虫酰肼对甜菜夜蛾六类酶活性的影响
2010-06-12王贻莲司升云汪钟信宫成玉周利琳
王贻莲, 司升云, 汪钟信, 宫成玉, 周利琳
(1.武汉市蔬菜科学研究所,武汉 430065; 2.山东省科学院中日友好生物技术研究中心,济南 250014;3.华中农业大学植物科学技术学院,武汉 430070; 4.烟台出入境检验检疫局,烟台 264000)
虫酰肼是一种昆虫生长调节剂,能模拟蜕皮激素20-羟基蜕皮酮与目标害虫蜕皮激素受体复合物相互作用,实现蜕皮激素的作用方式。到目前为止,国内外对酰肼类杀虫剂引起甜菜夜蛾抗药性的研究还较少,仅限于作用机理、抗性监测等方面,在抗性遗传及抗性机制等方面报道较少[1-3]。为研究甜菜夜蛾对虫酰肼的抗药性机制,本文采用活体测定法对甜菜夜蛾武汉敏感(CK)种群和自然(N)种群进行了表皮酚氧化酶(PPO)、乙酰胆碱酯酶(TCE)、谷胱甘肽-S转移酶(GST)、超氧化物歧化酶(SOD)和过氧化氢酶(CAT)酶活力测定,采用PAGE凝胶电泳对CK种群、汰选种群和N种群的酯酶同工酶(EI)进行比较,以期有助于阐明甜菜夜蛾对虫酰肼的抗药性机制,为甜菜夜蛾抗性治理提供理论依据。
1 材料与方法
1.1 供试昆虫
甜菜夜蛾武汉自然(N)种群(抗性比25.29)采于武汉市蔬菜科学研究所实验田;武汉敏感(CK)种群由武汉市蔬菜科学研究所无毒饲养多年获得;处理种群用按一定湿重比混匀虫酰肼(处理浓度为敏感种群的致死中量0.14 mg/kg)的带毒饲料饲喂的甜菜夜蛾CK种群;汰选种群由武汉敏感种群开始筛选,共筛选了4~6代。
1.2 供试药剂及试剂
20%虫酰肼悬浮剂,美国陶氏益农公司;牛血清白蛋白,Roche 分装 ;固蓝 RR 盐 、α-乙酸萘酯 、β-乙酸萘酯、丙烯酰胺、甘氨酸、β-巯基乙醇、过硫酸铵、溴酚蓝、三羟甲基氨基甲烷、甲叉双丙烯酰胺,Sigma公司;谷胱甘肽S-转移酶、超氧化物歧化酶、过氧化氢酶及乙酰胆碱酯酶测试盒,南京建成生物工程研究所;其他常规试剂均为国产。
1.3 虫酰肼对甜菜夜蛾六类酶活性测定
酶样品制备、测定方法均严格按测试盒说明书操作;数据用DPS软件处理系统处理;酶活抑制率计算公式:
抑制率=(对照组酶活力-处理组酶活力)/对照组酶活力×100%;
酯酶同工酶电泳:参照张龙翔方法,采用聚丙烯酰胺(PAGE)凝胶垂直板电泳。
2 结果与分析
2.1 虫酰肼对甜菜夜蛾 PPO、TCE、GST、SOD和CAT的影响
由表1可知,用混有0.14 mg/kg虫酰肼的带毒饲料饲喂CK种群的甜菜夜蛾,在饲喂后24、48 h和72 h测定酶活力,与不用药处理的CK种群相比,PPO比活力分别降低79.12%、10.54%和56.23%;SOD比活力分别上升71.59%、63.16%和79.55%;GST比活力在处理后24 h酶活力上升41.43%;TCE和CAT比活力差异不显著。N种群与不用药处理的CK种群比,PPO比活力降低38.47%;GST比活力升高34.20%;TCE、CAT和SOD比活力差异不显著。
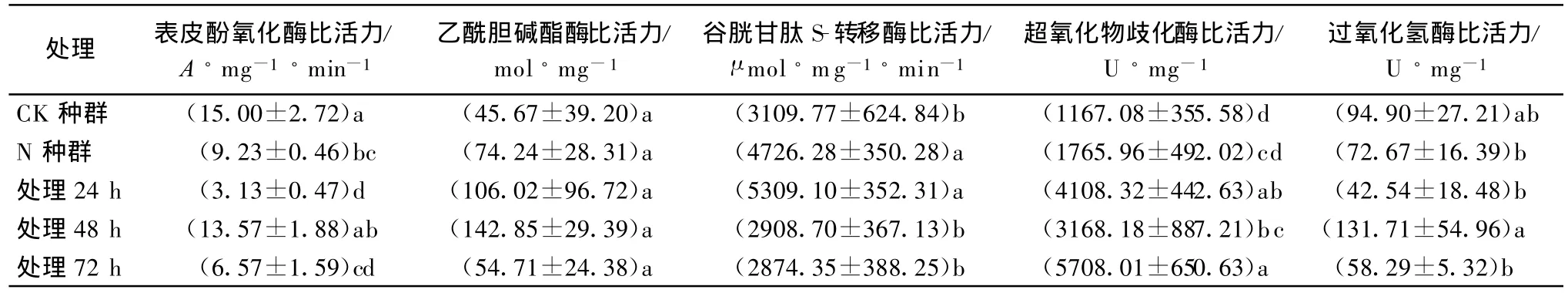
表1 处理种群、CK种群和N种群的酶活力比较1)
2.2 酯酶同工酶(EI)电泳
CK种群、N种群和汰选种群的酯酶同工酶电泳结果见图1。
由图1可知,CK种群与N种群、汰选种群的酯酶同工酶差异极显著,谱带的数量、位置、宽度及清晰度均有差异。CK种群酯酶同工酶共15条带,第2、12、13条带细且清晰度差,第5和6、7和8条带近,不易分开;N种群酯酶同工酶共10条带,第1、9条带细但清晰,第6、7、8、10条带清晰度差,第1条带的位置与其他种群的不同;汰选种群酯酶同工酶共10条带,第1、7、8条带清晰度差,第3和4、5和6条带很近,不易分开;汰选种群中的六汰后第2代(六汰F2)酯酶同工酶共14条带,即在CK种群第11条带的位置缺失,第2条带模糊不清,第12条带清晰度差。
3 讨论
研究表明甜菜夜蛾对虫酰肼的抗性可能与PPO、EI、GST、SOD 有一定关系,与 CAT 关系不大,与TCE无关。
多酚氧化酶广泛存在于昆虫的体壁和血液中,在昆虫体壁形成的硬化和黑化过程中起重要作用[4-5]。用混有虫酰肼的带毒饲料处理试虫后,处理种群的酶活力均低于CK种群,且在处理后24 h和72 h与CK种群存在显著差异,表明虫酰肼很可能可以抑制表皮酚氧化酶酶活力。抗性比为25.29的N种群酶的比活力为9.23A/(mg◦min),低于CK种群,且差异显著,表明N种群的抗性增长与表皮酚氧化酶有关。Retnakaran报道,供试昆虫在蜕皮中,虫酰肼抑制了多巴脱羧酶(DDC)的活性,使羽化激素的释放受到了抑制[6]。在昆虫表皮鞣化过程中,多酚氧化酶具有单酚酶活性和二酚酶活性,能将酪氨酸羟化,产生邻位二羟基苯丙氨酸(L-多巴),然后将多巴氧化成多巴醌,最终生成黑色素,使昆虫初生的柔软、色淡的新表皮得以黑化和硬化[5,7-8]。本研究认为,在这一过程中,多巴脱羧酶的活性起着至关重要的作用,加之Retnakaran的报道,这就很好地解释了虫酰肼为什么可以抑制表皮酚氧化酶的活性。
处理种群在饲喂带毒饲料后24、48 h和72 h,表皮酚氧化酶酶活抑制率分别为79.12%、10.54%和56.23%。酶活抑制率产生这种变化的原因可能是药剂和昆虫自身调节综合作用的结果,当杀虫剂通过昆虫取食进入虫体后,就会最大限度地发挥毒杀作用,抑制表皮酚氧化酶的活性;然而昆虫可以通过自身的调节功能来抵抗这种作用,提高表皮酚氧化酶的活性,在两者的相互作用达到平衡之前,酶活力的变化就会反复,直到相互作用达到平衡时,酶活力才会出现一个相对稳定值,后续结果有待进一步测试。另外,表皮酚氧化酶的释放高峰在昆虫蜕皮时,甜菜夜蛾幼虫在28℃下平均48 h蜕皮一次,所以这也可能是在试虫处理后48 h其酶活力与对照无明显差异的原因之一。
本研究用混有虫酰肼的带毒饲料饲喂试虫后,GST的比活力在饲喂后24 h与CK种群相比差异显著,与N种群相比差异不显著,而在饲喂后48 h和72 h GST的比活力与N种群存在显著差异,与CK种群相比差异不显著。说明虫酰肼很可能可以促使甜菜夜蛾的GST酶活力提高,增强其解毒代谢的能力,导致甜菜夜蛾对虫酰肼抵抗力增强。兰亦全等报道甜菜夜蛾福州自然种群对虫酰肼的抗性与谷胱甘肽S-转移酶有一定关系[9]与本研究相符。
本研究用混有虫酰肼的带毒饲料饲喂试虫后,SOD酶的比活力在饲喂后24、48 h和 72 h均明显高于CK种群,且差异显著。表明虫酰肼很可能可促使甜菜夜蛾SOD酶活力提高,增强其保护功能,提高甜菜夜蛾的抵抗能力。用混有虫酰肼的带毒饲料饲喂试虫后,CAT酶的比活力在饲喂后48 h,与24 h和72 h相比差异显著,处理种群与CK种群相比差异均不显著。说明甜菜夜蛾对虫酰肼的抗性与CAT的关系可能不大。
本研究用混有虫酰肼的带毒饲料处理试虫后,乙酰胆碱酯酶的比活力与CK种群、N种群无差异存在。说明甜菜夜蛾对虫酰肼的抗药性与乙酰胆碱酯酶可能无关。酯酶同工酶电泳显示,N种群、汰选种群(相对抗性种群)与CK种群均存在显著差异,说明甜菜夜蛾对虫酰肼的抗药性与酯酶同工酶可能有很大的相关性。
以上各种酶在用虫酰肼处理试虫后不同时间酶活力的变化差异,同样也可以用药剂和昆虫自身调节综合作用的理论来解释。
[1]杜育哲,刘安西.非甾醇蜕皮激素类杀虫剂的研究进展[J].农药,2002,41(6):7-11.
[2]冯坚.双酰肼类昆虫生长调节剂研究开发进展[J].现代农药,2004,3(2):4-8.
[3]黄琳瑞.甜菜夜蛾对虫酰肼抗药性选育和抗药性机理初探[D].北京:中国农业大学,2005.
[4]罗万春,高兴祥,于天丛.栎皮酮对甜菜夜蛾酚氧化酶的抑制作用[J].昆虫学报,2005,48(1):36-41.
[5]薛超彬,王勤,柯莉娜,等.铜铁试剂对菜青虫多酚氧化酶的抑制作用[J].昆虫学报,2005,48(2):290-294.
[6]Retnakaran A,Hiruma K,Palli S R,et al.M olecular analy sis of the mode of action of RH-5992,a Lepidopteran-specific,non-steroidalecdysteroidag onist[J].Insect Biochem Molec Biol,1995,29:1015-1027.
[7]许可,唐明,沈璐辉,等.昆虫蜕皮行为的生理生化和分子生物学研究进展[J].昆虫学报,2001,44(2):244-250.
[8]高兴祥,罗万春,谢桂英,等.甜菜夜蛾多酚氧化酶的特性及其对曲酸等抑制剂的反应[J].中国农业科学,2004,37(5):687-691.
[9]兰亦全,赵士煕.甜菜夜蛾抗药性监测及机理[J].福建农林大学学报(自然科学版),2004,33(1):26-29.
