多杀菌素低剂量处理西花蓟马对药剂敏感性的影响
2010-06-12龚佑辉吴青君张友军徐宝云
龚佑辉, 吴青君, 张友军, 徐宝云
(中国农业科学院蔬菜花卉研究所,北京 100081)
西花蓟马是一种世界性园艺害虫,多杀菌素是为数不多的防治效果非常好的药剂之一[1-3]。但已有研究报道西花蓟马对多杀菌素产生了抗药性[4-6]。杀虫剂用于田间后,随着个体接触药量的差异以及时间的推移,对部分个体还存在着亚致死效应,包括生物学的改变,生态学行为改变,生殖力的变化,生理生化改变,抗药性的发展等[7]。而且有学者认为害虫再猖獗的产生在相当程度上与低剂量农药刺激害虫增殖有关[8-9]。目前,多杀菌素已广泛应用于我国鳞翅目害虫和蓟马的防治中,在多杀菌素长期的低剂量选择压力下,西花蓟马是否对多杀菌素出现敏感性下降,以及是否会对其他杀虫剂的敏感性产生影响均有待研究。
本研究通过用多杀菌素低剂量继代处理西花蓟马,连续选育47代后测定了处理种群对其他5种不同类型杀虫剂的敏感性差异,拟探讨在连续低剂量选择压下西花蓟马对多杀菌素的抗药性风险,以及是否影响对其他杀虫剂的敏感性,研究结果可为田间西花蓟马综合防治和多杀菌素的合理使用提供指导。
1 材料与方法
1.1 供试昆虫及药剂
西花蓟马敏感种群(SS):2003年从中国农业科学院蔬菜花卉研究所温室内的甜瓜(CucumismeloL.)上采集[10],采用豆荚法在实验室饲养至今。饲养条件为:25℃±1℃,RH 65%,光周期L∥D=16h∥8h。饲养期间未接触任何杀虫剂。多杀菌素低剂量选育西花蓟马种群(Sub):用多杀菌素LC25浓度对敏感种群连续处理(每次处理的浓度为上一次生物测定的LC25值)。
药剂:多杀菌素(spinosad)(2.5%悬浮剂,美国陶氏益农公司)、阿维菌素原药 (abamectin)(97%,河北威远化工有限公司)、甲氨基阿维菌素苯甲酸盐原药 (emamectin benzoate)(57.6%,北京华戎化工有限公司)、吡虫啉原药(imidacloprid)(95.3%,江苏常隆化工有限公司)、高效氯氰菊酯原药 (beta-cypermethrin)(95.4%,南京红太阳股份有限公司)、吡丙醚原药(pyriproxyfen)(96%,南运施壮化工有限公司)。
1.2 方法
1.2.1 生物测定方法
参照 Alfredo & Anthony[11]的 TIBS法,做 适当的修改建立了叶管药膜法。将每个浓度药液分别注满1.5mL离心管4只,每只为1重复,盖紧盖子之后平放4h。然后倒掉药液,将离心管置于室温下晾干,待用。用长约10cm左右的细针针尖经酒精灯加热,在晾干的离心管(1.5mL)管底烫直径约2~3mm的孔。将无虫新鲜的嫩甘蓝叶洗干净,晾干后用打孔器打成直径为1.5cm的小圆片。分别在每个浓度药液中浸10s。浸过的叶碟置实验台上晾干,待用。在吸虫之前,用小镊子将叶碟夹入相应药液浓度的离心管中,每管1片,放置平整。真空泵的接口用纱布封好,套在以上处理好的离心管管口,打开真空泵,调节气流大小,把离心管管底的烫孔对准蓟马,使虫顺气流吸入离心管。先对照后处理,每管吸2龄若虫15~25头,然后盖好管盖,用封口膜封好烫孔。25℃环境下放置2d或3d后调查结果。数据用POLO软件处理,计算LC25、LC50值及其95%置信区间等。高效氯氰菊酯和吡丙醚原药用丙酮溶解,其他药剂原药都先用少量丙酮溶解后再用含1‰曲拉通(Triton X-100)的蒸馏水稀释。高效氯氰菊酯的试虫用刚孵化3d后的成虫,吡丙醚的试虫用1龄若虫,观察时间为4d。其他药剂都用2龄若虫作为试虫。
1.2.2 低剂量多杀菌素对西花蓟马的处理方法
自2007年1月开始从西花蓟马敏感种群中分出一部分西花蓟马单独饲养,用多杀菌素生物测定获得的LC25浓度浸泡新鲜干净的豆荚2h,自然晾干后喂食西花蓟马成虫,整个成虫期均用药剂处理后的豆荚饲喂,若虫用未用药处理的豆荚饲喂。每隔几代进行生物测定,每次处理的浓度为上一次LC25的计算值。到2009年5月为止,已用LC25剂量处理47代。
2 结果与分析
2.1 低剂量处理种群对多杀菌素的敏感性变化
2007年1月开始采用多杀菌素的LC25剂量对西花蓟马进行处理,到2009年5月处理第47代。
从表1可以看出,室内敏感种群在完全没有杀虫剂的环境中饲养,且没有外来种群混杂下,随着时间的推移,西花蓟马敏感种群对多杀菌素的敏感性不断上升,LC50到达0.020 0mg/L时基本稳定。而多杀菌素低剂量处理的种群在11代后对多杀菌素的敏感性下降1.5倍,处理36代后已下降5.19倍,处理47代后,Sub种群的LC50是敏感种群的6.72倍,达到了低水平抗性。
2.2 低剂量处理种群对其他杀虫剂的敏感性变化
对西花蓟马用多杀菌素LC25剂量处理41代后,测定了其对其他5种杀虫剂的敏感性。表2显示,当Sub种群对多杀菌素的敏感性下降为SS种群的5.19倍时,Sub种群对阿维菌素、甲氨基阿维菌素苯甲酸盐和吡虫啉敏感性也下降,分别为敏感种群的2.68、2.92倍和1.89倍,且差异显著。对高效氯氰菊酯和吡丙醚的敏感性无明显种群差异。
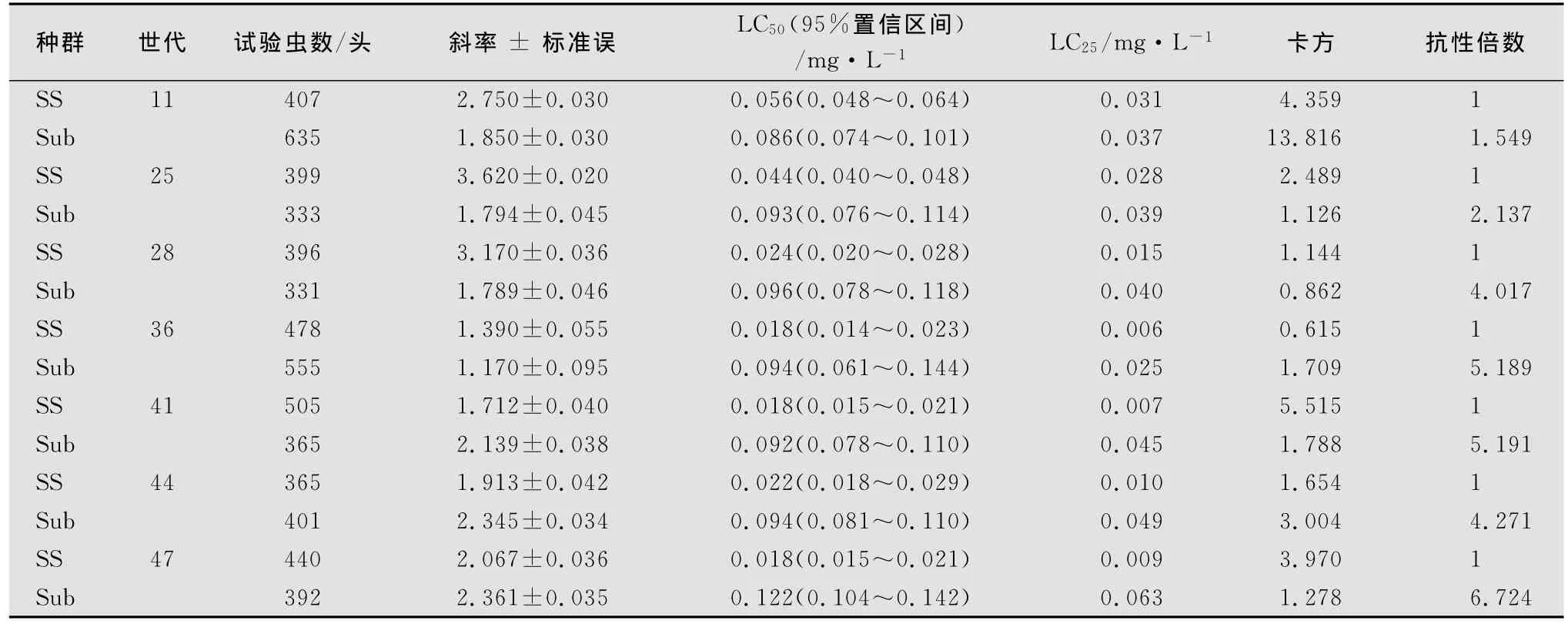
表1 西花蓟马敏感种群和多杀菌素低剂量处理种群对多杀菌素敏感性变化

表2 不同杀虫剂对西花蓟马敏感种群和多杀菌素低剂量处理种群毒力测定
3 结论与讨论
对于那些繁殖速率快、发生世代多的害虫如蚜虫、螨类、小菜蛾、蓟马等,如果长期处于药剂低剂量的选择压力下,对药剂敏感性低的个体存活和繁殖机会较大,对整个种群来说有利于抗药性能力的积累和发展。高宗仁等[12]用低剂量溴氰菊酯、氧乐果、杀虫脒和三氯杀螨醇等4种药剂对朱砂叶螨(Tetranychuscinnabarinus)连续选择3代后发现,其对这4种药剂的敏感度均有不同程度的下降。在室内,Young等[13]采用LD50剂量的多杀菌素连续处理烟蚜6代后,抗性倍数仅为1.68倍,但是连续处理14代后,抗性倍数高达1 068倍。在本试验中,随着低剂量多杀菌素处理代数的增加,处理种群对多杀菌素敏感性下降的趋势在增大,并逐渐产生低水平抗性,说明在长期低剂量的选择压力下,西花蓟马对多杀菌素有产生抗药性的风险。
许多研究认为,多杀菌素与其他药剂之间无交互抗性。在本研究中对多杀菌素有低水平抗性的种群对阿维菌素和甲氨基阿维菌素苯甲酸盐的敏感性下降了2倍多,说明用多杀菌素低剂量处理西花蓟马后,会影响其对其他药剂的敏感性,有可能导致田间西花蓟马的再猖獗危害。交互抗性多存在于具有相同作用机制的杀虫剂之间,不同类型杀虫剂也会由于相同的解毒代谢机制产生交互抗性。有报道显示,西花蓟马对一些传统类杀虫剂的抗性机制是多功能氧化酶介导的代谢抗性[14]。低水平抗性下,西花蓟马对多杀菌素的抗性可能主要由于解毒酶活性的升高而致。在本试验中,阿维菌素、甲氨基阿维菌素苯甲酸盐对多杀菌素低剂量处理种群的毒力降低,推测可能与西花蓟马对其具有相同的解毒机制有关。
吡丙醚是昆虫生长调节剂,与多杀菌素的作用机制完全不同,虽然吡丙醚对西花蓟马的活性不高,但也可以考虑在抗性治理中作为轮换使用的药剂。
[1]Elzen G W,Elzen P J,King E G.Laboratory toxicity of insecticide residues toOriusinsidiosus,Geocorispunctipes,HippodamiaconvergensandChrysoperlacarnea[J].Southwestern Entomol,1998,23:335-34.
[2]Ludwing S,Oetting R.Effect of spinosad onOriusinsidiosus(Hemiptera:Anthocoridae)when used forFrankliniellaoccidentalis(Thysanoptera:Thripidae)control on greenhouse pot chrysanthemums[J].Fla Entomol,2001,84:311-313.
[3]Eger J E,Stavisky J,Funderburk J E.Comparative toxicity of spinosad toFrankliniellaspp.(Thysanoptera:Thripidae),with notes on a bioassay technique[J].Fla Entomol,1998,81:547-551.
[4]Herron G A,James T M.Monitoring insecticide resistance in AustralianFrankliniellaoccidentalisPergande (Thysanoptera:Thripidae)detects fipronil and spinosad resistance[J].Austral J Entomol,2005,44:299-303.
[5]Loughner R L,Warnock D F,Cloyd R A.Resistance of greenhouse,laboratory,and native populations of western flower thrips to spinosad[J].Hortscience,2005,40:146-149.
[6]Bielza P,Quinto V,Contreras J,et al.Resistance to spinosad in the western flower thrips,Frankliniellaoccidentalis(Pergande),in greenhouses of south-eastern Spain[J].Pest Manag Sci,2007,63:682-687.
[7]Desneux N,Decourtye A,Delpuech J M.The sublethal effects of pesticides on beneficial arthropods[J].Annu Rev Entomol,2007,52:81-106.
[8]Morse J G,Zareh N.Pesticide-induced hormoligosis ofCitrus thrips(Thysanoptera:Thripidae)fecundity[J].J Econ Entomol,1991,84:1169-1174.
[9]Nandihalli B S,Pati1BV,Hugar P.Influence of synthetic pyrethroid usage on aphid resurgence in cotton[J].Karnataka J of Agric Sci,1992,5(3):234-237.
[10]Zhang Z J,Wu Q J,Li X F,et al.Life history of western flower thrips,Frankliniellaoccidentalis(Thysan.,Thripae)on five different vegetable leaves[J].J Appl Entomol,2007,131(5):347-354.
[11]Alfredo R,Anthony MS.Development of a bioassay systeMfor monitoring susceptibility inThripstabaci[J].Pest Manag Sci,2003,59:553-558.
[12]高宗仁,李巧丝,刘孝纯.杀虫剂对朱砂叶螨某些生物学特性的影响[J].植物保护学报,1991,18(3):283-287.
[13]Young H P,Bailey W D,Roe R M.Spinosad selection of a laboratory strain of the tobacco budworm,Heliothisvirescens(Lepidoptera:Noctuidae),and characterization of resistance[J].Crop Protection,2003,22:265-273.
[14]Espinosa P J,Contreras J,Quinto V,et al.Metabolic mechanisms of insecticide resistance in the western flower thrips,Frankliniellaoccidentalis(Pergande)[J].Pest Manag Sci,2005,61:1009-1015.
