北京地区枣疯病植原体16S rDNA基因序列分析
2010-07-09林文力牟海青廖晓兰
林文力,牟海青,廖晓兰
(1.湖南农业大学生物安全科学技术学院,湖南 长沙 410128;2.中国检验检疫科学研究院动植物检疫研究所,北京 100029)
植原体(Phytoplasma),原称类菌原体(Mycoplasma-like orpanism,简称MLO)是一类无细胞壁、存在于植物筛管内的专性寄生细菌,主要依靠叶蝉和飞虱等从韧皮部取食的昆虫传播,也可由菟丝子和人工嫁接传病,常引起植物从枝、黄化、花变叶、顶枯等症状[1]。综合世界各地的报道,植原体可引起多达1 000余种植物的系统性病害,涵盖各类农作物、园艺、花卉、果树和林木等病害。其中,我国报道的有100多种,给经济作物造成了巨大损失[2]。由于植原体尚不能进行体外培养,故对其生物学特性研究较少。近年来,随着分子生物学的迅速发展,核酸杂交、血清学和PCR等技术的广泛应用,极大改善了研究者对植原体鉴定和分类的手段[3]。根据植原体通用引物和特异性引物可以特异地扩增出16S rDNA基因序列,这是植原体分类的重要参考依据,也是被国际上普遍接受的植原体鉴定分类方法[4]。
枣树在我国广泛栽培,遍布全国各地,是一种重要的经济作物。枣疯病(Jujube witches′-broom,JWB)是由植原体引起的一类病害,枣树一旦患病,通常在2~3 a内丧失产量并很快导致植株死亡。1981年,王祈楷等通过系统比较,确定了枣疯病是由植原体侵染所引起的,而不是病毒类或者病毒与植原体的复合侵染所致。枣疯病发病过程从外观上看可分为以下几个阶段:健康叶片→花叶→变态花蕾→花变叶→丛枝[5]。
本研究对我国北京地区枣疯病植原体16S rDNA基因进行PCR扩增及序列测定,并与已知的植原体序列构建系统发育进化树,明确北京地区枣疯病的分类地位,以期为枣疯病植原体的遗传变异、致病机理等研究提供参考依据。
1 材料与方法
1.1 材料
发病枣树样品采自北京昌平区西峰山村小枣林。克隆载体PMD18-T、限制性内切酶及其它酶类等分子生物学试剂均购自TaKaRa公司;PCR产物回收试剂盒、质粒回收试剂盒购自北京全式金生物科技有限公司。
1.2 植物总DNA提取
照顾沛雯等[6]的方法提取植物样品的总DNA,于-20℃环境下保存备用。
1.3 PCR扩增、克隆及序列测定
利用Gundersen等[7]设计的通用引物,通过巢式PCR扩增植原体16S rDNA。PCR以植株总DNA为模板,以 R16F2n(5′-GAAACGACTGCTAAGACTG G-3′)和 R16R2(5′-TGACGGGCGGTGTGTACAAA CCCCG-3′)引物对进行扩增。扩增条件为:94℃预处理 5 min;94℃ 40s,55℃ 1 min,72℃ 2 min,共 35 个循环;最后72℃10 min。1.0%琼脂糖凝胶电泳检测PCR产物。PCR扩增出的目标产物纯化回收,克隆入PMD18-T载体中,通过PCR筛选阳性克隆,序列测定委托北京诺赛基因组研究中心有限公司完成。
1.4 序列分析及同源性比较
利用GenBank数据库中的BLAST程序对测序结果进行同源性检索(http://blast.ncbi.nlm.nih.gov/Blast.cgi),通过 DNAMAN5.2.2及 MEGA4.1软件进行同源性比较及系统关系树的构建。
2 结果与分析
2.1 PCR检测结果
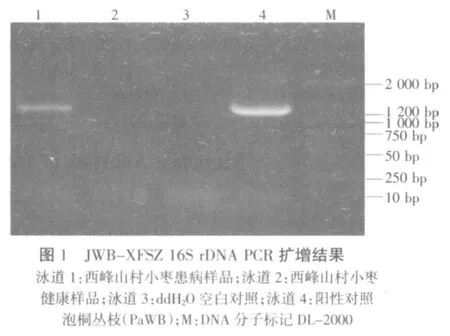
枣疯病感病植株总DNA经直接PCR及巢式PCR扩增后,得到大小约1.2 kb的特异片段,片段大小与引物设计的相符,而健康植株和双蒸水对照均未出现特异扩增带(图1)。将此植原体命名为枣疯病西峰山枣株系(JWB-XFSZ)。
2.2 植原体16S rDNA片段的序列分析
将16S rDNA目的片段克隆后测序,结果表明北京昌平区西峰山村小枣(JWB-XFSZ)的16S rDNA扩增片段含有1248个核苷酸(表1),G+C的含量为45.7%。利用GenBank中的BLAST程序对测序结果进行同源性检索,北京昌平地区枣疯病与16SrⅤ组各植原体的16S rDNA基因序列同源性达98.5%以上,其中与16S rⅤ组的榆树黄化有98.96%、与悬钩子丛枝病有98.64%的同源性。JWB-XFSZ与16SrV-C亚组的枣疯病韩国株系(登录号AB052879)同源性最高,达99.51%,而与其它16Sr其它各组的同源性均在95%以下,表明JWB-XFSZ和枣疯病韩国株系一样应归属于16SrV-C亚组。

表1 JWB-XFSZ 16S rDNA基因的测序结果
2.3 枣疯病植原体系统树构建与分析
将已登陆Genebank的部分植原体16S rDNA基因序列下载下来,与JWB-XFSZ所测定的16S rDNA序列进一步进行同源性分析,利用MEGA4.1软件构建系统发育树(图2)。由系统发育树可以看出,JWB-XFSZ与JWB不同地区的株系;与Elm yellows、Honeylocust yellows、Cherry lethal yellows、Hemp fiber witches-broom、Peach yellows以及 Paper mulberry witches-broom等16SrⅤ组的植原体在同一个进化枝上,说明JWB-XFSZ与16SrⅤ组的亲缘关系最近。
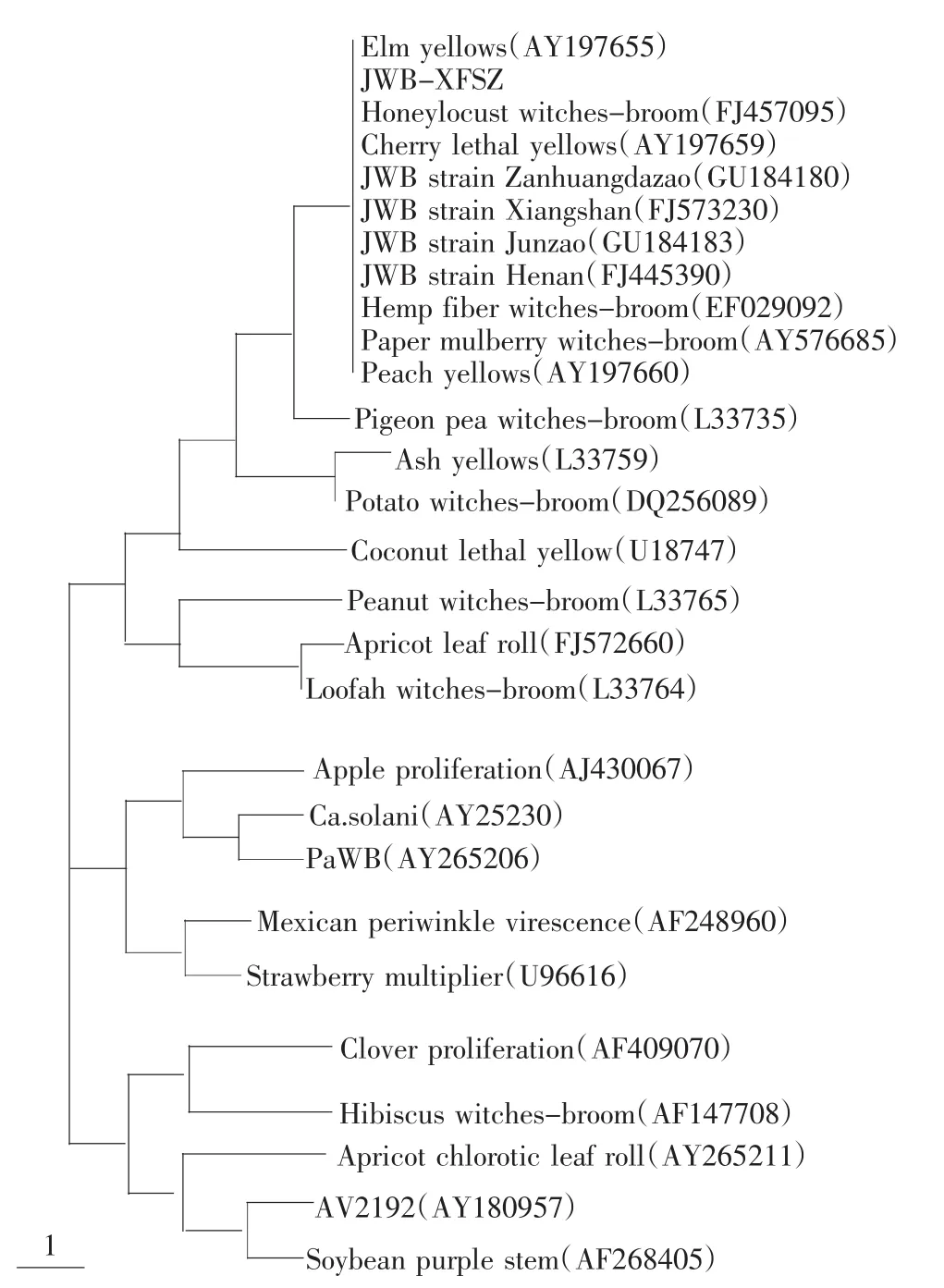
图2 构建28种植原体株系16S rDNA基因核苷酸序列同源关系进化树
3 讨 论
16S rDNA是原核生物染色体基因组中的一种rDNA编码区序列,在进化中高度保守,常作为原核生物系统发育及分类研究的标准方法。本研究运用分子生物学的方法对北京西峰山小枣枣疯病植原体的16S rDNA进行PCR扩增,并对扩增产物进行克隆、序列测定及同源关系分析,确定了该植原体株系的分类地位。
目前,国内外学者对枣疯病植原体病原做了大量研究,以期为枣疯病的防治提供参考依据。Seemüller等[8]认为植原体病害虽发生在世界各地,但不同组及亚组的植原体在分布上存在明显的地区差异,如16S rV-A主要发生在北美国家,16S rV-C主要发生在欧洲,而16S rV-B主要发生在亚洲。从本研究扩增到的枣疯病植原体16S rDNA基因来看,JWB-XFSZ 株系与 JWB-henan、JWB-Zahuangdazao、JWB-xiangshan、JWB-Junzao 以 及JWB-Kor各株系均只有几个碱基的差异,说明枣疯病植原体暂时还未存在株系上的变化。JWB-XFSZ株系与JWB-Kor韩国株系的差异最近,说明北京西峰山枣疯病植原体与韩国报道的植原体是同一种病害。在本研究过程中在病株样品中未扩增到其他植原体,说明未存在复合侵染的现象。
[1]Bai X,Zhang J,Ewing A,et al.Living with genome instability:the adaptation of phytoplas-mas to diverse environments of their insect and plant host [J].Journal of Bacteriology,2006,188:3682-3696.
[2]李 永,田国忠,朴春根,等.我国几种植物上植原体的快速分子鉴别与鉴定的研究[J].植物病理学报,2005,35(4):293-299.
[3]车海彦,罗大全,符瑞益,等.海南长春花黄化病植原体的16S rDNA 序列分析研究[J].植物病理学报,2009,39(2):212-216.
[4]Wei W,Robert E,Davis I-M,et al.Computer-simulated RFLP analysis of 16s rRNA genes:identification of ten new phytoplasma groups[J].Int J Syst Evol Microbiol,2007,57:1855-1867.
[5]侯晓杰,李正楠,冉隆贤,等.越冬病枝水培与PCR结合检测枣疯病植原体的存活动态[J].中国森林病虫,2007,(4):18.
[6]顾沛雯,安凤秋,吴云锋,等.小麦蓝矮病植原体16S rDNA基因片段的比较分析[J].植物病理学报,2005,(5):403-409.
[7]Gundersen D E,Lee IM.Ultrasensitive detection of phytoplasm as by nested-PCR assays using two universal primer pairs[J].Phytopatholo-gia Mediterranea,1996,35:144-151.
[8]Seemüller E,Marcone C,Lauer U,et al.Current status of molecular classification of the phytoplasmas [J].Journal of Plant Pathology,1998,80(1):3-26.
