重组人 Semaphorin3A诱导体外培养的大鼠大脑皮层神经元轴突损伤的实验研究
2010-09-20陶玉倩叶文华
陶玉倩, 田 青, 叶文华, 杨 颖, 张 赛
神经元的坏死、凋亡存在于很多神经系统疾病的病理生理过程中,既往研究表明[1],在神经元凋亡的典型形态学标志出现之前,常可观察到轴突损伤,轴突损伤对于整个神经元死亡来说可能是一个早期引发因素[2],然而在神经元死亡早期,有关轴突网络的退缩、崩解的分子生物化学机制还不清楚。Sema3A是分子量为 100kD的一种分泌型蛋白,由Luo等[3]首次从鸡脑中提取。在神经系统发育过程中,Sema3A作为一种抑制性轴突导向因子,通过诱发轴突末端生长锥崩解[3]来实现对轴突生长的定向导航作用。近年来研究表明,成年动物神经系统中也有 Sema3A的表达,尤其在神经系统遭受损伤后,例如脊髓创伤[4]、脑梗死[5]等,Sema3A与轴突再生的局限性有关[6]。而 Sema3A是否对体外培养的神经元轴突生长具有阻滞作用,还未见文献报道。本实验拟以体外培养的 SD大鼠神经元为细胞模型,观察外源性 Sema3A对于神经元轴突生长的影响,进而探讨 Sema3A在神经元死亡早期所起到的初始引发作用。
1 材料和方法
1.1 主要试剂与仪器 重组人 Sema3A/Fc融合蛋白购自 R&D System公司,溶于0.01mol/LPBS后,-20℃保存备用。胰酶、DNA酶、胰酶抑制剂、阿糖胞苷、L-多聚赖氨酸、Hoechst33342染液、MTT等均购自 Sigma公司。小鼠抗大鼠 MAP-2抗体购自 Programma公司;FITC偶联羊抗小鼠荧光二抗购自 Jackson公司。Neurobasal培养基、B27均购自 Invitrogen公司。细胞培养箱(Heraus公司)、荧光显微镜(Olympus公司)、酶标仪(Bio-TEK公司)等实验仪器均由中山大学附属第一医院神经科实验室提供。
1.2 神经元原代培养 新生 24h内的 SD大鼠,由中山大学医学院实验动物中心提供。解剖显微镜下,用显微手术镊无菌剥取大脑皮层,并剔除脑膜及血管组织,D-Hanks液冲洗后剪碎(以上操作均在冰上进行)。胰酶 (0.25mg/m l)消化,37℃,15min;离心 800rpm,1min,弃上清。加入 D-Hanks液(含 0.08mg/ml DNA酶及 0.56mg/ml胰酶抑制剂),终止消化,并吹散成悬液;静置 15min后,取上清;离心 1500rpm,5min,弃上清。Neurobasal培养液(含 1%胎牛血清、2%B27)重悬细胞,吹散为单细胞悬液。细胞计数后调整为合适的细胞密度,接种于 L-多聚赖氨酸(0.1mg/m l)预包被的培养皿中,置于 37℃,5%CO2的培养箱中培养。接种 24h后加入阿糖胞苷(10μmol)抑制胶质细胞生长,接种 3d后半量换液,以后每 3~4d半量换液一次。
1.3 神经元生长锥观察 原代神经元接种于六孔培养板,培养 24h后,进行 Sema3A(终浓度为0.5 mg/m l)处理,24h后置于倒置相差显微镜下观察。
1.4 MAP-2、Hoechst33342免疫荧光染色 原代神经元接种于放有灭菌盖玻片的六孔培养板中,制成细胞爬片。培养 3d后,进行 Sema3A处理。48h后,吸弃培养液,PBS漂洗 3次;4%多聚甲醛冰上固定 30min,PBS漂洗 3次;0.1%TritonX-100/PBS 37℃破膜 30min,PBS漂洗 3次;正常山羊血清室温封闭 30min后弃血清免洗,加入小鼠抗大鼠MAP-2一抗(1∶200,1%BSA/PBS稀释),湿盒内孵育 4℃过夜。次日吸弃一抗,PBS漂洗 3次;滴加FITC偶联的羊抗小鼠荧光二抗(1∶100,1%BSA/PBS稀释)避光室温孵育 1h,PBS漂洗后再滴加 Hoechst33342染液,染色 10m in;PBS充分漂洗后中性甘油封片。荧光显微镜下观察拍照。
1.5 轴突长度测量 神经元贴壁培养 3d后,加入终浓度为 5mg/m l的 Sema3A,连续作用48h后,MAP-2免疫荧光染色,并拍摄显微镜下荧光图像,利用 Image-Pro Plus 6.0软件进行图像分析,以轴突末端至其胞体结合部的循迹距离为尺度界定标准,测量图像中所有细胞的轴突长度。正常对照组在相同时间点进行荧光染色,并拍照测量轴突长度。两组分别随机选取 10个视野,共有 250个神经元的轴突纳入测量。
1.6 MTT法测细胞活力 原代培养神经元,调整细胞浓度为 2×104个/L,种植于 96孔培养板,培养 3d后,弃除原培养液,加入各种工作液。实验分组:正常对照组(工作液为正常培养液)、Sema3A处理组(工作液为含不同浓度 Sema3A的细胞培养液,其中 Sema3A终浓度分别为 0.1、0.5、1.0、2.5、5.0mg/ml),每组设立 6个平行复孔及 1个空白对照孔。各组工作液作用 48h后,进行 MTT检测:每孔加入 MTT 20μl(终浓度为 5mg/ml,避光),37℃,5%CO2培养箱继续培养 4h后,小心弃除上清,加入DMSO100μl,振荡 10min溶解紫色结晶物,用酶标仪检测 490nm波长处吸光度值(A490nm,OD)。两组数据均扣除空白对照孔 A490nm值后,计算细胞存活率 Survival(%)=Sema3A处理组 A490nm值/正常对照组 A490nm值 ×100%。
1.7 统计学处理 采用 SPSS 13.0统计软件分析数据,计量数据以均数±标准差(±s)表示,两组独立样本均数间比较采用 t检验,多组独立样本均数比较采用单因素方差分析,组间两两比较采用Bonferroni法,P<0.05为差异具有统计学意义。实验结果均经重复实验 3次。
2 结 果
2.1 Sema3A引起神经元生长锥崩解 倒置相差显微镜下观察,神经元贴壁培养 24h后,轴突开始由胞体发出并向外延伸。正常对照组神经元轴突末端呈扇形膨大,细小丝状分支由扇形膨大处发出并向外伸展,为正常生长锥形态(见图1A);而加入外源性 Sema3A的神经元轴突末端尖锐,扇形膨大结构消失,生长锥崩解(见图1B)。
2.2 Sema3A抑制神经元轴突生长 荧光显微镜下观察并测量神经元轴突长度。神经元贴壁培养5~6d后,观察可见对照组 MAP-2免疫阳性细胞的轴突较为粗壮、健康并且细胞之间轴突联系密切(见图2A);而 Sema3A处理组的神经元轴突较为纤细,部分细胞发生轴突退化与崩解(见图2B)。经测量对照组神经元轴突平均长度为(289.24±47.84)μm,而 Sema3A处理组的轴突平均长度显著缩短为(95.56±25.26)μm,两组比较有统计学意义(P<0.001,见图3)。表明 Sema3A对体外培养的皮层神经元的轴突生长具有明显的抑制作用。
2.3 Sema3A的神经元毒性作用 MTT法检测不同 Sema3A剂量组的神经元活性。实验结果显示,Sema3A 0.5mg/m l开始对神经元产生毒性作用,细胞存活率与正常对照组比较,降至 84.45±0.94(%);Sema3A的 IC50约为 2.5mg/ml;随着 Sema3A剂量梯度增加,细胞存活率逐渐下降(见表1)。
表1 Sema3A对神经元生存率的影响(±s,n=6)

表1 Sema3A对神经元生存率的影响(±s,n=6)
与正常对照组比较*P<0.001,△P>0.05
组别 A 490nm 生存率 (%)正常对照组Sema3A 0.1(mg/ml)Sema3A 0.5(mg/ml)Sema3A 1.0(mg/ml)Sema3A 2.5(mg/ml)Sema3A 5.0(mg/ml)0.53±0.03 0.53±0.03△0.45±0.03*0.38±0.01*0.24±0.02*0.23±0.02*100.00±0.00 98.64±6.31△84.45±0.94*70.78±1.64*45.11±2.35*43.44±2.29*

图1 倒置相差显微镜普光下生长锥观察 (×200)
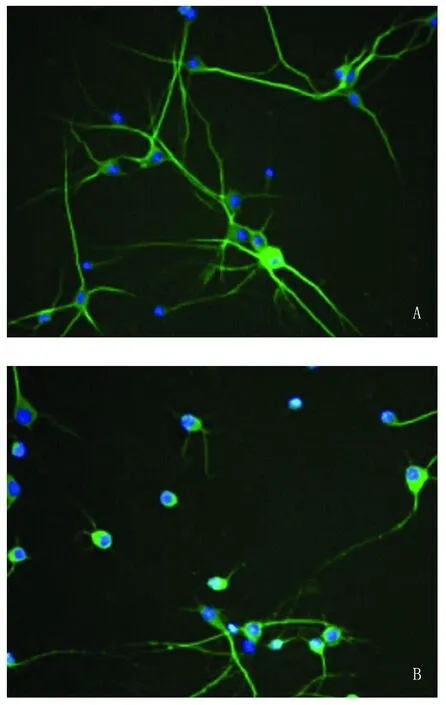
图2 MAP-2、Hoechst33342免疫荧光染色 (×200)

图3 神经元平均轴突长度测量结果(±s,n=6)
3 讨 论
在神经系统发育过程中,神经元的轴突沿着特定的路径不断延伸,直至到达将与其建立突触联系的靶点(靶细胞的胞体、树突或轴突),最终形成精密的神经网络。而这一过程的精确调节,需要轴突生长导向因子[7]的介导。在体内,Sema3A作为一种抑制性轴突导向因子,主要在胚胎期表达,在神经系统发育过程中通过介导位于轴突末端高度能动和敏感的结构-生长锥[8]的崩解来促使轴突生长方向的逆转,从而起到对轴突生长的定向导航作用[9]。而近年来研究表明,成年动物神经系统中也有 Sema3A的表达,尤其在神经系统遭受损伤后,Sema3A有一定的表达上调[4,5,10]。已有体内实验证实,中枢神经系统损伤后形成胶质瘢痕,侵入损伤核心区的脑脊膜成纤维细胞能够表达分泌型 Sema3A[11],并且外周神经损伤后,脊髓背根神经节(DRG)发出的轴突不能穿越有 Sema3A表达的瘢痕组织[12]。
既往研究已经证实,Sema3A在体内对胚胎发育中的神经元起到轴突导向的作用[9]。本次实验以体外培养的新生乳鼠大脑皮层神经元为细胞模型,予以外源性蛋白 Sema3A(终浓度 0.5mg/ml)处理 24h后,可通过倒置相差显微镜观察到神经元生长锥发生崩解,说明出生后动物的神经元仍然对于 Sema3A敏感,在体外 Sema3A对于新生动物的神经元同样具有诱导生长锥崩解的作用。
已有体内实验证明,Sema3A在中枢神经系统损伤后的胶质瘢痕中表达,起到抑制神经元轴突延伸的作用[11],而相关体外实验尚无文献报道。本次实验将体外培养的皮层神经元予以 Sema3A(终浓度 5mg/ml)处理48h,可观察到神经元轴突变得纤细,部分细胞发生轴突退化与崩解;从而在细胞水平证明了 Sema3A对于出生后动物的神经元,可发挥明显的诱发轴突损伤及抑制轴突生长的作用。另外,本实验应用不同浓度的 Sema3A处理神经元,观察到随着 Sema3A剂量梯度增加,细胞存活率逐渐下降;提示 Sema3A不仅抑制轴突生长,而且随其浓度增加也逐渐具有一定的神经元毒性。并且,已有文献报道,Sema3A在一定浓度范围内能够诱导体外培养的小脑颗粒神经元凋亡[13]。那么,Sema3A作为一种轴突导向因子,不仅在轴突导向方面发挥作用,在特定的因素诱导下,可能通过某种抑制性信号导致神经元活性降低甚至诱发细胞凋亡,而其轴突导向和细胞死亡的信号转导是否通过同一条信号通路,还需要进一步的实验证实。
通过本次实验观察,并结合以往文献报道,可以推测,由Sema3A激活的抑制性信号不仅局限于神经系统发育的早期,其在动物出生乃至成年后仍然发挥作用。在体内神经系统遭受损伤后,在病理性细胞凋亡的过程中,病灶周围处于应激状态但仍有活力的神经元或相关胶质细胞可能会分泌 Sema3A等抑制性轴突导向因子,继而被周围细胞表面 Sema3A受体复合体Neurotrophin-1/Plexin-A1(Nrp-1/PlexinA-1)[14]所识别,最终激活胞内信号通路,导致轴突损伤,甚至细胞死亡。
综上所述,在正常神经系统发育过程中,Sema3A起到重要的轴突导向作用,以确保建立精密有效的神经网络,而在成年动物神经系统损伤后,Sema3A可能通过诱导轴突损伤,对轴突再生产生消极影响,阻碍新的轴突网络的建立形成,并进一步诱发神经元损伤甚至凋亡。然而对于 Sema3A诱发轴突损伤的机制,其激活的信号转导通路如何,本实验未做深入研究。对于 Sema3A诱导神经元轴突损伤的实验研究有助于阐释 Sema3A在神经元死亡早期所起到的初始引发作用,进而为早期神经保护提供新的靶点和思路。
[1]Wakade TD,Palmer KC,Mccauley R,etal.Adenosine-induced apoptosis in chick embryonic sympathetic neurons:a new physiological role for adenosine[J].JPhysiol,1995,488(Pt 1):123-138.
[2]Raff MC,Whitmore A,Finn JT.Axonal self-destruction and neurodegeneration[J].Science,2002,296(5569):868-871.
[3]Luo Y,Raible D,Raper JA.Collapsin:a protein in brain that induces the collapse and paralysisof neuronal growth cones[J].Cell,1993,75(2):217-227.
[4]Pasterkamp RJ,Giger RJ,Ruitenberg MJ,etal.Expression of the gene encoding the chemorepellent semaphorin IIIis induced in the fibroblast component of neural scar tissue formed following injuries of adult but not neonatal CNS[J].Mol Cell Neurosci,1999,13(2):143-166.
[5]Hou ST,Keklikian A,Slinn J,etal.Sustained up-regulation of semaphorin 3A,Neuropilin1,and doublecortin expression in ischem ic mouse brain during long-term recovery[J].Biochem Biophys Res Commun,2008,367(1):109-115.
[6]Pasterkamp RJ,Verhaagen J.Semaphorins in axon regeneration:developmental guidancemolecules gone wrong[J]?Philos Trans R Soc Lond B Biol Sci,2006,361(1473):1499-1511.
[7]Chilton JK.Molecular mechanisms of axon guidance[J].Dev Biol,2006,292(1):13-24.
[8]Lowery LA,Van Vactor D.The trip of the tip:understanding the growth conemachinery[J].Nat Rev Mol Cell Biol,2009,10(5):332-343.
[9]Yazdani U,Terman JR.The semaphorins[J].Genome Biol,2006,7(3):211.
[10]Klebanov O,Nitzan A,Raz D,etal.Upregulation of Semaphorin 3A and the associated biochem ical and cellular events in a ratmodel of retinal detachment[J].Graefes Arch Clin Exp Ophthalmol,2009,247(1):73-86.
[11]Niclou SP,Franssen EH,Ehlert EM,etal.Meningeal cell-derived semaphorin 3A inhibits neurite outgrowth[J].Mol Cell Neurosci,2003,24(4):902-912.
[12]DeWinter F,Oudega M,Lankhorst AJ,etal.Injury-induced class 3 semaphorin expression in the ratspinal cord[J].Exp Neurol,2002,175(1):61-75.
[13]Shirvan A,Ziv I,Fleminger G,etal.Semaphorins as mediators of neuronal apoptosis[J].JNeurochem,1999,73(3):961-971.
[14]Takahashi T,Fournier A,Nakamura F,etal.Plexin-neuropilin-1 complexes form functional semaphorin-3A receptors[J].Cell,1999,99(1):59-69.
