茉莉酸诱导对枸杞叶生化物质及酶活性的影响
2010-06-12宫玉艳段立清王爱清
宫玉艳, 段立清,2*, 王爱清
(1.内蒙古农业大学林学院,呼和浩特 010019; 2.内蒙古农业大学农学院,呼和浩特 010019)
枸杞(Lycium barbarum Linn.)是名贵的中草药,也是我国西北地区广泛栽培的经济、生态建设树种之一,在内蒙古西部、宁夏、甘肃、新疆等地均有大面积栽培。棉蚜((Aphis gossypii Glover)、枸杞木虱(Paratrioza sinica Yang&Li)、枸杞瘿螨(AceriapallidaKeifer)等害虫常年大发生,人们一直采用化学农药防治枸杞害虫。连续使用化学农药,不仅使害虫产生抗药性,也造成果实污染,使枸杞销量和出口量下降,更为严重的是降低了其作为中草药及保健品的作用[1]。因此,研究外源诱导素诱导枸杞增强抗虫性、减少农药使用量、培育枸杞抗虫品种具有十分重要的意义。
茉莉酸(jasmonic acid,JA)和茉莉酸甲酯(methyljasmonate,MeJA)是植物的受害激素,起信号传递作用,对植物有抑制生长、诱导抗逆、促进衰老等许多生理功能[2-5]。茉莉酸可诱导植物产生生物碱和酚酸类次生物质,它们与蛋白质结合,影响植食者对营养物质的吸收,干扰其神经信号的传递,延缓其发育[6]。植物中某些次生物质和营养物质的质与量的变化对害虫生长发育及代谢有相当大的影响[7]。黄酮是对昆虫有毒的一类次生代谢物质,可影响昆虫的行为和代谢,使之发生忌避和拒食,或破坏昆虫的正常代谢过程,使昆虫中毒甚至死亡[8];单宁酸可干扰昆虫肠道消化和抑制酶活性,阻碍它们的生长发育及繁殖力[9];氮和糖是食物中的两类基本物质,是昆虫消化、活动所需的能量来源,并影响昆虫的生长发育和繁殖[10]。茉莉酸可诱导植物体内某些酶类活性的增强,如多酚氧化酶(PPO)及过氧化物酶(POD),从而影响害虫的生长发育。PPO是在植物体内广泛存在的一种抗营养酶类,能够催化多酚化合物次生代谢,降低植物的营养价值,阻止昆虫的取食,抑制昆虫的生长发育[11-14]。POD是植物保护酶系的重要保护酶之一,在植物体内与乙烯的生物合成及吲哚乙酸的氧化作用有关[15]。目前,枸杞对外源茉莉酸的生理反应尚未有任何报道,本文就外源茉莉酸对枸杞叶黄酮、单宁酸、蛋白质、可溶性糖、PPO和POD的影响进行了研究,探讨枸杞受到外界刺激后的生理反应,为今后深入研究枸杞诱导抗性、培育枸杞新品种提供科学依据。
1 材料与方法
1.1 材料
供试枸杞:为宁夏枸杞(Lycium barbarum Linn.),将枸杞分批播种于盆内,根据土壤湿度,定期浇水,待枸杞长至20cm时进行试验。
仪器与试剂:智能气候培养箱(哈尔滨东拓科技有限责任公司)、电热恒温培养箱(湖北省黄石市医疗器械厂)、索氏提取器(姜堰市腰庄玻璃仪器制造厂)、高速冷冻离心机(Hettich Zentrifugen GmbH&Co.KG)、卧式低温保存箱(青岛海尔特种电器有限公司)、HH-S型恒温水浴锅(武汉金宝华科技有限公司)、TU-1810紫外可见分光光度计(北京普析通用仪器有限责任公司)、移液枪(法国GILON公司);茉莉酸(jasmonicacid,简称 JA)购于 Sigma公司、丙酮、磷酸缓冲液(pH6.0、pH6.5、pH7.0)、F-D试 剂、Na2CO3、甲醇、蒽酮试剂、NaNO2、AL(NO3)3、考马斯亮蓝G-250、邻苯二酚、三氯乙酸 、H2O2、愈创木酚 。
1.2 枸杞苗木处理
将茉莉酸先溶于丙酮,然后加蒸馏水配制成所需浓度。PPO、POD的测定设置 0.0001、0.001、0.01、0.1、1.0mmol/L5 种浓度 ;黄酮、单宁酸 、蛋白质、可溶性糖的测定设置0.001、0.01、0.1mmol/L3种浓度,以丙酮+蒸馏水作为对照。用手持式喷雾器向健康枸杞苗木均匀喷施茉莉酸溶液,液滴下流为止,然后放置在气候培养箱培养。分1、3、5、7d4个时间段进行测定。
1.3 测定方法
黄酮含量的测定采用索氏回流法[16]。
单宁酸含量的测定采用磷钼酸-磷钨酸比色法(F-D法)[17]。
蛋白质含量的测定采用考马斯亮蓝G-250法[18]。可溶性糖含量的测定采用蒽酮比色法[18]。
PPO活性的测定参照朱广廉的方法进行提取和测定[19]。
POD活性的测定采用愈创木酚法[20]。
1.4 数据处理
采用SPSS13.0统计软件进行Duncan分析。
2 结果与分析
2.1 茉莉酸处理对枸杞叶黄酮含量的影响
3种浓度的JA处理使枸杞叶黄酮含量均较对照显著增加(p<0.05),增加的程度与JA的浓度不成正相关关系(图1)。黄酮含量在处理后第5天增幅最大,到第7天时,增幅降低。浓度为0.1mmol/LJA处理使枸杞叶黄酮含量在处理后的第5天较对照增加了1.3倍,由处理前的3.998mg/g增加到5.351mg/g。JA 浓度相同,时间不同,枸杞黄酮含量差异显著,而对照的差异不显著,黄酮含量趋于平稳。
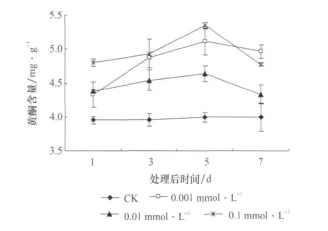
图1 茉莉酸处理后枸杞叶黄酮含量随时间的变化
2.2 茉莉酸处理对枸杞叶单宁酸含量的影响
3种浓度的 JA处理使枸杞叶单宁酸含量均较对照组显著增加(p<0.05),增加的程度与JA的浓度不成正相关关系(图2)。单宁酸含量增加的持续性表明,处理后第5天增幅最大,到第7天时,增幅降低。浓度为0.1mmol/LJA处理使枸杞叶单宁酸含量在处理后的第5天较对照增加了近1.4倍,由处理前的 0.655mg/g增加到0.892mg/g。JA浓度相同,时间不同,枸杞单宁酸含量差异显著,而对照的差异不显著,单宁酸含量趋于平稳。
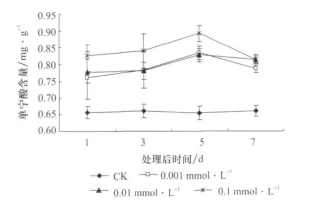
图2 茉莉酸处理后枸杞叶单宁酸含量随时间的变化
2.3 茉莉酸处理对枸杞叶蛋白质含量的影响
3种浓度的JA处理使枸杞叶蛋白质含量均较对照显著降低(p<0.05),降低的程度与JA的浓度不成正相关关系(图3)。蛋白质含量降低的持续性表明,处理后第7天降幅最大,浓度为0.001mmol/LJA处理使枸杞叶蛋白质含量由处理前的72.226mg/g降低到47.047mg/g。JA浓度相同,时间不同,枸杞蛋白质含量差异显著,而对照的差异不显著,蛋白质含量趋于平稳。
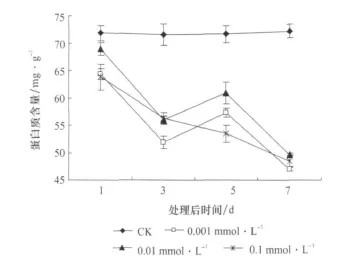
图3 茉莉酸处理后枸杞叶蛋白质含量随时间的变化
2.4 茉莉酸处理对枸杞叶可溶性糖含量的影响
3种浓度的JA处理使枸杞叶可溶性糖含量均较对照显著降低(p<0.05),降低的程度与JA的浓度不成正相关关系(图4)。可溶性糖含量降低的持续性表明,处理后第 1天降幅最大,浓度为0.1mmol/LJA处理使枸杞叶可溶性糖含量由处理前的63.604mg/g降低到54.962mg/g。JA浓度相同,时间不同,枸杞可溶性糖含量处理组及对照均有差异。

图4 茉莉酸处理后枸杞叶可溶性糖含量随时间的变化
2.5 茉莉酸处理对枸杞叶PPO活性的影响
5种浓度的JA处理使枸杞叶PPO活性均较对照显著增加(p<0.05),增加的程度与JA的浓度不成比例(表1)。PPO活性增加的持续性表明,浓度为0.1mmol/LJA处理的枸杞叶在处理后的第3天增幅最大。JA浓度相同,时间不同,枸杞叶PPO活性除0.001mmol/L外差异显著,而对照的差异不显著,PPO活性趋于平稳。

表1 茉莉酸处理后枸杞叶PPO活性随时间的变化1)
2.6 茉莉酸处理对枸杞叶POD活性的影响
5种浓度的JA处理使枸杞叶POD活性均较对照显著增加(p<0.05),增加的程度与 JA的浓度不成比例(表2)。POD活性增加的持续性表明,浓度为0.01mmol/LJA处理的枸杞叶在处理后的第7天增幅最大。JA浓度相同,时间不同,枸杞叶POD活性处理组及对照均有差异。

表2 茉莉酸处理后枸杞叶POD活性随时间的变化1)
3 讨论
茉莉酸处理后枸杞叶黄酮、单宁酸含量显著增加,蛋白质、可溶性糖含量显著降低、PPO和POD活性增加,体现了枸杞叶对茉莉酸的诱导反应。枸杞叶受到茉莉酸刺激后黄酮、单宁酸含量的升高,蛋白质、可溶性糖含量的降低可能是一种应激反应[21],与其他植物一样,通过次生物质合成量的增加,营养物质合成量的降低以达到抗虫目的[22]。枸杞叶黄酮、单宁酸、蛋白质、可溶性糖含量增加或降低与茉莉酸浓度不成线性相关,可能与植物的应激反应存在抗性阈值有关[23]。
应用外源茉莉酸处理番茄(Lycopersiconesculentum),番茄叶PPO、POD的活性增强,取食该番茄的烟草天蛾幼虫生长受到抑制[24],减少了4类昆虫(鳞翅目幼虫、甲虫、蚜虫和蓟马)的危害数量[25]。茉莉酸处理的枸杞叶PPO、POD的活性增强,但能否减少枸杞的害虫种类与危害程度有待于进一步深入研究。茉莉酸甲酯处理茶树诱导茶树叶PPO和蛋白酶抑制素活性增加[26];MeJA处理烟草(N.attenuata)和杨树(Populustrichocarpa),两者的PPO活性分别增加了22.9倍和5.4倍[27]。茉莉酸处理引起枸杞叶PPO和POD活性的增加,但增加程度较杨树、烟草小,并未成倍增加。茉莉酸刺激是否可引起枸杞蛋白酶抑制素、植物凝聚素等其他影响昆虫生长发育物质的变化,还有待于进一步深入研究。
[1]段立清,邹晓林,冯淑军,等.枸杞上的主要害虫、天敌及其综合管理[J].内蒙古农业大学学报,2002,23(4):51-54.
[2]Sembdner G,Parthier B.The biochemistry and the physiological and molecular action of jasmonates[J].Annu Rev Plant Physiol Plant M ol Biol,1993,44:569-589.
[3]Creelman R A.Jasmonic acid distribution and action in plants:Regulation during development and response to biotic and abiotic stress[J].Proc Natl Acad Sci USA,1995,92:4114-4119.
[4]Creelman R A,Mullet J E.Biosynthesis and action of jasmonates in plants[J].Annu Rev Plant Mol Biol,1997,48:355-381.
[5]王妮妍,蒋德安.茉莉酸及其甲酯与植物诱导抗病性[J].植物生理学通讯,2002,38(3):279-284.
[6]Wasternack C,Parthier B.Jasmonate-signalle plant gene expression[J].Trends Plant Sci,1997,2:302-309.
[7]Masttsten W J,Sciber J M.Nutritional ecology of insect folivores of woody plants:nitrogen,water,fiter and mineral considerations[M]∥Slansky Jr F,Rodriguez J G,eds.Nutrition-al ecology of insects,mites and spiders.New York:Wiley Press,1987:105-146.
[8]武予清,郭予元.棉花单宁—黄酮类化合物对棉铃虫的抗性潜力[J].生态学报,2001,21(2):286-289.
[9]王海波,周纪纶.蚕豆对蚕豆蚜虫刺吸胁迫的生理防御策略[J].生态学报,1993,8(3):195-200.
[10]黄炜东.两色绿刺蛾为害对毛竹营养物质的影响[J].竹子研究汇刊,2006,25(4):24-27.
[11]Stout M J,Workman K V,Bostock R M.Stimulation and attenuation of induced resistance by elicitors and inhibitors of chemical induction in tomato(Lycopersicon esculentum)foliage[J].Entomologia Experimentalis et Applicata,1998,86:267-279.
[12]Felton G W,Donato K,Broadway R M.Impact of oxidized plant phenolics on the nutritional quality of dietary protein to a noctuid S podoptera exigua[J].Journal of Insect Physiology,1992,38:277-285.
[13]Keinanen M,Oldham N J,Baldwin I T.Rapid HPLC screening of jasmonate-induced increase in tobacco alkaoids,phenolics and diterpene glycosides in N icotiana attenuata[J].Journal of Ag ricultural Food Chemistry,2001,49:3553-3558.
[14]Tscharntke T,Thiessen S,Dolch R,et al.Herbivory,induced resistance,andinterplant signal transfer in Alnusgltinosa[J].Biochemical Systematics and Ecology,2001,29:1025-1047.
[15]Shuban K R,A tal K R.Changes in enzyme activity and isoperoxidases in haploid tobacco callus during organogenesis[J].Plant Science Letter,1982,24:67-77.
[16]汤章城.现代植物生理学试验指南[M].北京:科学出版社,1999.[17]张治安,张美善.植物生理学试验指导[M].北京:农业科学技术出版社,2004.
[18]陈毓荃.生物化学试验方法和技术[M].北京:科学出版社,2002.
[19]朱广廉.植物生理学实验[M].北京:北京大学出版社,1990.
[20]华东师范大学生物系.植物生理学实验指导[M].人民教育出版社,1980:143-144.
[21]张思禄.竹篦舟蛾为害对毛竹次生物质及营养物质的影响[J].竹子研究汇刊,2008,27(2):21-24.
[22]张潮巨.毛竹受害后化学物质含量的变化及对刚竹毒蛾种群参数的影响[J].华东昆虫学报,2006,15(4):281-283.
[23]陈巨莲,倪汉祥,孙京瑞.主要次生物质对麦蚜的抗性阈值及交互作用[J].植物保护学报,2002,29(1):7-12.
[24]Cipollini D F J R,Redman A M.Age-dependent effects of jasmonic acid treatment and wind exposure on foliar oxidase activity and insect resistance in tomato[J].Journal of Chemical Ecology,1999,25(2):271-281.
[25]Thaler J S,Stout M J,Karban R,et al.Jasmonate-mediated induced plant resistance affects a community of herbivores[J].Ecol Entomol,2001,26:312-324.
[26]桂连友,陈宗懋,刘树生.外源茉莉酸甲酯处理茶树对茶尺蠖幼虫生长的影响[J].中国农业科学,2005,38(2):302-307.
[27]Constabel C P,Ryan C A.A survey of wound-and methyl jasmonate-induced leaf polyphenol oxidase in crop plants[J].Phy tochemistry,1998,47(4):507-511.
[27]ConstabelCP,RyanCA.Asurveyofwound-andmethyljasmonate-inducedleafpolyphenoloxidaseincropplants[J].Phytochemistry,1998,47(4):507-511.
