太行山猕猴掌(跖)骨重量与颅长的异速生长分析
2010-11-26党晓云梁芝栋赵晓进段文存丁金芝
党晓云,梁芝栋,赵晓进,段文存,丁金芝
(1.中国地质大学 地球科学与资源学院,北京 100083;2.河南师范大学 体育学院,河南 新乡 453007;3.河南师范大学 生命科学学院,河南 新乡 453007)
有关灵长类的掌骨和跖骨的形态学、性差及种间相似性长期以来成为人类学家和动物学家感兴趣的话题.McFadden等[1-2]对狒狒的掌骨和跖骨的长度和重量进行了相关研究;之后又对大猩猩和黑猩猩的掌(跖)骨相对长度的性差进行研究;赵晓进等[3-5]对太行山猕猴掌骨和跖骨的形态学、掌骨和跖骨长度排序、性差等进行了研究,尤其对太行山猕猴的掌(跖)骨长度和重量的性差等方面进行了比较翔实和深入的研究.对探讨灵长类的食性、适应、性别鉴定、种间比较和系统演化等方面都具有重要意义.对于猕猴掌骨和跖骨有关变量与颅长的异速生长分析的相关文献报道较少.太行山猕猴目前是我国黄河以北分布最集中、数量最大的猕猴自然分布种群[4].在形态、生理、生态、行为、遗传和食性等方面均有其特殊性,所以具有极高的理论研究意义和应用价值.本文中通过对太行山猕猴的掌(跖)骨重量和颅长进行测量和异速生长分析,为灵长类生物多样性研究提供参考.
1 材料和方法
成年的太行山猕猴,颅骨标本28例,掌(跖)骨标本28例,其中雄性12例,雌性16例.标本来自太行山区河南省济源市和卫辉境市辖区,现存于河南师范大学生命科学学院标本馆.
颅长的定点测量采用吴汝康等的方法,测量仪器采用精密度为0.02 mm的游标卡尺.掌(跖)骨重量(以质量计)的测定采用BS110S型号的电子天平测量,精确到0.001 g.
为了便于描述,掌骨和跖骨分类标记,第1掌骨标记为1Mc,第2掌骨标记为2Mc,其它类推;第1跖骨标记为1Mt,第2跖骨标记为2Mt,其它类推.
指数方程通常被用来研究机体不同部位有关变量间的关系.异速生长分析的指数方程模型:Y=aXb; 在这里X为自变量(颅长),Y为因变量(掌跖骨质量),a为异速生长常数,b为异速生长指数.如果b=1.0,则两变量之间具有几何学上的呈等速生长;当b>1.0时,X与Y之间为正的异速生长;当b<1.0时则为负的异速生长.对上述等式进行对数转换得到相应的线性回归方程:lnY=lna+blnX,这其中,lna是截距,b是斜率.
采用SPSS 13.0分析软件,用线性模型和幂函数模型进行拟合.
2 结果
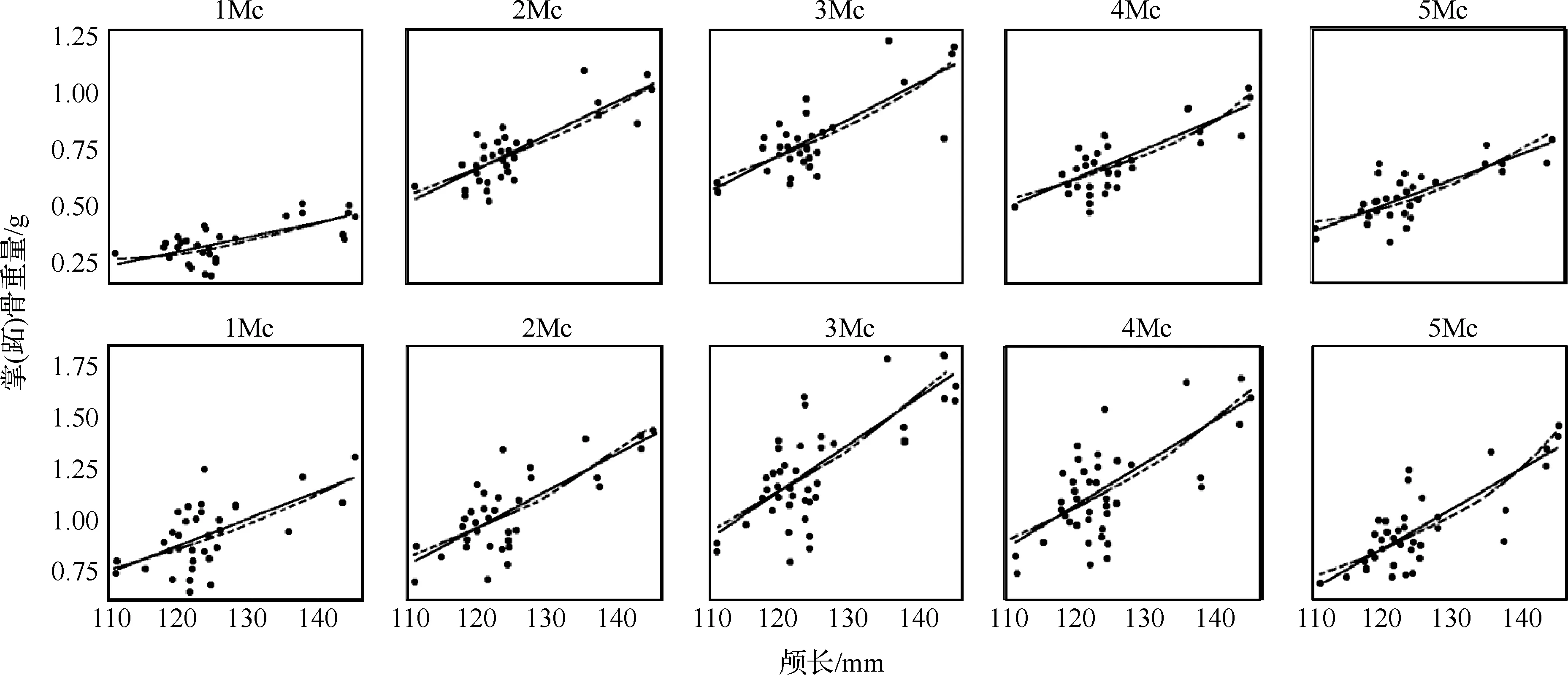
掌(跖)骨重量的异速生长系数及决定系数见表1.掌(跖)骨的重量与颅长大多数呈正异速生长(b>1.0),拟合程度较好.总体来看第1根掌(跖)骨重量的b值最小,分别为1.989和1.634.第5根掌(跖)骨重量的b值最大,分别为2.357和2.367,其它3根掌(跖)骨的b值比较接近.掌骨与跖骨之间的差异不是很大.雌雄之间的差异主要表现在第1根掌骨之间,雄性个体第1根掌骨呈正异速生长(b>1.0),雌性个体第1根掌骨呈负异速生长(b<1.0).所测数据进行对数转换后,其线性图见图1.实线为雄性,虚线为雌性,可以看到:1Mc、5Mc、3Mt、4Mt的雌雄个体之间有明显的差异.从拟合程度上来看,雄性的较好,决定系数值较大.而雌性的较差,决定系数值较小.
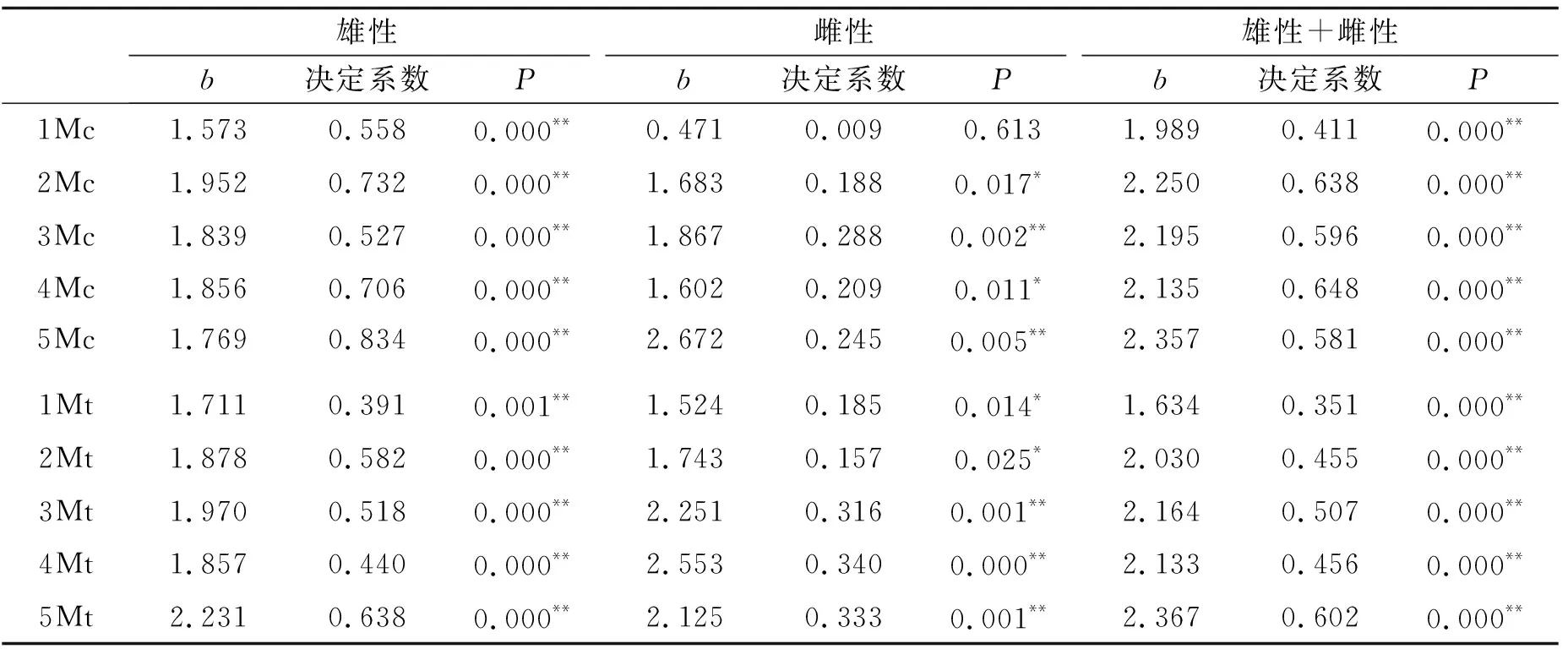
表1 太行山猕猴掌(跖)骨重量异速生长分析结果
*P<0.05,**P<0.01

线性模型和幂函数模型拟合结果进行比较见表2,可以看出各掌(跖)骨中采用线性模型的拟合程度均高于幂函数模型.
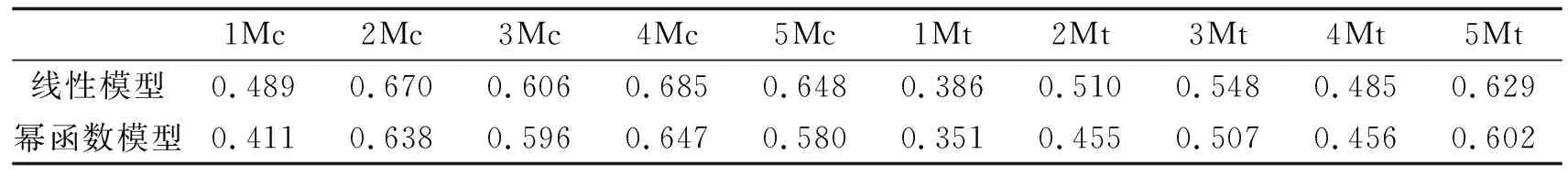
表2 线性和幂函数模型拟合程度的比较决定系数
3 讨论
在动物形态学研究中,多采用异速生长分析的方法来研究局部与整体变量在生长发育过程中的形态变化以及功能与环境的适应性[6-9].表1的分析结果可以看出雄性的决定系数值较大(P<0.001),说明曲线拟合程度较好.由其掌(跖)骨的重量来推测颅长(或身体大小)的应用价值比较高;而雌性的决定系数值比较小,拟合程度较差,应用价值比较小.雄性第5根掌骨和第5根跖骨的决定系数值最大,分别为0.864和0.638,由此推出它们的颅长的相关性较高.
选择异速生长拟合模型时发现,使用幂函数模型的效果并不理想,各掌(跖)骨的拟合程度还没有线性模型拟合程度高.在以前异速生长研究中很少出现这种情况.这说明,掌(跖)骨重量与颅长的相关性很大,接近线性关系.有资料报道指数方程比简单的线性模型更适于描述两个变量之间的关系,更符合实际的生长发育规律[6-7].可见这种情况并不一定适用于每个变量,有些变量之间的线性模型优于其他的拟合模型.所以在定量研究动物的形状和大小时,需认真考虑要解决的实际问题,注意选择构成这些变量间相互关系的合适模型及曲线拟合技术.这里为什么要采用幂函数分析?主要原因有两个:(1)在描述两个变量特别是部分变量与整体变量进行异速生长或相关分析时,以前的相关资料都是采用异速生长曲线拟合[6-7].实际上大部分情况下,变量之间的关系都是非线性的.为了便于比较这里采用了线性和非线性两种模型.(2)猕猴掌跖骨重量的发育包括与颅长的相互关系是比较特殊的,有很多资料表明在很大程度上它是受出生前遗传和激素的调节,并不像其他长骨那样主要受出生后各种环境因子的影响;为了表明这种特殊性,本文中采取两种模型进行比较.
太行山猕猴的掌(跖)骨重量均呈正异速生长.即在太行山猕猴的生长发育过程中,随着颅长(体重)的变化,掌(跖)骨的重量以更大的比例增长,生长趋势与其功能相适应;各掌(跖)骨间的增长速度是有差异的,第1根掌(跖)骨增长的速度最慢,第5根掌(跖)骨增长的速度最快,其它3根接近同步.雌雄个体之间也存在着显著的差异,雄性的拟合程度要优于雌性的,表明雄性掌(跖)骨的重量与颅长有更好的相关性,雌雄之间存在着不同的生长模式,这可能与出生后的生活环境、营养、体质、健康状况、激素、以及个体在群体中的地位等有关.研究结果表明太行山猕猴的掌(跖)骨的长度变量和重量变量与颅长的相关性是不同的.掌跖骨的重量变化可能与动物身体大小(颅长)的关系更为密切[8].
对于指数模型Y=aXb来说,自变量X的选择并不相同.虽然体重因为涉及到身体构筑、运动、生态、种群结构、生活史、社会结构、性差及其他参数而通常被作为灵长类研究的基础[5].但是有时研究异速生长时(尤其是研究化石材料时),最好的方法是用与身体构筑其他有关的变量.如有的学者用肱骨最大长、颅骨最大长和下颌长等作自变量代表体重或个体大小[8-10].通过其他变量(如掌跖骨重量)与这些变量(如颅长)之间的异速生长分析,进而探讨它们与体重之间的相互关系.一般不直接用体重作自变量的主要原因是直接引用灵长类野生体重比较困难或体重在化石中不能直接测量,有时是因为体重变量受较多因素影响(例如,季节、营养、疾病等).然而在现存的动物种类中,使用体重来进行比较研究还是被认为是很有效的方法.
参考文献:
[1] McFadden D,Bracht M S.The relative lengths and weights of metacarpals and metatarsals in baboons(Papiohamadryas) [J].Horm Behav,2003,43(2):347-355.
[2] Mcfadden D,Bracht M S.Sex difference in the relative lengths of metacarpals and metatarsals in gorillas and chimpanzees[J].Horm Behav,2005,47(1):99-111.
[3] 赵晓进,赵俊杰,王耿,等.太行山猕猴掌骨和蹠骨长度的性别判别[J].人类学学报,2008,28(1):88-94.
[4] 赵晓进,王训练,党晓云,等.太行山猕猴掌骨和蹠骨长度比率的性别差别[J].解剖学报,2009,40(6):993-996.
[5] 党晓云,赵晓进,梁芝栋,等.太行山猕猴掌骨和趾骨体质量的性差与排序[J].解剖学杂志,2009,32(4):533-535.
[6] 潘汝亮,王红.金丝猴长骨的异速生长研究[J].动物学研究,1989,10(1):23-30.
[7] 薛德明.太行山猕猴肩胛骨变量间的相关性和异速生长分析[J].兽类学报,2000,20(1):76-79.
[8] 路纪琪,薛德明,吕九全,等.太行山猕猴颅骨变量的异速生长研究[J].动物学报,2001,47:172-177.
[9] 翟鹏飞,赵晓进,雷梦云,等.太行山猕猴下颌骨的异速生长分析[J].解剖学杂志,2005,28(2):219-221.
[10] Barrie P.The allometry of primate skeletal weight [J].International Journal of Primatology,1986,7(5):457-466.
