不同基因型烟草品种接种赤星病菌后抗病蛋白的变化研究
2010-09-12曹祥炼宋彦君程海峰豆显武杨铁钊
曹祥炼,宋彦君,程海峰,豆显武,杨铁钊
(1.湖北省襄樊市烟草公司 保康烟叶分公司,湖北 保康 441600;2.河南农业大学 烟草学院,河南 郑州 450002)
烟草赤星病是由链格孢菌[Ternaria alternata(Fries)Keissler]引致的烟叶成熟后期重要的叶部真菌性病害,严重影响着烟草的产量和品质,制约着烟草生产的发展。赤星病在世界各烟草产区均有发生,是烟草生产上威胁最大的病害之一。赤星病具有潜育期短、流行速度快的特点,在环境条件有利于发病的情况下,短时间内即可造成大的流行,给烟叶生产带来巨大损失[1]。烟草赤星病1892年首次在美国发现,曾几次给世界烟叶生产带来巨大的经济损失[2]。1916年我国在北京附近首次发现,1964年在山东烟区流行,随后全国各烟区有日益严重的趋势。1989至1991年3年间,河南省平均每年因赤星病造成的损失达人民币2 300万元[3],成为毁灭性的病害。2000年8月初在美国的康乃狄克州和麻萨诸塞州,烟草赤星病大爆发,分别减产至少达75% 和89%[4]。
植物受到病原真菌侵染时,会产生一系列的主动防御反应。这些反应包括:合成植保素,感染区域细胞结构的加厚与增强,病程相关蛋白 (PR蛋白)的诱导和积累。这些反应还参与了防卫反应中的信号传递,使植物获得系统性抗性[5-6]。
病程相关蛋白中研究较多的是几丁质酶(Glu)和β-1,3-葡聚糖酶 (Cht)。很多植物病原真菌细胞壁的主要成分是几丁质和葡聚糖,体外抑菌试验表明,它们能抑制一些病原真菌孢子的萌发和病菌生长,因此几丁质酶和葡聚糖酶一直被看作是植物抗真菌病害的潜在物质[5]。
1 材料与方法
1.1 试验材料与种植
供试烟草品种为感赤星病品种长脖黄和高抗赤星病突变体净叶黄,豫烟4号作感病对照,烟草品种来自河南省农业科学院。
试验地点设在河南农业大学科教园区。种子、植烟土壤经过严格消毒,采用盆栽种植。3个品种分别于2008年5月10号移栽,重复3次,随机区组排列,正常管理。
1.2 菌种与培养
孢子悬浮液。
1.3 接种与取样
从烟草苗下部起选第3到第4片成熟叶片,喷雾接种,每个小区的各品种处理一致。接种后人工保温30℃并保湿培养,分别于接种前0 h,接种后12,24,36和48 h取样。以同生育期同部位成熟叶片喷施清水做空白对照。
田间采集到的赤星病菌强毒株在PDA培养基上培养7 d后,用无菌水洗下,配制成孢子浓度为106个·mL-1的孢子悬浮液。
1.4 测定方法
几丁质酶活性的测定参照 Boller等[7]的方法。β-1,3-葡聚糖酶活性的测定参照史益敏[8]的方法。
2 结果与分析
2.1 几丁质酶活性
几丁质是许多真菌细胞壁、昆虫体壁和甲壳类动物中的主要结构成分,但在高等植物中尚未发现有几丁质的存在。当植物受到病原物侵染时体内会积累病程相关蛋白 (pathogenesis-relatedprotein,PR),已证明部分PR是几丁质酶[10]。很多研究表明,几丁质酶在参与寄主植物和病原菌之间相互作用有2个方面,即几丁质酶有潜在的抗菌特性的直接作用和通过释放几丁质寡聚物而诱导植物产生抗性的间接作用[11]。现已有大量的试验证实,植物几丁质酶是可以被诱导的,大多数高等植物体内所含的几丁质基因表达水平很低或不表达,而在受到如病原菌侵染、外源几丁质、壳聚糖、水杨酸和乙烯等因子作用时均能被诱导表达[12]。
测定不同基因型的烟草品种接种赤星病菌后在正常生长条件下其叶片内几丁质酶的基础活性,结果如图1所示:抗病品种净叶黄几丁质酶总活性的平均值为7个活力单位 (U),感病品种长脖黄为3个活力单位,而另一感病品种豫烟4号的几丁质酶的基础活性为4个活力单位左右。由此可见在正常生长条件下,抗病品种净叶黄比感病品种长脖黄和豫烟4号体内几丁质酶基础活性高。

图1 抗性烟草品种未接种几丁质酶的变化
从图2-4可以看出,无论是抗病品种还是感病品种,接种赤星病菌后几丁质酶含量与其基础含量相比都呈现明显的上升趋势。3个品种在接种赤星病菌后的前12 h几丁质酶活性均上升较慢,净叶黄在接种后12~24 h几丁质酶含量达到10.06个活力单位,此后几丁质酶活性仍一直上升,但升高速度减慢。感病品种长脖黄和豫烟4号几丁质酶活性变化规律基本一致,在接种赤星病菌后48 h内酶活性呈上升趋势。几丁质酶潜在的抗菌特性的直接作用导致酶活性在接种前期剧烈上升,之后通过释放几丁质寡聚物而诱导植物产生抗性的间接作用,从而导致几丁质酶活性的在接菌后的48 h内一直上升。从图5可以看出,不同抗性烟草品种在接种赤星病菌后的前12 h,抗病品种净叶黄几丁质酶活性上升速度比感病品种长脖黄和豫烟4号要快,且在整个过程中净叶黄的几丁质酶活性都比长脖黄和豫烟4号要高,这可能是净叶黄抗赤星病的原因。

图2 净叶黄接种后几丁质酶的变化
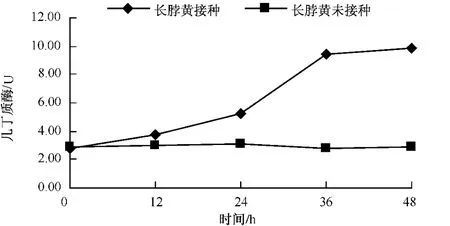
图3 长脖黄接种后几丁质酶的变化

图4 豫烟4号接种后几丁质酶的变化

图5 抗性烟草品种接种后几丁质酶的变化
2.2 β-1,3-葡聚糖酶活性
测定不同基因型的烟草品种接种赤星病菌后,在正常生长条件下其叶片内β-1,3-葡聚糖酶的基础活性,结果如图6所示:不同基因型烟草品种β-1,3-葡聚糖酶活性变化与几丁质酶基础活性变化基本一致,都表现为长脖黄酶基础活性最低,净叶黄基础活性最高,豫烟4号居中。抗病品种净叶黄在正常生长条件其β-1,3-葡聚糖酶基础活性的平均值为1 950个活力单位,感病品种长脖黄基础活性为1 700个活力单位,而另一感病品种豫烟4号为1 850个活力单位上下。由此可见在正常生长条件下,抗性品种净叶黄比感病品种长脖黄体和豫烟4号体内几丁质酶的活性高。
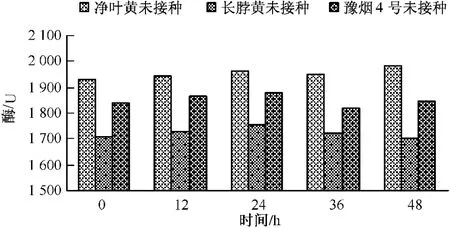
图6 抗性烟草品种β-1,3-葡聚糖酶活性的变化
测定不同品种在接种赤星病菌后β-1,3-葡聚糖酶酶活性的变化,结果如图7-9所示,无论是抗病品种还是感病品种,接种赤星病菌后β-1,3-葡聚糖酶含量与其基础含量相比都呈现明显的上升趋势。净叶黄在接菌后12 h内 β-1,3-葡聚糖酶达到2 272.4个活力单位,此后 β-1,3-葡聚糖酶活性仍一直上升,但升高速度减慢。这可能与葡聚糖在细胞壁内部外部均存在有关。感病品种长脖黄和豫烟4号β-1,3-葡聚糖酶活性变化规律基本一致,在接种赤星病菌48 h内酶活性呈上升趋势。从图10可以看出,不同抗性烟草品种在接种赤星病菌后的前12 h,抗病品种净叶黄β-1,3-葡聚糖酶活性上升速度比感病品种长脖黄和豫烟4号要快,而感病品种长脖黄和豫烟4号在接种后12~24 h β-1,3-葡聚糖酶酶活性上升较快,但比抗病品种晚12 h。在整个过程中净叶黄的β-1,3-葡聚糖酶活性都比长脖黄和豫烟4号要高,这可能是净叶黄抗赤星病的原因。

图7 净叶黄接种后β-1,3-葡聚糖酶活性的变化

图8 长脖黄接种β-1,3-葡聚糖酶活性的变化

图9 豫烟4号接种β-1,3-葡聚糖酶活性的变化
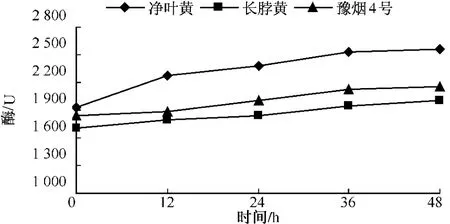
图10 不同抗性品种β-1,3-葡聚糖酶活性的变化
2.3 几丁质酶和β-1,3-葡聚糖酶活性变化趋势
从图11可以看出,净叶黄在接菌后 β-1,3-葡聚糖酶变化比较平稳,几丁质酶在接菌后12 h剧烈上升,24~48 h缓慢上升;感病品种长脖黄在接菌后,β-1,3-葡聚糖酶和几丁质酶活性变化趋势基本一致,在整个过程中一直呈上升趋势,几丁质酶在24~36 h剧烈上升,在36 h上升缓慢 (图12);豫烟4号 β-1,3-葡聚糖酶活性在12~36 h上升明显,几丁质酶酶活性在24~36 h明显增强(图13)。从时间上看感病品种长脖黄和豫烟4号几丁质酶酶活性剧烈上升比抗病品种净叶黄晚12 h。3个品种在接菌后几丁质酶和 β-1,3-葡聚糖酶酶活性均表现一定的相似性,几丁质酶酶活性剧烈上升时间比β-1,3-葡聚糖酶晚。分析认为,可能与β-1,3-葡聚糖在病菌细胞壁内部外部都存在,而几丁质主要存在与病菌的细胞壁内部有关。
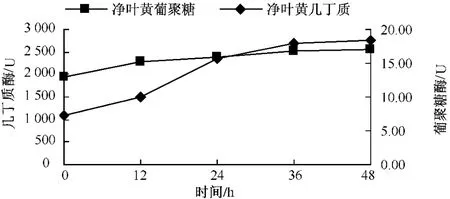
图11 净叶黄接种后几丁质酶和β-1,3-葡聚糖酶酶活性的变化
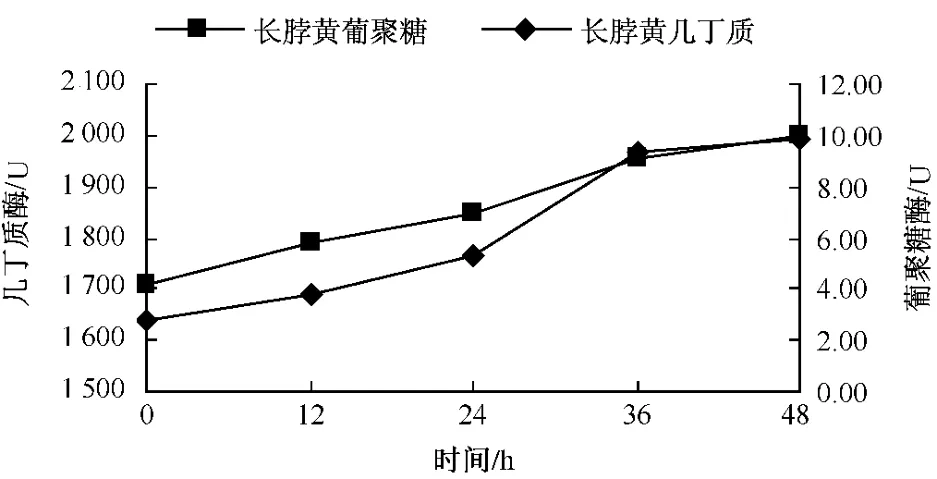
图12 长脖黄接种后几丁质酶和β-1,3-葡聚糖酶酶活性的变化

图13 豫烟4号接种后几丁质酶和β-1,3-葡聚糖酶酶活性的变化
3 小结与讨论
几丁质和β-1,3-葡聚糖是真菌细胞壁的重要结构成分 ,在许多真菌的菌丝顶端 ,β-1,3-葡聚糖和几丁质暴露在细胞壁表面 ,能够直接受到β-1,3-葡聚糖酶和几丁质酶的水解[14],这不仅使真菌菌丝生长点受到破坏 ,而且在水解过程中由真菌细胞壁释放出来的寡糖能够作为植物多种抗病反应的激发因子 ,诱导植物的全面防卫反应[14]。在正常生长条件下,抗病品种净叶黄比感病品种长脖黄和豫烟4号体内几丁质酶基础活性高,接种赤星病菌后抗病品种净叶黄几丁质酶活性上升速度比感病品种长脖黄和豫烟4号要快。净叶黄在接种后12 h几丁质酶活性迅速升高,在之后的24 h几丁质酶活性又一次剧烈上升。可能由于净叶黄在赤星病菌接种初期,直接攻击病原物真菌的细胞壁,使病原菌瓦解失去致病力,同时酶解后病原菌的细胞壁破碎物又可作为激发子再次刺激植物产生抗病防卫反应,导致酶活性又一次升高,从而达到抗病的目的。
在接种后的整个反应过程中长脖黄和豫烟4号的几丁质酶活性与其对照相比都有不明显的提高,但比净叶黄的要低得多。分析认为赤星病菌侵染感病品种长脖黄和豫烟4号后,由于烟草体内被诱导的几丁质酶的活性很低,抵抗和杀死病菌的能力差,是导致感病品种发病的原因之一。
无论是抗病品种还是感病品种,接种赤星病菌后β-1,3-葡聚糖酶含量与其基础含量比都有明显的上升。推断由于接种后,随着赤星病菌的入侵,烟草启动体内有关的抗病防卫机制,相关抗病蛋白酶被激活。由于β-1,3-葡聚糖酶直接作用于赤星病菌的细胞壁,因此酶活性的升高对赤星病菌起到抑制作用。净叶黄的β-1,3-葡聚糖酶的积累不管是在量上还是在时间上都比长脖黄和豫烟4号要快,对于病原菌的入侵净叶黄反应较迅速,能及时抑制病原菌,阻止病害进一步扩展。从实验结果可以看出,3个品种几丁质酶积累时间与β-1,3-葡聚糖酶积累时间相比具有一定滞后性,可能与葡聚糖在真菌细胞壁内外都有,而几丁质主要存在于细胞壁内部有关。
[1]Shew H D,Lucas G B.Compendium of tobacco disease[J].The American Phytopatho logical Society,1990,25:10-12.
[2]Stavely R,Echaplin J E,Gwynn G R.Registration of Bel 921 brown spot resistant flue-cured tobacco germplasm[J].Crop Science,1984,24:830-831.
[3]谈文.烟草赤星病的发病规律及综合治理[J].烟草科技,1993(2):45-48.
[4]Lamondia J A.Outbreak of brown spot of tobacco caused by alternaria alternata in connecticut and massachusetts[J].Plant Disease,2001,85(2):230.
[5]Kunze I,Nilsson C,Adler k,et al.Correct targeting of a vacudar tobacco chitinase in saccharomyces cerevisice posttranslational modifications are dependent on the rain[J].Biophys Acta,1998,1395(3):329.
[6]Lamb C J.Signals and transduction mechanismsfor activation of plant defeises against microbial attack[J]. Cell,1989,56:215.
[7]Boller T,Gehria A,Mauch F,et al.Chitinase in bean leaves:induction by ethylene,purification,properties, and possible function[J].Planta,1983,157:22 -31.
[8]史益敏.β-1,3-葡聚糖酶活性的测定[G]//中国科学院上海植物生理研究所.现代植物生理学实验指南.北京:科学出版社,1999:128.
[9]Boller T. Chitinase in bean:induction by ethylene,purification,properties and possible function[J]. Plant,1983,157(1):22-31.
[10]赵蕾,汪天虹.几丁质、壳聚糖在植物保护中的研究与应用进展[J].植物生理学通讯,1994,30(4):263-266.
[11]Benhamou N.菜豆内切几丁质酶对立枯丝核菌的抗菌作用[J].微生物学杂志,1994(14):66-70.
[12]王维荣,裴真明,欧阳光察.几种因子对黄瓜幼苗几丁质酶的诱导作用[J].微生物学杂志,1994(14):66-70.
[13]Bonfante P, Perotto S, Teata B, etal. Ultrasturctural localization of cell surface sugar residues in ericoid mycowhizal fungi by gold lLabeled lectina[J].Protoplasma,2987,137:25-35.
[14]Selitrennikoff C P.Antifungal proteins[J].Apple Environ Microbiol,2001,67:2883-2894.
