used sample)。以样方中心为参照点,通过随机数字表(table of random numbers)确定利用样方和可利用样方(available sample)中心的方位角和距离,设置相同面积大小的可利用样方(Fowler et al, 1998)。在大样方四角及中央设置2 m×2 m小样方各5个,以大样方作地形、乔木、灌木的测度,以小样方作草本和落叶层的测度。可利用样方测量的因子和方法同利用样方。测量和记录的生态因子如下:(1)地形特征:海拔、坡位和距水源距离;(2)植被特征:植被类型、乔木层盖度、优势乔木、灌木层盖度、优势灌木、草本层盖度、优势草本和落叶层盖度。其中优势乔木、优势灌木和优势草本指样方中盖度和密度最大的乔木、灌木和草本种类。在11个生态因子中,坡位、优势乔木、优势灌木、优势草本和植被类型等5个因子为定性因子(nominal factor),其余6个为定量因子(quantitative factor)。
鉴于工作量的原因,未一一设置对应的可利用样方,而是采用间隔2个利用样方(有足迹链的样方)设置1个可利用样方,如果可利用样方的中心点恰好落在河中或其他不便调查区时则放弃。
1.4 数据处理
1.4.1 分布格局图制作
利用Erdas 8.4软件将勐满片与龙门片地形图(1∶50 000)数字化;将GPS点与数字化地图导入GIS软件(Arc View GIS 9.2);将GPS点与数字化地形图相结合,通过配准和叠加,在地形图上生成各足迹点,即成威氏小鼷鹿的分布格局图。
1.4.2 生态因子分析
1.4.2.1定性因子分析 坡位、优势乔木、优势灌木、优势草本、植被类型等5个定性因子采用Vanderploeg & Scavia选择系数Wi和选择指数Ei作为威氏小鼷鹿对生境喜好程度的指标。
①资源选择率ωi是动物对于资源i的选择率

式中:Oi为资源i被利用的样方数占利用样方百分比,πi为具有资源i特征的样方占样方总数百分比。

式中:Ei为资源选择指数,Wi为选择系数,n为资源数。Ei值介于−1与+1之间,若Ei= 0,表示动物对资源i的选择是随机的,用“0”表示;若Ei<0表示动物回避资源i,用“-”表示;若Ei>0表示动物偏好资源i,用“+”表示(Vanderploeg & Scavia, 1979)。
1.4.2.2定量因子分析 首先将百分数型因子(乔木层、灌木层、草本层和落叶层盖度)作反正弦函数转换为角度型因子,以提高数据的正态性。接着采用Kolmogorov-SmirnovZ检验分析利用样方和可利用样方中的6个定量因子的正态性,当数据符合正态分布时,采用独立样本的t-检验;当因子不符合正态分布,采用非参数Mann-WhitneyU-检验。选取差异显著的因子进入下一步分析。采用Spearman Correlation Coefficient判断因子之间的相关性。当两因子之间的相关系数ρ的绝对值大于或等于0.60时,则取这些比较重要的因子进入逐步判别分析(stepwise discriminant analysis),以确定影响栖息地选择的关键因子(Lahaye & Gutierrez, 1999)。使用逐步判别分析时,所有选项均为系统默认值。数据处理在SPSS 11.0 for Windows上进行(Yu & He,2003)。
2 结 果
2.1 足迹链与分布
在勐满片共收集到有效足迹链37条,威氏小鼷鹿种群呈沿河流分布的特点,种群密度在三岔河地区,即南哈河与阿山河交汇处达到最高。此处也是河流分布较为密集的地区(图1A)。
在龙门片共收集到有效足迹链73条,威氏小鼷鹿种群密度在大榕树地区最为集中,该地区属于大包包河流域。在大、小南满河地区的分布亦呈现出明显的沿河流分布的特点(图1B)。
2.2 生境选择利用
在勐满片和龙门片分别设了样方800和1 047个,仅分别收集到有效足迹链37和73条,将有足迹链的样方均作为利用样方,采集生态因子数据。按对照样方设置原则,另设可利用样方30个。
2.2.1 定性因子比较
分析结果表明,威氏小鼷鹿偏好季雨林和常绿阔叶林的中下坡位活动。对优势乔木千果榄仁(Terminalia myriocarpa)、土沉香(Aquilaria sinensis)和粗叶榕(Ficus hirta),优势灌木木奶果(Baccaurea samiflora)、斑鸠菊(Vernonia esculenta)、云南地桃花(Urena lobata)和大叶蒲葵(Livistona chinensis),优势草本长柄山姜(Alpinia kwangsiensis)和柊叶(Phrynium capitatum)有选择偏好(表1)。
2.2.2 定量因子比较
对6个定量因子的正态性检验结果表明,它们均不符合正态分布,故采用非参数Mann-WhitneyU-检验。利用样方与可利用样方相比较,仅海拔和草本盖度2个因子差异不显著,其余4个因子差异均显著,其中乔木盖度、落叶层盖度和距水源距离均差异极显著(表2)。
2.2.3 影响栖息地选择因子分析
判别分析结果显示,乔木、灌木和落叶层盖度及距水源距离是区分利用样方和可利用样方的关键因子,两类样方的区分正确率达95.7% (表3)。
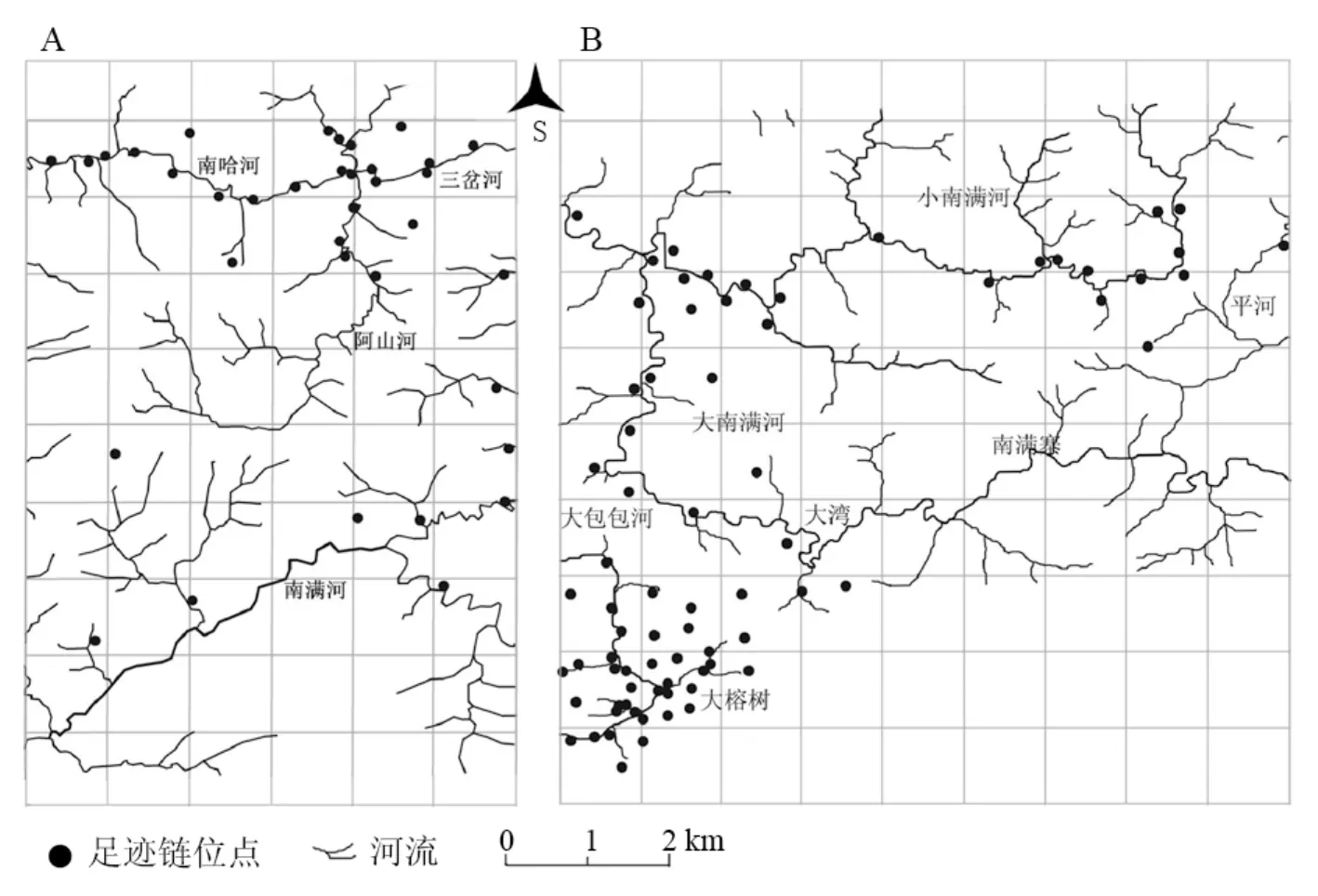
图 1 勐腊地区威氏小鼷鹿分布图(A:勐满片;B:龙门片)Fig. 1 Distributions of Tragulus javanicus in Mengla area (A:Mengman region; B:Longmen region)

表 1 威氏小鼷鹿对栖息地定性因子的选择Tab. 1 Selection of nominal factors in habitats by Tragulus williamsoni

I:资源(Resource);Oi :资源i中被利用的样方数(Quantity of used sites include resource i);πi :具有资源i特征的样方总数(Quantity of total sites include resource i);ωi :资源选择率(Selective frequency of resource);Wi :资源选择系数(Selective coefficient of resource);Ei: 资源选择指数(Selective exponent of resource)。DSTT:优势乔木(Dominant species of tall tree);DSS:优势灌木(Dominant species of shrub);DH:优势草本(Dominant herb);POS:坡位(Position on slope);VT:植被类型(Vegetation type)。 +:偏爱选择(observed usage is significantly higher than expected); 0:随机选择(observed usage in proportion to its availability);-:避免选择(observed usage is significantly lower than expected)

表 2 威氏小鼷鹿利用与可利用样方间定量因子比较Tab. 2 Comparisons of quantitative factors between used and available samples of Tragulus williamsoni

表 3 威氏小鼷鹿利用与可利用样方间因子的逐步判别分析结果Tab. 3 Results of stepwise discriminant analysis of factors between used and available samples of Tragulus williamsoni
3 讨 论
3.1 地理空间与分布格局
威氏小鼷鹿对低海拔的河谷浅滩地带有明显的选择偏好。勐满和龙门的威氏小鼷鹿均沿海拔690~936 m河谷地带的中下坡位分布。Luo et al (1999) 的工作也表明,海拔710~1 000 m的足迹较多。本项工作结果表明,威氏小鼷鹿主要分布在勐满的三岔河地区(南哈河与阿山河交汇处)和龙门的大榕树地区(大包包河流域)。据野外调查,这两个地区河面均较宽,水流较慢,河流两岸坡度平缓,植被茂密。这一切环境条件有利于胆小、活动能力较弱的威氏小鼷鹿遭遇天敌时迅速逃避或就近隐藏。这也是利用样方中距水源距离小于可利用样方,两者差异极显著的重要原因(表2)。
3.2 植被环境与分布格局
威氏小鼷鹿对植被垂直空间结构的选择与安全性的需求密切相关。检验结果显示,威氏小鼷鹿对较高的乔木和灌木盖度有选择偏好(表2)。利用样方中,较密的乔木树冠层能在乔木层顶端提供较高密度的枝叶,保证了较高的郁闭度,对于威氏小鼷鹿来说是一种很好的隐蔽条件。在利用样方和可利用样方之间,草本盖度差异不显著(表2),说明研究区域草本植被的密度和盖度变化不大;相比之下,灌木盖度差异显著(表2),从而看出威氏小鼷鹿主要以寻找有更茂密的灌木区域来满足反捕食的生境需求。对该地区威氏小鼷鹿生境选择的初步研究结果亦反映,植被类型和下木郁闭度是其首选生态因子(Luo et al, 1999)。高密度的灌木丛可提供良好的隐匿条件,高大型乔木能提供减少被天敌发现的机率。这样的植被环境对于威氏小鼷鹿来说,就形成了双重的隐蔽条件,大大提高了安全性。逐步判别分析结果亦显示:乔木盖度和灌木盖度是影响威氏小鼷鹿生境利用的主要因子,它们分别排在第一位和第二位(表3)。调查区域内生长着千果榄仁、土沉香和粗叶榕等大型乔木,林下均有较密的灌木分布,植被属季节性雨林和常绿阔叶林类型。在威氏小鼷鹿集中分布的三岔河与大榕树地区,人烟稀少,较小的人为干扰保证了该地区植被的完整,为威氏小鼷鹿提供了较大面积的适宜生境。此外,在海拔较低的河谷地带,由于距水源较近,乔木层及林下灌层生长状况较高海拔地区更为茂密,亦是威氏小鼷鹿在勐腊地区形成沿海拔较低的河岸集中分布格局的重要原因。国外的研究结果亦表明:鼷鹿科动物喜爱在近水源、有密集灌丛的区域活动(Barrette, 1987; Nowak, 1991)。
食源植物沿河谷的两岸分布是影响威氏小鼷鹿分布格局的重要原因。经走访调查,威氏小鼷鹿主要食物包括榕树科植物、木奶果和余干子的果实,也取食柊叶、姜科等草本植物的嫩叶。资源选择指数(Ei)的结果显示:威氏小鼷鹿对以上食源植物均具选择偏好(表1)。在威氏小鼷鹿集中分布的溪沟河流区域,植被形成了一条天然的林窗带或林缘带。在此区域,粗叶榕、木奶果、长柄山姜和柊叶等植物密度较高,食源植物沿河谷的两岸集中分布,促使了威氏小鼷鹿沿河流的两岸分布格局的形成。对婆罗洲Kabili-Sepilok保护区的爪哇小鼷鹿生境研究结果亦发现,林窗或林缘区域的植物多样性高,可为小鼷鹿提供数量更多的果实和嫩叶,是小鼷鹿取食的重要场所(Whitmore, 1998; Matsubayashi et al, 2003)。
致谢:西双版纳国家级自然保护区尚勇管理所王力繁所长和勐腊管理所许海龙所长及两个所的管理人员为本项研究提供了热情周到的后勤服务和人力资源支持;西南林学院野生动植物保护与利用专业硕士研究生刘钊(2006级)和刘恺(2007级)参加部分野外调察,在此一并表示谢意!
Barrette C. 1987. The Comparative Behavior and Ecology of Chevrotains, Musk Deer, and Morphologically Conservative Deer [C]// Wemmer CM. Biology and Management of the Cervidae.Washington:Smithsonian Institution Press, 200-213.
Dubost G. 1975. Le comportement du chevrotain african, Hyemoschus aquaticus Ogiby (Artiodactyla, Ruminantia) [J]. Z Tierpsychol,37: 449-501.
Dubost G. 1978. Un apercu sur l'ecologie du chevrotain african Hyemoschus aquaticus Ogiby, Artiodactyle Tragulide [J]. Mammalia,42: 1-62.
Fowler J, Cohen L, Jarvis P. 1998. Practical Statistics for Field Biology. [M]. 2nd ed. West Sussex: Open University Press.
Lahaye WS, Gutierrez RJ. 1999. Nest sites and nesting habitat of the northern spotted owl in north western California. [J]. Condor,101: 324-330.
Luo ID, Huang JG, Dong YH, Song JP, Tang AJ, Feng GQ. 1999. Preliminary study on habitat selection of Tragulus javanicus williamsoni Kloss in Xishuangbanna Nature Reserve [J]. Yunnan forest Sci Technol,87(2): 66-70. [罗爱东,黄建国,董永华,宋军平,唐爱军,冯国庆. 1999.西双版纳自然保护区鼷鹿生境选择的初步研究.云南林业科技,87(2): 66-70.]
Matsubayashi H, Bosi E, Kohshima S. 2003. Activity and habitat use of lesser mouse-deer (Tragulus javanicus) [J]. J Mammal,84(1): 234-242.
Meijaard E, Groves CP. 2004. A taxonomic revision of the Tragulus mouse-deer (Artiodactyla) [J]. Zool J Linnean Soc,140: 63-102.
Miura S, Idris AH. 1999. Present status and group size of the mouse-deer on Pulau Tioman, Malaysia [J]. Malayan Nat J,53: 335-339.
Miura S, Yasuda M, Ratnam LC. 1997. Who steals the fruits? Monitoring frugivory of mammals in tropical rain forest [J]. Malayan Nat J,50: 183-193.
Nowak RM. 1991. Walker's Mammals of the World [M]. 5thed. New York: The Johns Hopkins University Press.
Peng HS, Yang L, Yang YG. 1962. New Mammalian Record of Family, Genera and Species from South Yunnan [C]// The Abstract Collection about Animal Ecology and Fauna in the Professional Workshop. 206. [彭鸿绶,杨岚,杨余光. 1962.云南南部兽类科属种的新记录.动物生态及分类区系专业学术讨论会论文摘要汇编. 206.]
Vanderploeg HA, Scavia D. 1979. Calculation and use of selective feeding Coefficients: Zooplankton Grazing [J]. Ecol Model,7: 135-150.
Whitmore TC. 1998. An Introduction to Tropical Rain Forests [M]. NewYork: Oxford University Press.
Guo YQ, Yang YM, Tang JS, Chen SW, Lei FG, Wang JH, Yang ZH, Yang GZ. 1987. The Vegetation Investigation of the Conservation Area in Xishuangbanna [C]// Xu YC, Jiang HQ, Quan F. Comprehensive Investigation Reports of Xishuangbanna Nature Reserve. Kunming: Yunnan Science and Technology Press, 88-169. [郭荫卿,杨宇明,汤家生,陈述旺,雷福光,王建皓,杨中汉,杨冠中. 1987.西双版纳自然保护区植被考察报告.徐永椿,姜汉侨,全复.西双版纳自然保护区综合考察报告集.昆明:云南科技出版社, 88-169.]
Yang DH, Zhang CJ. 1987. Quantitative Distribution and Protection of Rare Animals in Xishuangbanna [C]// Xu YC, Jiang HQ , Quang F (eds) Comprehensive Investigation Reports of Xishuangbanna Nature Reserve. Kunming: Yunnan Science and Technology Press, 311-325. [杨德华,张存杰. 1987.西双版纳珍稀兽类数量分布及其保护.徐永椿,姜汉侨,全复.西双版纳自然保护区综合考察报告集.昆明:云南科技出版社, 311-325.]
Yu J Y, He XH. 2003. Statistics and Analysis for Data the Application of SPSS [M]. Beijing: Post & Telecommunications Press, 292-310. [余建英,何旭宏. 2003.数据统计与SPSS应用.北京:人民邮电出版社, 292-310.]
Habitat Use of Williamson’s Mouse-deer (Traguluswilliamsoni) in Mengla Area, Southern Yunnan
CAO Ming1, ZHOU Wei1,*, BAI Bing1, ZHANG Qin1, WANG Bin2, CHEN Min-Yong2
(1. Faculty of Conservation Biology, Southwest Forestry University, Key Laboratory of Forest Disaster Warning and Control in Yunnan Province, Kunming 650224, China; 2. Administration Bureau of Xishuangbanna National Nature Reserve, Jinghong 666100, China)
Combining line-transect method and sampling method, the geographic coordinates of footprint links for Williamson’s Mouse-deer (Traguluswilliamsoni) were collected in Mengman and Longmen regions of Mengla area, southern Yunnan, from June to December 2007. A distribution pattern map of the mouse-deer was produced by GIS software. Eleven ecological factors were measured in collected and available samples. The results indicated that the mouse-deer preferred valleys along banks of rivers or streams in low altitude, and were mainly distributed in Sanchahe of Mengman region and Darongshu of Longmen region. Comparing nominal factors between collected and available samples, the results showed that the mouse-deer tended to inhabit middle and lower slopes and favored the plant species ofFicus hirtal,Baccaurea samiflora,Alpinia kwangsiensisandPhrynium capitatum. These plants occur along banks of rivers or streams, one of the important reasons affecting distribution of the mouse-deer populations. Comparing quantitative factors between collected and available samples, the results showed that four factors including tall tree coverage, shrub coverage and herb coverage, and distance to water, were significantly different. These four factors were key in discriminating collected and available samples and main factors affecting the habitat use of the mouse-deer. The dense shrub with taller trees could form good shady condition for the mouse-deer, and greatly strengthen their security. Due to their preference for areas near water, there were more flourishing tall trees and dense shrubs at valleys in low altitude, which was one of important reasons affecting the distribution of the mouse-deer populations massing in valleysin low altitude.
Williamson’s Mouse-deer (Traguluswilliamsoni); Habitat use; Distribution pattern; Southern Yunnan
Q959.849;Q958.11
A
0254-5853-(2010)03-0303-07
10.3724/SP.J.1141.2010.03303
2009-08-05;接受日期:2009-11-11
云南省林业厅物种保护项目(200607)资助;云南省重点学科野生动植物保护与利用项目(XKZ200904)支持
曹明(1983-),男,云南西双版纳人,硕士研究生,主要从事野生动物与自然保护区管理研究
*通讯作者(Corresponding author),周伟(1957-),男,云南保山人,教授,博士生导师,主要从事野生动物与自然保护区管理研究, Email: weizhouyn@163.com
