紫菜属海藻性别表现特征研究
2010-08-06许璞
许 璞
(常熟理工学院 生物与食品工程学院,江苏 常熟 215500)
自1862年Thuret报道了紫菜属海藻Porphyra umbilicalis与P.leucosticta精子囊的观察结果以来,对于紫菜生殖器官形成及其性别的研究基本上为描述性的.紫菜的性别表现依据分类学描述可基本分为:雌雄同株型,如条斑紫菜(P.yezoensis);雌雄异株型,如拟线形紫菜(P.pseudolinearies)和狭叶紫菜(P.angusta);雌雄同株兼有雌雄异株(或雌雄异株兼有雌雄同株)类型,如华北半叶紫菜(P.katadai var.hemiphylla)和坛紫菜(P.haita⁃nensis).作为分类学的重要依据,雌雄生殖细胞在藻体上的分布规律代表紫菜属物种的特征.在雌雄同株类型中,有的种类分裂为小区域间生或混生,而有的种类雌雄细胞分别位于同株藻体的两侧或上下两部分.条斑紫菜精母细胞分裂成熟后形成条带状精子囊器区域,镶嵌在果孢子囊群中,成为该物种的鲜明特征.然而同为生殖细胞小区域混生的种类,有的物种精子囊器区域较小,且较分散.而少精紫菜(P.oligospermatangia)成熟藻体的外型更象全雌个体,雄性细胞分裂区域极小,精子囊器区呈很小的点状出现于果孢子囊群中.半叶紫菜藻体在尚未成熟的时期就可区分出雌雄区域,两性区域之间有明显的界线,分生于藻体的两侧.雌雄同株的坛紫菜在早期生长阶段难以分辨两性区域,当藻体成熟时,果孢子囊群和精子囊器常以直线或曲线区别,各自分布于藻体的不同区域.雌雄异株的藻体,生殖细胞多沿藻体的前端和两侧边缘形成.日本学者对紫菜生殖细胞分布类型做了一个近似的归纳总结:雌雄异株,即藻体只产生单一性别生殖细胞而有雌、雄藻株之分;雌雄同株,同一藻体上发生雌、雄生殖细胞,但两性生殖组织区域分生明显;雌雄同株,两性生殖细胞分裂成小区域间生或混生在一起;雌雄同株,雄性生殖区域极小或不明显[1].
紫菜的性别发生有什么规律,为什么会出现如此多样的性别表现类型,它们是性决定还是性分化?随着紫菜减数分裂发生在壳孢子萌发时期的新证据出现[2-4],遗传学研究者认为性别遗传机制可以阐明这一现象.上世纪90年代初,加拿大藻类遗传学家ven der Meer和他的学生Mitman,对一种生长分布在北美地区的紫菜P.purpurea开展了性别遗传及其决定机制的研究,其主要结果包括:P.purpurea减数分裂发生在壳孢子萌发之时,被突变色素标记的萌发细胞发育历程表明,P.purpurea藻体形成的两性生殖细胞区域,起源于壳孢子最初的分裂细胞,一个细胞仅对应于一种性别的细胞组织区域;P.purpurea藻体两性生殖区域分生特征是由减数分裂第一次分裂后,处于顶端位置细胞第二次分裂所作横向分裂所致,即这一雌雄同株型藻体的性别表现出分界明晰的左右性,是由早期细胞分裂生长模式所决定;对野外生长和实验培养的藻体所作的性别比例分析,其结果符合1∶1分离规律[5].这一研究结果说明,P.purpurea的性别发生受一对等位基因控制,在壳孢子初始萌发分裂时,性等位基因随减数分裂分离,表现两性的初始细胞在同一个体上各自分别生长发育,长成雌雄组织的嵌合体,表现为雌雄同株、两性区域分生的藻体.
对于P.purpurea性决定机制的研究,是首次对紫菜性别发生机制的研究工作.由于紫菜属海藻在性别发生上的多样性,仅由性决定机制解释显然是不够的.华北半叶紫菜(P.katadai var.hemiphylla)、少精紫菜(P.oligo⁃spermatangia)、坛紫菜(P.haitanensis)和条斑紫菜(P.yezoensis)为我国沿海常见紫菜物种,其中坛紫菜和条斑紫菜也是人工栽培利用的种类,属重要经济海藻.4种紫菜在生殖器官发生以及性别表现上有明显差异[6],对这些紫菜种类表现的性别特征进行比较研究,有助于认识紫菜属海藻性别发生的规律.
1 材料与方法
1.1 藻体性别观察
华北半叶紫菜:每年春季3~4月青岛海区自然发生群体达到盛期,对数量较大的藻落进行采集,藻体长3~10cm.采集藻体保湿带回实验室,逐一记录检测结果.
坛紫菜:对不同区域自然生长和人工栽培生长群体开展多批次取样,统计样本数量>200株/次,每一藻株除外观检测外还作镜检验证,以保证统计分析的可靠性.野外取样开展进一步的实验培养观察,包括成熟藻株和未成熟藻株,培养条件:20±1℃,100μmol m-2s-1,14L:10D,10~25d.
条斑紫菜和少精紫菜:每年春季采集青岛海区成熟藻体,条斑紫菜还包括江苏沿海人工栽培的藻体,采集藻体的检测结果再由镜检逐一确认.
1.2 藻体早期发育模式和生长特点的观测
色素突变体诱变获得方法参照有关文献[7].采集突变型丝状体成熟释放的壳孢子,玻片水滴培养,显微镜观测记录.重点观察比较各物种壳孢子萌发个体的早期生长特点和色素突变体标记的细胞组织生长发育过程,研究区分各物种藻体的发育特征以及与性别表现的关系.
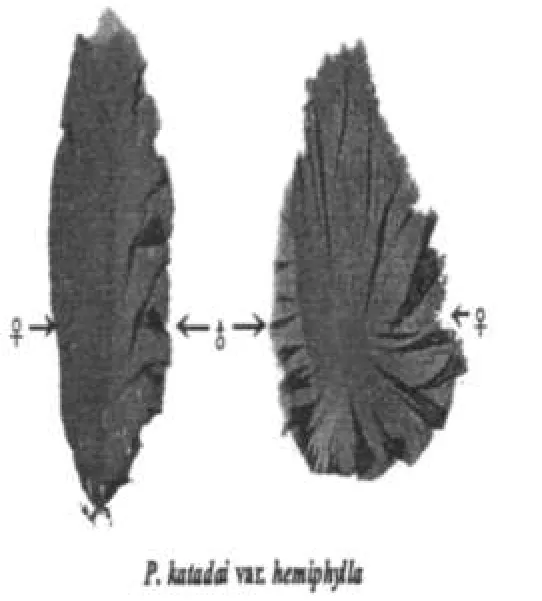
图1 华北半叶紫菜性别表现特征
2 结果
2.1 4种紫菜性别表现特征与性比观测
2.1.1 华北半叶紫菜性别表现特征与性比
华北半叶紫菜在青岛沿岸海域春季发生,3~4月为生长盛期.野外生长的植株数毫米大小就可镜检观察到色泽深浅分明的两种藻体组织,一般以藻体生长中轴为界,两侧分生.藻体成熟时,色泽较深区域细胞发育为果胞和果孢子囊,色泽较浅部分形成精子囊器.其植株虽为雌雄同株,但性别的两侧分生特征明显(见图1).
对2005年、2006年生长盛期采集的所有3cm以上植株进行性别观察分析,获得3481株,其中雌雄同株3356株,雌株68株,雄株57株,雌、雄性别比例为1∶0.997(表1).这一结果表明华北半叶紫菜符合等位性状分离的数量关系.
2.1.2 坛紫菜性别表现特征与性比
坛紫菜藻体秋季发生,在主产地福建海域的自然生长群体初冬进入生长盛期,翌年春季衰老流失.人工栽培藻体生长旺盛期较早,闽浙粤海区稍有差异,一般为10~12月,但生长群体衰老时间也较早于当地的野生群体.坛紫菜藻体进入快速生长期后,不同时期均可采集到成熟植株.成熟植株发生的生殖细胞多沿藻体前端边缘分布,充分成熟的藻体前端部充满雌或雄的生殖器官,并延伸至藻体中、后部位的边缘区域.同一植株一般表现一种性别(图2A),但仍可观察到雌雄性别同株的藻体,雌雄同株藻体两性生殖细胞有各自的分布区域,以曲线、直线或斜线分界(图2B).
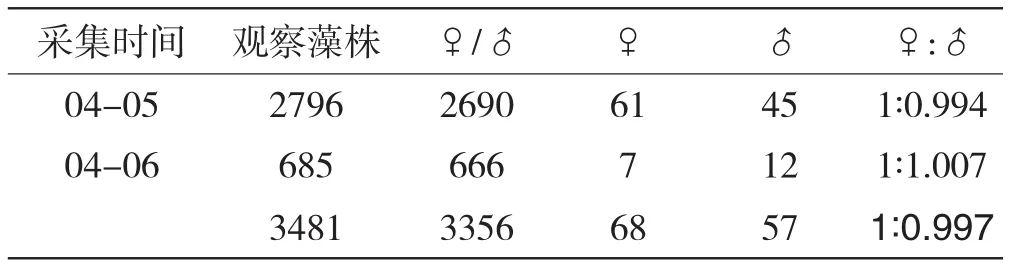
表1 半叶紫菜自然种群藻体性比观测
采集2004~2006年不同生长期的藻株材料进行观察,总数达到6411株,获得成熟植株5050株,其中表现雌性的植株2455株,雄株2581株,雌雄同株14株,性别比例1∶1.05(表2).观察结果显示,不同批次采集的生长群体性别比例有所差异,但无偏性规律,随着观察样本的增加,其性别比例接近1∶1数量关系.
2.1.3 坛紫菜性别发生特点的实验观察
在野外采集的观察材料中,每批都有不同比例的未成熟藻体(表2).随机取用部分未成熟藻体(组别1、组别3)进行室内培养,继续观察其性别发生特点.结果显示:较短培养时间(11~12d)达到性成熟的藻体不足半数,其中表现雄性性别的植株数明显多于雌性;经较长时间培养(25d)后,多数藻体可以成熟,雌雄植株比例相等(表3).这一结果表明,在同一群体内雄性生殖细胞发生较早,可能影响性别比例的观察结果,但充分成熟的藻体性比仍然具有1∶1关系,印证了野外生长群体的观察结果.

图2 坛紫菜性别表现特征
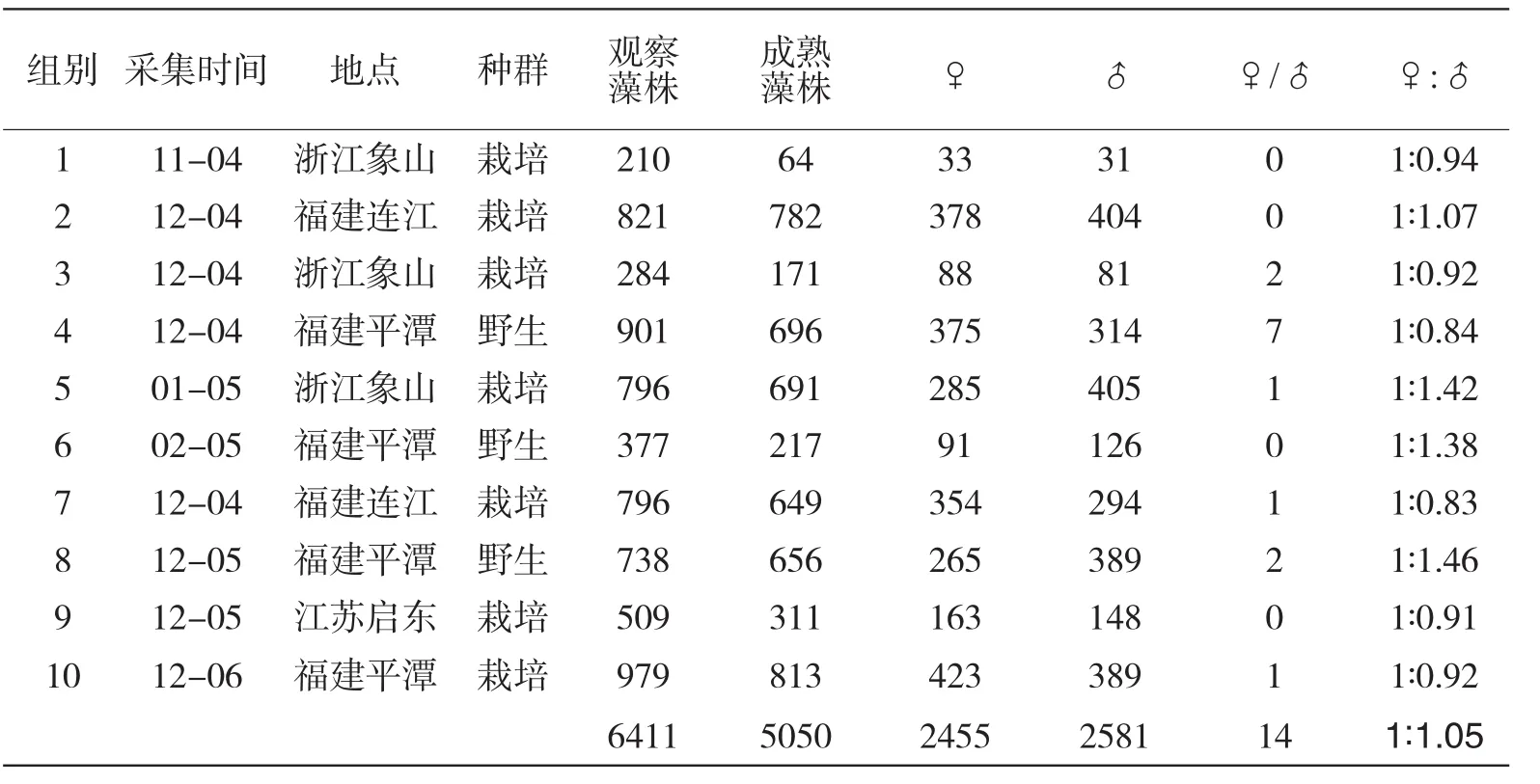
表2 坛紫菜野外生长藻体性比观测

表3 坛紫菜野外未成熟藻体实验培养的性比观测
继续对野外已成熟的藻体进行定株培养观察(组别1、组别2、组别3、组别4、组别7和组别8),发现经一段培养时间(11~24d)后,随着藻株的充分成熟,生殖细胞向藻体中、后部位延伸,或前端部位生殖细胞大量放散离体,两种性别培养植株都出现表现相反性别性状的个体,并出现雌雄同株个体.在培养观察的1818株藻体中,异性生殖组织出现率为2.1%(表4).
2.1.4 条斑紫菜和少精紫菜性别表现及其特征观察
条斑紫菜和少精紫菜自然生长藻体均在春季发生,生长盛期一般为3~4月,少精紫菜旺盛生长期来得略迟.人工栽培的条斑紫菜秋季发生,11月进入生长盛期,直至翌年4月.两种紫菜进入快速生长期均可观察到成熟藻体.成熟的条斑紫菜藻体前端及周缘形成很多条纹状精子囊区域,镶嵌在果胞或果孢子囊区,成为该物种鲜明的特征;少精紫菜成熟初期不易观察到雄性生殖细胞,但镜检可观察到小块的雄性生殖细胞区域,充分成熟的藻体在果孢子囊区域中呈现点状或小块状精子囊器区(图3).
两种紫菜性别表现的共同特征是,无论是壳孢子或单孢子萌发生成的藻体,从未观察到单一性别表现的植株,两性生殖细胞总是在同一植株上发生,生殖器官形成区域间生或混生,无性别组织分生现象,性别表现无可量的等位关系.
2.2 4种紫菜藻体早期发育特点
由突变色素标记的杂合丝状体(2n)成熟后释放壳孢子,其萌发分裂个体显示遗传学机制的四分子特点(图4).表明紫菜叶状体是具有四分子性质的特殊生长体,由于壳孢子分裂形成的4个配子生长体不能自由分离,因而其发生植株是具有双亲背景或不同遗传性质藻体组织的嵌合体.
在所观察的4种紫菜物种中,突变色素标记的嵌合体显示,条斑紫菜、坛紫菜和少精紫菜具直线生长发育特性;华北半叶紫菜绝大多数初期分裂个体为典型的四分子缺失型,由于其第二次分裂顶端细胞分裂的横向性,致使生长个体表现出两侧发育的特性.继续培养观察,可以清晰地看到两种早期发育模式对藻体生长及形态建成的影响作用(图5).
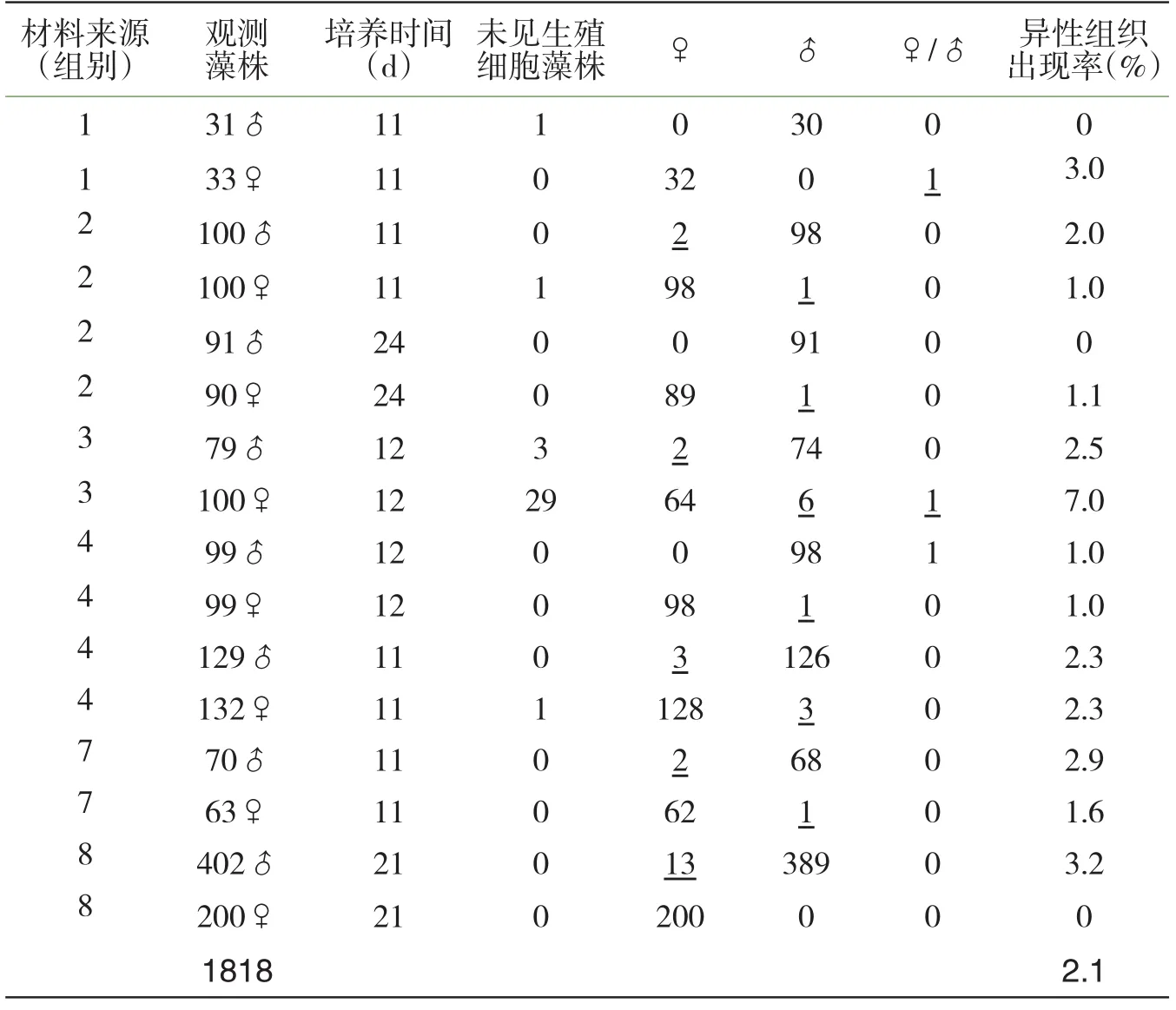
表4 坛紫菜野外成熟藻体实验培养的性比观测
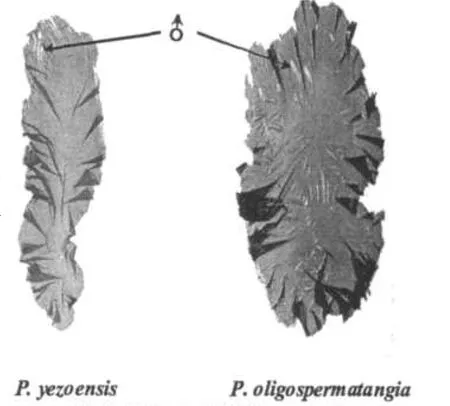
图3 条斑紫菜、少精紫菜性别表现特征
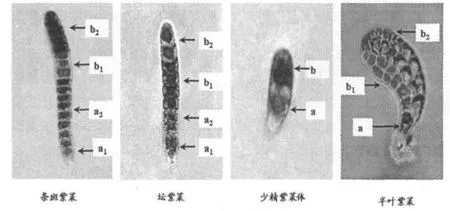
图4 紫菜壳孢子萌发分裂表现的四分子
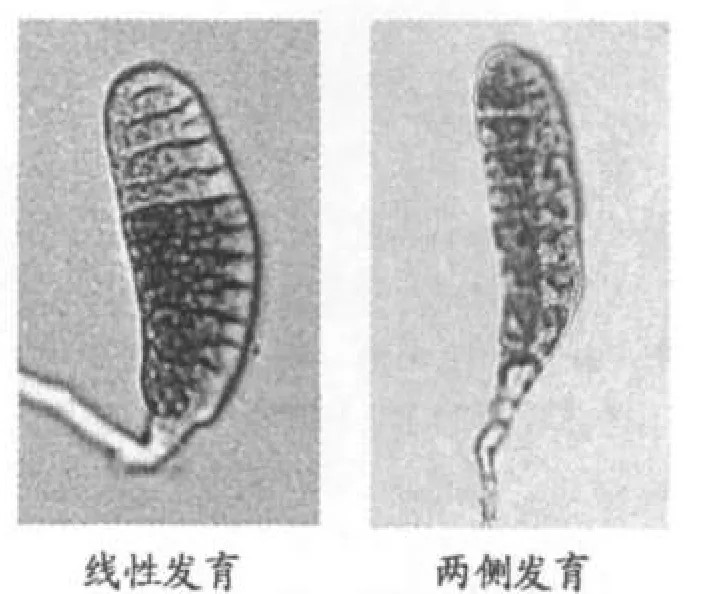
图5 紫菜生长藻体两种基本发育模式
2.3 4种紫菜性别表现特征比较
综合野外调查和实验观察结果,4种紫菜的性别表现可由表5概括.经比较表明:在紫菜物种中有两种基本的性别类型,可由性比等位关系与否加以区别雌、雄同株性别表现在不同类型的物种中,其表现方式与机制不同,具性比等位关系的物种,生长藻体(华北半叶紫菜和坛紫菜)的性别单独表达,或以分生组织表达,而两性生殖细胞间生(混生)的藻体,其物种(条斑紫菜和少精紫菜)性别无性比等位关系.
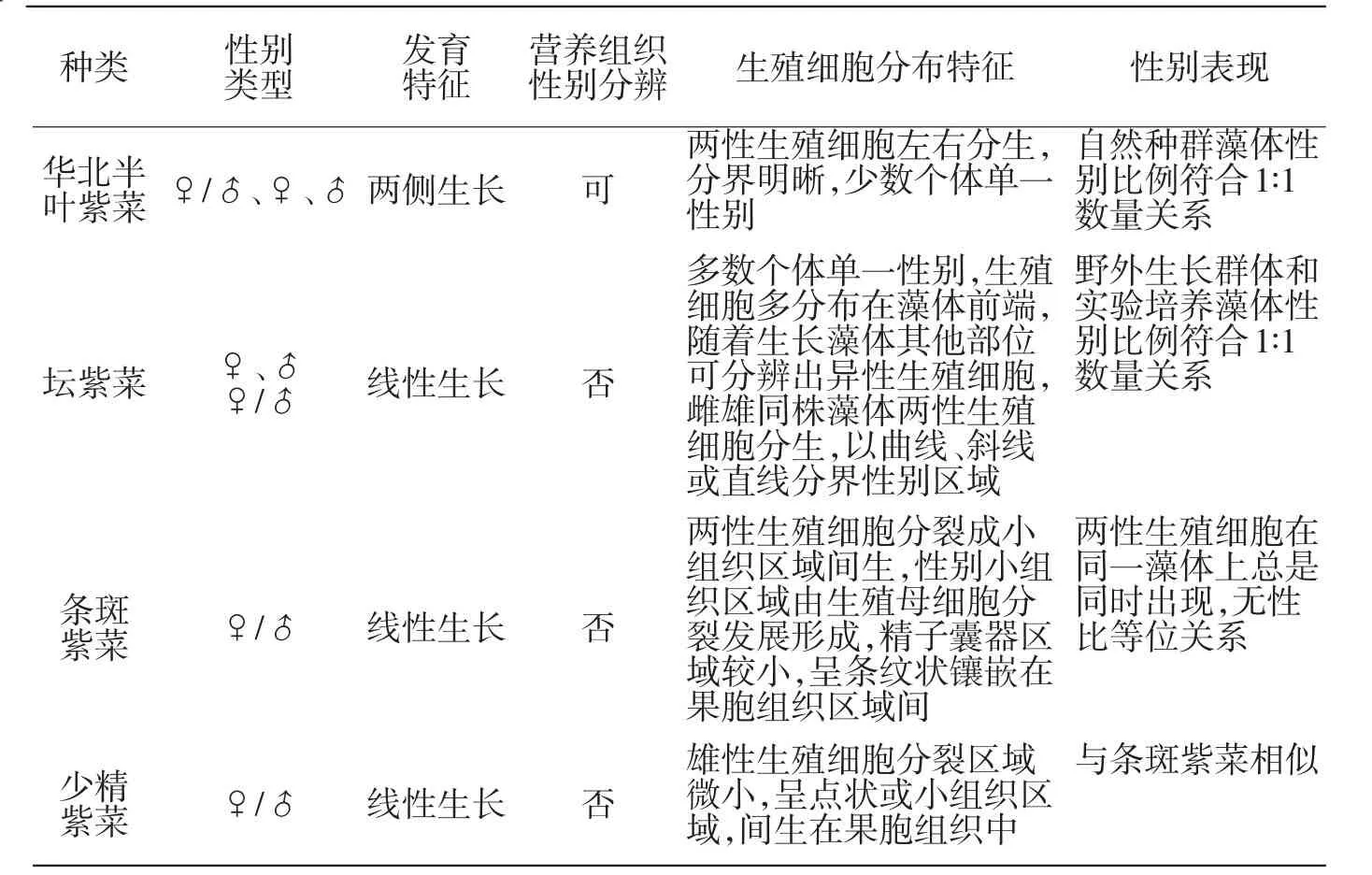
4种紫菜性别特征比较
3 讨论
在我国产的4种紫菜属海藻中,华北半叶紫菜的性别特征与P.purpurea极为相似[5].而同具性别等位分离性质的坛紫菜,尽管在一些重要的生物学特征上与华北半叶紫菜、P.purpurea相同,例如有等同的染色体数[8],但在性别表现上却有着重要差异,这也是在紫菜属海藻性别认识中容易混淆的重要问题之一.以P.purpurea为代表物种的性别表现特征是,早期发育模式具两侧生长性,性别区域主要由壳孢子减数分裂第二次分裂形成的顶端2细胞发育而来,其雌、雄性别同株的性质,是由很高的第二次分裂分离频率决定的[5].由于这一类型紫菜植株的营养组织可以识别雌雄性别,明晰的组织区域标记使其生长发育特点以及性别决定机制被直观地表现出来.从坛紫菜生长发育的观察结果看,其植株发生主要以线性生长为主,由突变色素标记的生长个体显示线性四分子特征(图4).野生型藻体在生长过程中无营养组织区域标记可识别,仅以藻体成熟部位特别是藻体前端部分发生的生殖细胞区分植株性别,容易在观察中得到雌雄异株的结果.继续生长的藻体,特别是雌雄同株藻体,显示坛紫菜藻体性别表现有先后的时序,这与其藻体的线性生长特点相关.野外采集生长充分但未成熟藻体、以及已成熟藻体的实验观察结果,可以印证这一结论.
条斑紫菜和少精紫菜显示出与性决定机制不相符合的性别表现特征.从细胞染色体数[8]、无性生殖行为[9]等生物学特征来看,这似乎也表明它们代表的是另一类型的紫菜种类.然而,即使可能同属性分化的物种,其分化的表现形式也有差异.条斑紫菜两性生殖细胞区域作较有规律的间生分布,而少精紫菜雄性细胞区域微小,只在雌性生殖细胞群中作点状或小块状分布.生殖细胞区域的大小,可以由两性生殖母细胞分化的早晚、分裂次数以及生殖器官的结构与形状等特征决定,但若两性生殖母细胞的形成是相同体细胞分裂的结果,其性别分化一定与某种调控规律有关.
[1]岩琦.浅海养殖的进展[M]//今井丈夫.浅海完全养殖.东京:株式会社恒星社厚生阁,1973:1-49.
[2]马家海,三浦昭雄.スサビノリ殻胞子とその发芽体における核分裂の观察[J].Jap J Phycol,1984,32(4):373-378.
[3]孙爱淑,曾呈奎.紫菜属的细胞学研究—膨大细胞和壳孢子萌发核分裂的观察[J].海洋与湖沼,1987,18(4):328-332.
[4]Burzycki G,Waaland J.On the position of meiosis in the life history of Porphyra torta(Rhodophyta)[J].Bot Mar,1987,30:5-10.
[5]Mitman G,van der Meer J.Meiosis,blade development,and sex determination in Porphyra purpurea(Rhodophyta)[J].J Phycol,1994,30:147-159.
[6]Tseng CK.Common seaweeds of China[M].Beijing:Science Press,1983.
[7]Wang JF,Xu P,Zhu JY,et al.The Characterization of Color Mutations in Bangiaceae(Bangiales,Rhodophyta)[J].Appl Phycol,2008,20:499-504.
[8]曾呈奎,张德瑞.紫菜的研究I.甘紫菜的生活史[J].植物学报,1954,(3):287-302.
[9]Tseng CK,Sun AS.Studies on the alternation of the nuclear phases and chromosome numbers in the life history some species of Porphyra from China[J].Bot Mar,1989,32:1-8.
