CUL5基因沉默对连续照射期间CNE2细胞增殖状况影响的研究
2010-09-20朱小东李烨曲颂李龄黎丹戎张玮
朱小东 李烨 曲颂 李龄 黎丹戎 张玮
广西医科大学附属肿瘤医院放疗科,*第六实验室,广西 南宁 530021
鼻咽癌常规分割放射治疗期间存在的肿瘤细胞加速再增殖现象是导致鼻咽癌治疗失败的重要原因之一,但其机制未明。有研究证实,细胞受照射后其增殖活性的改变与细胞周期基因有关[1]。本研究已在前期应用基因芯片技术、实时荧光定量PCR筛选出与照射后鼻咽低分化鳞癌细胞(CNE2)增殖状态相关的差异基因,其中CUL5为细胞增殖活性增高时的上调基因之一[2]。本研究通过离体细胞实验的方法,采用RNA干扰技术探索CUL5表达情况对鼻咽癌放射治疗中细胞增殖活性的影响,初步探索鼻咽癌放射治疗中细胞加速增殖的机制。
1 材料和方法
1.1 材料 pGPU6/GFP/Neo质粒(上海吉玛公司);鼻咽低分化鳞癌细胞株CNE2及感受态DH5α均由本实验室保存;限制性内切酶(BamHⅠ,PstⅠ,BbsⅠ)及T4 DNA连接酶(TakaRa公司);lambda/Eco130I marker(MBI公司);质粒小量抽提试剂盒(Omega Bio-tek公司);胶回收试剂盒(上海捷瑞生物工程有限公司);Trizol(Invitrogen公司)、LipofectamineTM2000(Invitrogen公司)、逆转录试剂盒(MBI公司);G418、四甲基偶氮唑蓝(MTT)(Sigma公司);Cycle TESTTMPLUS DNA Reagent Kit(BD公司)。
1.2 方法
1.2.1 细胞培养 CNE2细胞加入含10%小牛血清的RPMI1640培养基,于37 ℃、CO2体积分数为5%的饱和湿度条件下培养,常规传代,培养至对数生长期。
1.2.2 寡核苷酸的设计合成 根据 Genebank中CUL5的序列,按照Tuschl 设计原则[3]由吉玛公司设计合成3对siRNA序列(表1),赠送一对通用阴性对照(negative control,NC)序列。用LipofectamineTM2000将以上序列瞬时转染CNE2细胞,48 h后提取细胞总RNA,逆转录合成cDNA,行CUL5基因的PCR扩增(上游引物序列:5’-AATGGCATAGGCACCTTGTC-3’;下游引物序列:5’-GGGAATGAACAGCAGCAAAT-3’,产物长度130 bp),以β-actin为内参(上游引物序列:5’-GATGACCCAGATCATGTTTG-3’;下游引物序列:5’-TGGAGTTGAAGGTAGTTTCG-3’,产物长度491 bp),根据凝胶成像系统分析所得CUL5灰度比值筛选出沉默效果最好的siRNA序列,此序列按照表达质粒pGPU6/GFP/Neo的要求,设计寡核苷酸序列。CUL5正义链:5’-CACCGACACGACGTCTTATATTATTCAAGAGATAATATAAGACGTCGTGTCTTTTTTG-3’;反义链:5’-GATCCAAAAAAGACACGACGTCTTATATTATCTCTTGAATAATATAAGACGTCGTGTC-3’。由上海生物工程技术服务有限公司合成。

表1 siRNA靶位及序列Tab.1 Targets and sequences of siRNA
1.2.3 重组质粒pGPU6/GFP/Neo-CUL5的构建及鉴定 将合成的DNA oligo退火处理所得shRNA模板通过T4 DNA连接酶与pGPU6/GFP/Neo载体双酶切(BamHⅠ+BbsⅠ)后胶回收所得线性化空载体进行连接。连接产物常规转化、筛选及限制性内切酶BamHⅠ、PstⅠ分别酶切鉴定,对可能重组成功的质粒,送上海英骏生物技术有限公司进行DNA测序鉴定以进一步证实。
1.2.4 重组质粒的转染及细胞克隆筛选 用LipofectamineTM2000将重组干扰质粒pGPU6/GFP/Neo-CUL5转入CNE2细胞中,设为实验组(称为pGPU6/GFP/Neo-CUL5组)。同时设通用阴性对照重组质粒pGPU6/GFP/Neo-NC转染CNE2细胞的阴性对照组(pGPU6/GFP/Neo-NC组)及未转染的空白对照组(CNE2组)。转染48 h后,用含G418(筛选浓度300 μg/mL)的培养基培养14 d至抗性克隆出现,采用有限稀释法获取G418抗性单克隆,并以G418维持浓度(150 μg/mL)扩大培养,建立稳定传代的阳性细胞。
1.2.560Co-γ射线照射 使用60钴远距离治疗机照射细胞,剂量率为108~111 cGy/min,放射源到细胞的距离为80 cm,机头和机架角度均为0°,调整细胞上方RPMI 1640 培养液(含10%小牛血清)的深度为5 mm,以此作为60Co-γ射线照射所需剂量建成区,照射野面积为30 cm×30 cm,且照射细胞摆放位置均位于照射野边缘2 cm以内的区域。2 Gy/d,每天1次,连续照射5 d。
1.2.6 RT-PCR检测60Co-γ照射前、后阳性细胞CUL5基因mRNA表达水平 取未经照射及连续分割照射第1、3、5天的细胞,培养24 h后,提取细胞总RNA,逆转录合成cDNA,行CUL5基因的PCR扩增,以β-actin为内参,凝胶成像系统分析CUL5的灰度比值,实验重复3次。
1.2.7 MTT法测连续照射期间阳性细胞相对增殖率 将单细胞悬液以每孔1×103个细胞接种于96孔板中进行细胞培养及照射。分别于照射第1~5天,每次照射后24 h,每孔加入MTT溶液(5 mg/mL)20 μL继续温育4 h,终止培养,小心吸弃孔内培养上清液,每孔加入150 μL二甲基亚砜(DMSO),振荡10 min,使结晶充分溶解;在酶联免疫检测仪上测定各孔光吸收值(A值),取相同生长天数及转染相同质粒的A值的均值作为该组细胞某天的A值,计算照射后细胞相对增殖率。
1.2.8 流式细胞术(FCM)检测连续照射期间阳性细胞的细胞增殖情况 将单细胞悬液以每孔1×105个细胞接种于6孔板中进行细胞培养及照射,分别于照射第1~5天,每次照射后24 h,胰酶消化,室温300×g离心5 min,吸去上清液,加入1 mL缓冲液重悬细胞,重复几次,调整细胞浓度到1×106个/mL,固定,上机检测。具体步骤详见Cycle TESTTMPLUS DNA Reagent Kit 说明书。
1.2.9 统计处理 采用SPSS 13.0软件分析,数据以均数±标准差表示,组间差异采用单因素方差分析,以P<0.05为差异有统计学意义。
2 结 果
2.1 siRNA靶序列选择 RT-PCR检测各种转染细胞CUL5基因的表达情况(图1),通过凝胶电泳条带分析结果(图2)显示,转染阴性对照的细胞CUL5基因表达无明显变化,转染siRNA的细胞CUL5基因表达有不同程度下调,以β-actin为内参校正,靶位699、1190、2039的灰度比分别为0.398、0.776、0.337,靶位2039的siRNA下调最明显,可作为构建CUL5干扰RNA真核细胞表达载体的靶点。

图1 CUL5基因表达的电泳结果Fig.1 Expressions of CUL5 after the transfection of siRNAs
2.2 重组质粒的鉴定
2.2.1 重组质粒的酶切鉴定 酶切结果表明,所有质粒均为阳性重组载体(图2)。
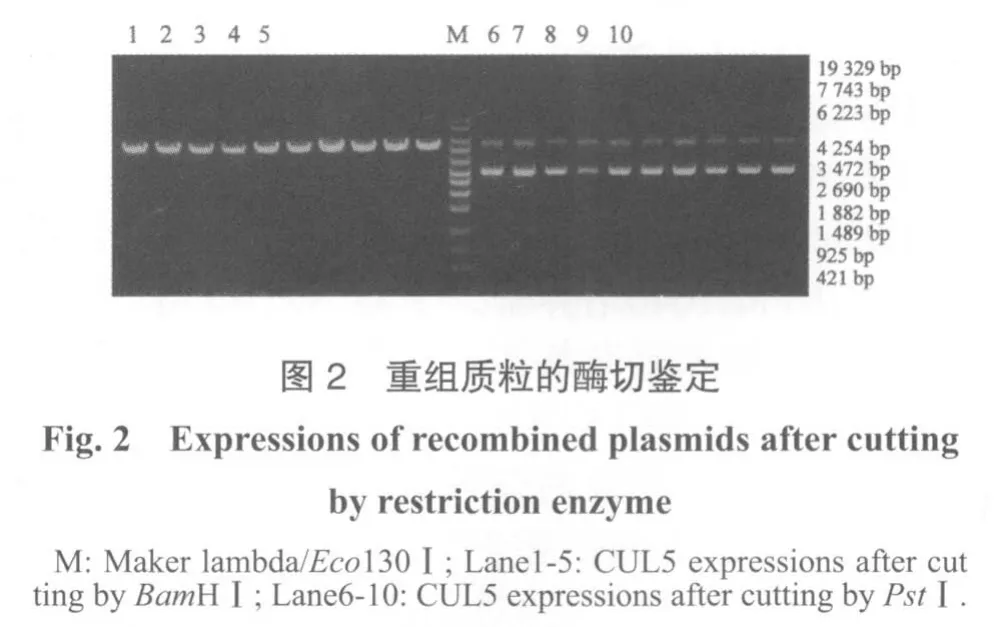
2.2.2 重组质粒测序鉴定 DNA序列测定结果表明,序列已成功插入质粒pGPU6/GFP/Neo,载体构建成功(图3)。将重组质粒命名为pGPU6/GFP/Neo-CUL5。

图3 重组质粒pGPU6/GFP/Neo-CUL5测序图(部分)Fig.3 Part of the sequence map of pGPU6/GFP/Neo-CUL5
2.3 未照射阳性细胞CUL5基因沉默效果RT-PCR检测未经照射的3组细胞CUL5基因的表达情况(图4),通过凝胶成像系统分析电泳条带,以β-actin为内参校正,测定CNE2组、pGPU6/GFP/Neo-NC组、pGPU6/GFP/Neo-CUL5组CUL5的灰度比分别为0.122、0.175和0.004,表明pGPU6/GFP/Neo-CUL5组的CUL5基因表达受到明显抑制。

图4 照射前各细胞株CUL5基因表达的电泳结果Fig.4 Expressions of CUL5 in each group before irradiation
2.4 照射后阳性细胞CUL5基因沉默效果照射第1天CNE2组、pGPU6/GFP/Neo-NC组、pGPU6/GFP/Neo-CUL5组CUL5灰度比分别为0.667、0.786和0;照射第3天分别为0.538、1.222和0;照射第5天分别为0.818、1.381和0,表明pGPU6/GFP/Neo-CUL5组在受照射后CUL5基因表达仍受到明显抑制。
2.5 MTT法测各组细胞照射后不同时间相对增殖率 结果显示连续照射5天期间,CNE2组及pGPU6/GFP/Neo-NC组细胞以第3天相对增殖率最高,pGPU6/GFP/Neo-CUL5组第1天相对增殖率最高,3组均以第5天最低(表2)。经单因素方差分析,pGPU6/GFP/Neo-CUL5组与pGPU6/GFP/Neo-NC组及有CNE2组差异均有统计学意义(P<0.001)、pGPU6/GFP/Neo-NC组与CNE2组的差异无统计学意义(P=0.741)。

图5 照射后各细胞株CUL5基因表达的电泳结果Fig.5 Expressions of CUL5 in each group after irradiation at the 1st, 3rd and 5th day
2.6 FCM检测照射后细胞增殖情况 FCM通过测定细胞内DNA含量,分析照射后pGPU6/GFP/Neo-CUL5及CNE2组细胞中处于G0、G1、S期的比例,比较其增殖能力。根据公式计算出每天的S期细胞百分率(SPF)及增殖指数(PI)值(表3、4),可知2 Gy/d连续照射期间,pGPU6/GFP/Neo-CUL5组细胞在照射第1天SPF及PI值均为最高值,说明此时细胞增殖最旺盛,而在照射第5天时,SPF及PI值均为最低值,提示此时细胞增殖活性最低,增殖速度减慢;CNE2组细胞在照射第3天SPF及PI值均为最高值,增殖活性最高,照射第5天时,SPF及PI值均为最低值,增殖速度减慢。
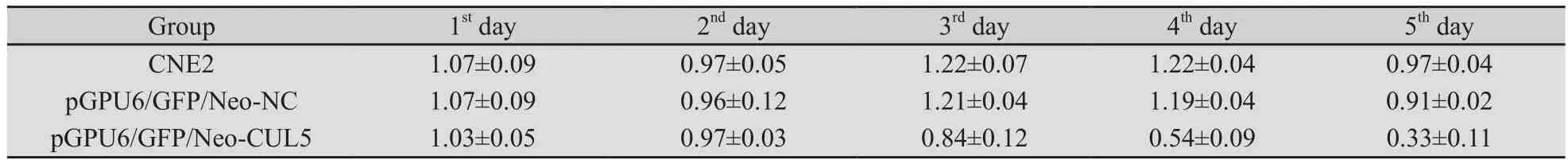
表2 各组细胞照射后不同时间相对增殖率Tab.2 Relative growth rate of each group after irradiation at different days

表3 照射后pGPU6/GFP/Neo-CUL5组细胞周期分布情况Tab.3 Distribution of cell cycle in cells transfected with pGPU6/GFP/Neo-CUL5 after irradiation(%)

表4 照射后CNE2组细胞周期分布情况Tab.4 Distribution of cell cycle in CNE2 after irradiation(%)
3 讨 论
Hermens和Barendsen[4]研究大鼠横纹肌肉瘤时发现照射1周后,肿瘤中增殖细胞比例上升,克隆细胞倍增时间由4 d缩短到1.5 d,认为肿瘤干细胞已开始加速再增殖;本课题组前期研究[5]应用不同的实验方法,已明确了人鼻咽癌细胞株CNE2在连续分割照射中确实存在中后期加速再增殖现象,并通过基因芯片筛选出CUL5为细胞增殖活性增高时的上调基因之一,其在照射增殖活性最高时相与最低时相表达差异在2倍以上。
Singhal等[6]在检测早期肺腺癌与正常肺组织间存在差异的细胞周期基因时首次发现了CUL5基因。此后,Burnatowska-Hldin等[7]和Baxter 等[8]也分别研究发现乳腺癌T47D细胞增殖速度、急性早幼粒细胞性白血病中粒细胞系的分化和CUL5的表达存在相关性。CUL5基因编码的Cul5支架蛋白,可介导蛋白酪氨酸激酶的泛素化,还与p53/TP53的降解有关[9],其具体功能目前尚在研究之中。
本研究为了明确CUL5基因表达增强是否真与临床观察到的鼻咽癌细胞的增殖活性增高有关,采用RNA干扰技术沉默CUL5基因后,给予实验组、阴性对照组、空白对照组鼻咽低分化鳞癌细胞株CNE2 2 Gy/d,连续照射5 d,并采用MTT对每天照射后的细胞进行检测,按公式计算每次照射后细胞相对增殖率,实验组分别为1.03±0.05、0.97±0.03、0.84±0.12、0.54±0.09、0.33±0.11,增殖速度呈下降趋势,照射第4、5天,细胞增殖明显减慢;阴性对照组分别为1.07±0.09、0.96±0.12、1.21±0.04、1.19±0.04、0.91±0.02,空白对照组分别为1.07±0.09、0.97±0.05、1.22±0.07、1.22±0.04、0.97±0.04,均在照射第3、4天时,细胞增殖加速,照射第5天时增殖速度减慢,这与我们[5]前期研究结果一致。实验组与阴性及空白对照组比较差异均有统计学意义(P<0.001),提示CUL5基因沉默后,照射中后期CNE2细胞加速增殖现象改变,说明CUL5基因表达增强与CNE2细胞照射中后期加速增殖有关。S期细胞百分率(SPF)指正在整个细胞群体中处于S期的细胞所占的百分比,是表示细胞增殖活性的重要指标。细胞增殖指数(PI)代表了细胞群的增殖状态,是指细胞群各期细胞分布百分比中的DNA合成期(S期)和RNA合成期(G2/M期)细胞百分比之和。连续分割照射期间,流式细胞术检测细胞周期结果显示,实验组在连续照射第1天S期细胞百分率最高(SPF=29.35%),细胞增殖指数同样最高(PI=43.32%),后SPF及PI值均逐渐下降,第5天最低(SPF=15.47%,PI=25.32%),说明细胞增殖活性逐渐降低,增殖速度逐渐减慢;对照组第3天增殖活性最强(SPF=35.34%,PI=50.69%),第5天增殖速度最慢(SPF=19.35%,PI=30.67%)。这与MTT法所得结果一致,进一步验证了其结果。
RNA干扰技术是通过将19nt-23nt的siRNA导入细胞,降解与其同源的mRNA,特异、高效地阻断目的基因表达[10]。RNA干扰技术因其特异性和高效性,近年来已成为研究基因功能的重要工具。本实验应用RNA干扰技术使CUL5基因沉默,导致其编码的Cul5蛋白表达抑制,进而影响Cul5蛋白介导的一系列蛋白质的降解,如p53蛋白和蛋白酪氨酸激酶。
野生型p53蛋白在DNA损伤后使细胞生长停滞于G1期,诱导细胞程序性死亡,抑制肿瘤细胞生长,维持遗传稳定性[11]。离子辐射、紫外辐射和DNA损伤剂等均可造成DNA损伤而导致p53蛋白活化,然后p53转录激活下游与肿瘤抑制有关的基因。石慧英等[12]研究发现高表达的P53蛋白可以使DNA损伤的CNE2细胞停滞于G1期,但仍然参与放射诱导的鼻咽癌细胞的损伤和凋亡过程。蛋白酪氨酸激酶在正常细胞的调节、通讯和发育等生物学方面起着十分重要的作用;其活性过高可导致下游信号途径激活,从而导致细胞转化、增殖、对抗细胞凋亡、促进细胞存活。本实验中CUL5基因表达抑制后CNE2细胞连续5 d分割照射后呈增殖状态,推测可能是以上两种途径共同作用的结果。照射前2天实验组细胞的增殖活性略高于对照组,可能是CUL5基因沉默后,Cul5蛋白表达减少,蛋白酪氨酸激酶因不能正常泛素化、降解,活性增高,激活其下游信号途径,导致细胞增殖能力加强;中后期加速增殖现象改变,增殖速度减慢,则可能是由于照射剂量累加,引起细胞DNA双链断裂损伤,激活细胞内因降解减少而高表达的P53蛋白,P53进而转录激活靶基因,阻滞细胞周期抑制G0/G1-S转换、阻止DNA复制抑制细胞生长增殖并启动细胞凋亡,其抑瘤作用与蛋白酪氨酸激酶作用相拮抗,并最终起主导作用。
需说明的是,本实验虽证实CUL5基因表达情况与CNE2细胞照射中后期增殖活性有关,但是其机制尚不明确。我们推测的可能性是否存在,是否有其他未知因素在起作用,结果与活体实验是否一致,确切的结论尚需更多的后续研究,如配合不同功能状态的p53及进行动物实验等来加以证实。
[1] Marcu L, Doorn T, Olver I.Modelling of post-irradiation accelerated repopulation in squamous cell carcinomas[J].Phys Med Biol, 2004, 49(16): 3767-3779.
[2] 朱小东, 曲颂, 李力, 等.细胞周期基因芯片筛选不同增殖状态下CNE-2细胞的差异表达基因[J].肿瘤预防与治疗, 2008, 21(1): 18-21.
[3] Elbashir SM, Lendeckel W, Tuschl T.RNA interference is mediated by 21- and 22-nucleotide RNAs[J].Genes Dev,2001, 15(2): 188-200.
[4] Hermens AF, Barendsen GW.Changes of cell proliferation characteristics in a rat rhabdomyosarcoma before and after x-irradiation[J].Eur J Cancer, 1965, 5(2): 173-189.
[5] 朱小东, 曲颂, 李力, 等.鼻咽癌细胞株CNE-2连续照射期间细胞增殖状况的研究[J].现代肿瘤医学, 2008, 8(16):1281-1284.
[6] Singhal S, Amin KM, Kruklitis R, et al.Alterations in cell cycle genes in early stage lung adenocarcinoma identified by expression profiling[J].Cancer Biol Ther, 2003, 2(3): 291-298.
[7] Burnatowska-Hldin MA, Kossoris JB, Van Dort CJ, et al.T47D breast cancer cell growth is inhibited by expression of VACM-1,a cul5 gene[J].Biochem Biophys Res Commun,2004, 319(3): 817-825.
[8] Baxter SS, Carlson LA, Mayer AM, et al.Granulocytic differentiation of HL-60 promyelocytic leukemia cells is associated with increased expression of Cul5[J].In Vitro Cell Dev Biol Anim, 2009, 45(5-6): 264-274.
[9] Luo K, Ehrlich E, Xiao Z, et al.Adenovirus E4orf6 assembles with Cullin5-ElonginB-ElonginC E3 ubiquitin ligase through an HIV/SIV Vif-like BC-box to regulate p53[J].FASEB J,2007, 21: 1742-1750.
[10] Hannon GJ.RNA interference[J].Nature, 2002, 418(6894):244-251.
[11] Oren M.Decision making by p53:Life, death and cancer[J].Cell Death Differ, 2003, 10(4): 431-442.
[12] 石慧英, 孙懿, 易红, 等.p53沉默对人鼻咽癌细胞株CNE2放射生物学特性的影响[J].国际病理科学与临床杂志,2008, 2(28): 97-102.
