温度对绿僵菌LA菌株孢子萌发、酶活力及毒力的影响
2010-06-12雷仲仁王兆勇姚君明
岳 梅, 雷仲仁, 孟 涛, 王兆勇, 姚君明
(中国农业科学院植物保护研究所,植物病虫害生物学国家重点实验室,北京 100193)
绿僵菌是虫生真菌主要类群之一,寄主范围广,能寄生8目30科约200种昆虫、螨类及线虫[1],是当今虫生真菌研究的主要对象之一。影响绿僵菌生长和生理代谢的因素较多,其中温度是最重要的因素之一[2],绿僵菌入侵昆虫的每一步都与温度紧密相关。
东亚飞蝗是我国历史上的重要害虫,其为害植物以禾本科、莎草科为主,至今仍严重威胁我国的粮食生产[3]。近年来,全球性气候变化为蝗灾的发生创造了有利条件,加重了蝗灾的发生。绿僵菌防治蝗虫具有很好的效果,但是近些年的研究表明,蝗虫具有较强的体温调节行为,当体温升高至一定程度便能有效地降低绿僵菌的致病力,因此绿僵菌耐受高温的能力便成为影响病原与寄主互作关系的重要因素。作者即对绿僵菌LA菌株在不同温度下的萌发率、胞外蛋白酶和几丁质酶活力以及对东亚飞蝗的毒力进行了研究,并与东亚飞蝗室内接受辐射热可保持的适宜体温结合,测定了不同高温处理下绿僵菌孢子萌发情况,以期明确温度对绿僵菌与蝗虫互作的影响。
1 材料与方法
1.1 供试昆虫与菌株
本试验所用东亚飞蝗系实验室多年续代饲养,于网室内每天用新鲜玉米叶片和麦麸饲喂的健康群体。
绿僵菌LA菌株,最早分离自天津大港的东亚飞蝗,本实验室续代保存。
1.2 温度对LA菌株孢子萌发的影响
采用微室萌发法[4]测定孢子萌发率,取载玻片置于保湿的培养皿中,分别滴入孢子萌发液(1%蛋白胨、1%葡萄糖和0.05%吐温80)和孢子液各一滴混匀,封口膜密封培养皿,制成萌发微室。萌发率为选定视野内萌发孢子数与总孢子数的比值,每次计数5个视野求平均值。
1.2.1 不同温度下孢子萌发速率
将上述培养皿分别置于 20、25、30、35 ℃和40℃人工气候箱(RXZ智能型,宁波江南仪器厂)内培养,设置温度的误差为1℃,箱内湿度保持在40%~50%。4h后开始计数萌发率,每隔4h计数1次,记录24h。重复3次求平均数。
1.2.2 高温下孢子萌发速率测定
根据1.2.1部分的结果,设定35~40℃之间每隔1℃为一温度梯度,即分别将待测孢子置于36、37、38℃和39℃,培养8h后开始测定萌发率,每隔8h计数1次,记录48h,测定孢子可萌发的最高温度。重复3次求平均数。
1.2.3 不同高温处理对孢子萌发速率的影响
用无菌水配制孢子悬浮液,倒入2mL离心管中,分别置于42、45 ℃和 48 ℃水浴中,处理 3、6h和9h,取出后用微室培养法置于30℃下培养,4h后计数萌发率,每隔4h计数1次,记录 24h。重复3次求平均数。
1.3 温度对LA菌株酶活力的影响
1.3.1 培养基
SDA 培养基 :葡萄糖40g/L,蛋白胨10g/L,酵母浸膏 10g/L。蛋白酶诱导培养基:1%明胶,K2HPO4,NaCl和MgSO4◦7H2O 各 0.3g/L。各组分溶入pH7.0的0.02mol/L磷酸缓冲液。几丁质诱导培养基:蛋白胨0.5%,KH2PO4,KCl,MgSO4◦7H2O各0.05%,ZnSO4◦7H2O0.1%,脱矿几丁质1%,pH6.0。
胶体几丁质的制备:称取几丁质25g,85%磷酸250mL,30℃反应1.5d,加蒸馏水待胶体沉淀后,反复水洗至中性,用 0.02mol/L磷酸缓冲液(pH6.0)调整终浓度至2%。
1.3.2 不同培养时间胞外蛋白酶活力的测定
将少量LA菌粉加入胞外蛋白酶诱导培养基中,30℃,130r/min旋转式摇床培养,分别提取第3天、第4天、第5天、第6天、第7天的酶液,将2%酪蛋白1mL加入1mL酶液中,40℃反应10min,加2mL0.4mol/L三氯乙酸溶液中止反应,室温放置待沉淀完全。用Folin-酚试剂检测,以每分钟催化分解1μg酪蛋白的酶量为一个活性单位(U/mL)。重复3次求平均数。
1.3.3 温度对胞外蛋白酶活力的影响
1.3.4 不同培养时间几丁质酶活力的测定
将少量菌粉加入到几丁质诱导培养基中30℃,130r/min旋转式摇床培养,分别提取第3天、第4天、第5天、第6天、第 7天的酶液,在1mL胶体几丁质中,分别加入酶液0.5mL,50℃反应1h,用DNS法测定产生的还原糖,以每分钟产生相当于1μL的NAG所需的酶量定义为一个酶活力单位(U/mL)。重复3次求平均数。
1.3.5 温度对几丁质酶活力的影响
取1.3.4中培养第5天的酶液,在1mL胶体几丁质中 ,加入酶液 0.5mL,20 、25、30、35、40、45、50、55℃下反应1h,用DNS法测定产生的还原糖。以每分钟产生相当于1μg的NAG所需的酶量定义为一个酶活力单位(U/mL)。重复3次求平均数。
1.4 温度对LA菌株毒力的影响
配制孢子浓度为2.6×107个/mL,每头蝗虫接种0.5μL,设定人工气候箱温度分别为 20、25、30、35、40℃5个温度,选用5龄东亚飞蝗若虫,每个处理3个重复并设对照,每个重复选用20头试虫,每日调查蝗虫死亡情况并将死虫保湿培养,记录15d,计算 LT50和LT90。
1.5 数据处理分析
应用DPS软件进行数据处理分析并计算LT50、LT90。
2 结果与分析
2.1 不同温度下孢子萌发速率
采用微室萌发法测定了不同温度下LA菌株分生孢子萌发情况,结果如图1,LA孢子在30℃下4h即有10.2%孢子萌发,8h萌发率达到72.3%,35℃下4h也有5.1%孢子萌发,总体而言,前12h孢子萌发速率依次为30℃>35℃>25℃>20℃>40℃,LA孢子在20~35℃均可萌发,24h内均能达到90%以上的萌发率,而40℃下24h内孢子无萌发现象。

图1 温度对LA分生孢子萌发的影响
2.2 高温下孢子萌发速率
LA分生孢子在36~39℃下培养48h内孢子萌发情况如图2所示。温度每升高1℃,孢子萌发率均有较大程度降低,36℃下48h孢子萌发率达到80.2%,而39℃下仅为10.1%,此温度范围属LA萌发的不适合温度,高于39℃的环境温度下分生孢子基本不能正常萌发。
互联网与通信技术的发展打破了传统学习的时空约束,改变了学生的学习行为,使学习更具有灵活性和主动性,网络学习的即时性也最大限度的帮助成人学生克服工学矛盾, 但网络学习改变成人学习行为的同时也产生了一些消极的影响。在未来成人教育体系中,学生不仅仅是传统课堂学习的“补充型”学习,而且是终身学习的践行者,而网络学习给成人学生的终身学习提供了条件。成人教育的研究者和工作者,应更客观地看待互联网在网络学习中的作用和学生具体学习行为的嬗变,努力扬长避短,使成人学生在网络学习中朝着更有益的方向发展。

图2 高温下LA分生孢子的萌发情况
2.3 不同高温处理对孢子萌发速率的影响
不同时间高温处理对孢子萌发率的影响见表1,由表可以看出,随着处理温度的升高和处理时间的延长,孢子萌发率均有不同程度的降低,48℃下对孢子萌发影响最大,处理9h后30℃下微室培养24h萌发率仅为10%。42℃和45℃高温处理转至30℃下培养24h,孢子萌发率均在85%以上,试验2.1中未经高温处理30℃下8h萌发率达70%,而经高温处理后8h萌发率最高仅为11.7%。整体而言,在本试验所处理的时间范围内,48℃下分生孢子受高温影响,转至最适培养温度下也无法正常萌发;42℃和45℃处理一定时间,延缓了孢子的萌发,但不能最终影响孢子萌发率。
2.4 LA菌株胞外蛋白酶活力测定
2.4.1 不同培养时间胞外蛋白酶活力测定
培养第3、4、5、6、7天所得胞外蛋白酶活力曲线如图3所示,随着培养天数的增加,酶活力逐渐升高,至第6天达到最高,活力值为31.37U/mL。
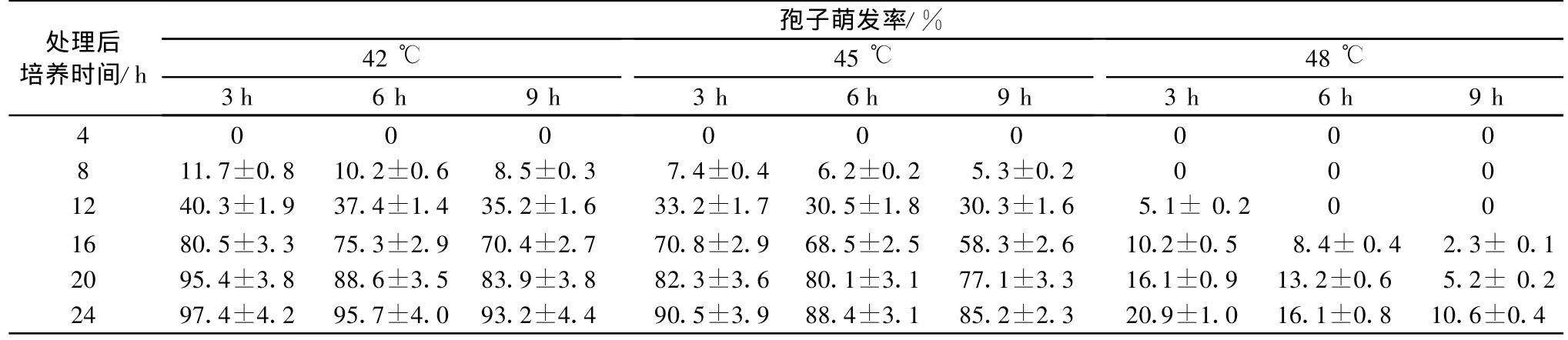
表1 不同高温处理时间对LA孢子萌发率的影响1)

图3 不同培养时间LA胞外蛋白酶活力
2.4.2 温度对胞外蛋白酶活力的影响
取培养第6天的蛋白酶液,在不同温度下反应所得的酶活力结果见图4,由图可以看出,37℃左右蛋白酶活力最高,20℃下蛋白酶活力最低,活力值为20.62U/mL,酶活力是一个先增加后降低的过程。
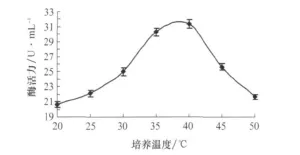
图4 不同温度下LA胞外蛋白酶活力
2.5 LA菌株几丁质酶活力测定
2.5.1 不同培养时间几丁质酶活力的测定
培养第 3、4、5、6、7天所得几丁质酶活力曲线如图5所示,酶活力至第 5天达到最高,活力值为34.62U/mL,之后继续培养呈下降趋势。

图5 不同培养时间LA几丁质酶活力
2.5.2 温度对几丁质酶活力的影响
取培养第5天的几丁质酶液,在不同温度下反应所得的酶活力结果见图6,由图可以看出,几丁质酶活力在50℃下最高,活力值为 34.62U/mL,20℃下最低,活力值为23.53U/mL,几丁质酶活力是一个先增加后降低的过程。

图6 不同温度下LA几丁质酶活力
2.6 温度对LA菌株毒力的影响
不同温度下LA对东亚飞蝗5龄若虫的毒力见表2,由此表可以看出30℃下LT50和 LT90最低,20℃和40℃下致死时间较长,25℃和35℃下致死时间居中,30℃为 LA对东亚飞蝗致病力最强的温度。

表2 不同温度下LA对东亚飞蝗致死时间1)
试虫一般从第3天开始大量死亡,不同温度下试虫的累计死亡率见图7,不同温度下LA的毒力差异可从死亡曲线的延伸角度和高度直观看出,30℃下第8天试虫死亡率已达100%,而40℃下至15d仍有部分试虫未死亡。
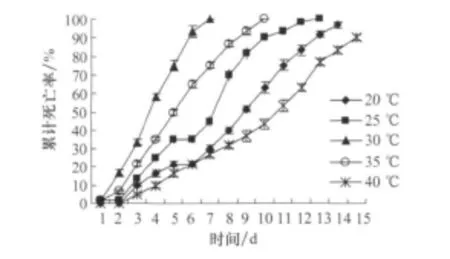
图7 不同温度下接种LA东亚飞蝗的累计死亡率
3 讨论
在适宜的温度范围内,绿僵菌LA孢子萌发速率快,致病力强,相反,如果环境温度过高或过低,超出了其适合生长侵染的温度,致病力便会大大降低。
本研究结果显示,30℃是LA孢子萌发的最适温度,在此温度下菌株LA对蝗虫的致死速率最快,38℃下培养48h,孢子的萌发率只有10%左右,40℃下培养孢子的萌发率趋近于0,说明该菌株孢子萌发受温度影响较大。蝗虫适宜发育的温度在25~40℃之间,最适宜温度范围为28~34℃[5],在有体温调节或太阳辐射条件下,虫体温度可以达到42℃,此温度限制了LA孢子的萌发,这也是在田间应用绿僵菌治蝗后如遇晴天高温,其防效不好的主要原因。
绿僵菌的致死温度(10min内杀死孢子的速度)介于49~60℃之间[6],菌丝体生长的上限温度约在37~40℃[7-8]。经高温处理的孢子,移至适合温度下培养,萌发速率和萌发率均有所降低,说明孢子萌发及存活情况与所达到的高温程度和该温度处理的时间有关。本试验最高处理温度为48℃,该温度下导致绝大部分孢子失去萌发能力,恢复至30℃培养萌发率24h仅为20%左右。
冯明光[9]研究认为胞外蛋白酶活性可作为球孢白僵菌大量菌株初筛的参考性毒力指标,但应谨慎使用,不能完全取代常规的毒力测定。林海萍等[10]对15个球孢白僵菌菌株进行了酶活力和毒力测定,认为这些菌株的蛋白酶、几丁质酶、脂肪酶活性与对松墨天牛毒力间呈极显著正相关。本试验选用LA菌株蛋白酶和几丁质酶活力最高的培养时间,进行了不同温度下的酶活力测定,结合对应温度下对东亚飞蝗毒力的测定,结果表明,在菌株的极限高温和极限低温时菌株的胞外蛋白酶产量明显减少,其相应的LT50也较大;而在菌株的最适生长温度时,其相应的LT50值较小,说明胞外蛋白酶产生水平与毒力之间存在一定的协同性,即产酶水平越高,菌株对东亚飞蝗的毒力就越大,因此可以认为绿僵菌的胞外蛋白酶合成能力是其毒力的决定因素之一。Kucera[11]和St.Leger等[12]也指出绿僵菌的蛋白酶活性与其毒力密切相关。但几丁质酶活力表达最高的温度与LA毒力最强的温度不一致,因此几丁质酶活力不能作为毒力表达的指标。
[1]蒲蛰龙.昆虫病理学[M].广州:广东科技出版社,1994:360-368.
[2]毕国昌,郭秀珍,臧穆.在纯培养条件下温度对外生菌菌根真菌生长的影响[J].林业科学研究,1989,2(3):247-252.
[3]黄冬如.绿僵菌对东亚飞蝗致病机理的初步研究[D].长沙:湖南农业大学,2004.
[4]张立钦.松墨天牛致病菌优良菌株筛选及其应用的研究[D].江苏:南京林业大学,2000.
[5]郭郛,陈永林,卢宝廉.中国飞蝗生物学[M].济南:山东科学技术出版社,1989:420.
[6] Liu H,Skinner M,Brownbridge M,et al.Characterization of Beauveria bassiana and Metarhizium anisopliae isolates for management of tarnished plant bug,Lygus lineolaris(Hemiptera:Miridae)[J].Journal of Invertebrate Pathology,2003,82:139-147.
[7] Fargues J,Goettel M S,Smits N,et al.Variability in susceptibility to simulated sunlight of conidia among isolates of entomopathogenic hy phomycetes[J].Mycopathologia,1996,135:171-181.
[8] Hallsworth J E,Magan N.Water and temperature relations of growth of the entomogenous fungi Beauveriabassiana,Metarhizium anisopliae and Paecilomy ces farinosus[J].Journal of Invertebrate Pathology,1999,74:261-266.
[9]冯明光.胞外蛋白酶和脂酶活性作为球孢白僵菌毒力指标的可靠性分析[J].微生物学报,1998,38(6):461-467.
[10]林海萍,魏锦瑜,毛胜凤,等.球孢白僵菌蛋白酶、几丁质酶、脂肪酶活性与其毒力相关性[J].中国生物防治,2008,24(3):290-292.
[11]Kucera M.Protease from the fungus Metarhizium anisopliae toxic for Galleria melonella larvae[J].Journal of Invertebrate Pathology,1980,35:304-310.
[12]St.Leger R J,Durrands P K,Charnley A K,et al.Role of extracellular chymoelastase in the virulence of Metarhizium anisopliae for Manduca sexta[J].Journal of Invertebrate Pathology,1988,52:285-293.
