声音在耳内的信号转导及其分子生物学机制(5)
2010-06-05汤浩石丽娟于利曹宇
汤浩 石丽娟 于利 曹宇
·继续教育园地·
声音在耳内的信号转导及其分子生物学机制(5)
汤浩1石丽娟1于利1曹宇1
近年来对豚鼠的实验研究发现,与外淋巴接触的毛细胞的基底膜上存在两种可被Ca2+激活的钾通道,两者的开放均依赖于细胞内Ca2+浓度的升高。纤毛的弯曲使毛细胞顶部的机械门控离子通道开放,导致内淋巴中高浓度的K+流向细胞内,使毛细胞发生去极化反应。此时位于侧膜上的电压依赖型钙通道开放,导致Ca2+内流。毛细胞内的Ca2+浓度升高引起毛细胞底部的递质向突触间隙释放,同时又激活毛细胞基底侧膜上的Ca2+激活钾通道,造成K+外流,使毛细胞的膜电位接近于K+平衡电位,为毛细胞顶部的机械门控通道提供大量的电化学驱动力,有助于毛细胞的机械-电换能作用。
3 来自毛细胞的突触传递在低振幅感受器电位时被触发
毛细胞除是感觉感受器外,也是突触前终端。每个细胞的基底外侧膜包括了许多突触前活性区,在这里化学神经递质被释放出来。每个活性区域都有四个显著的形态特征(图10)。在临近释放位置的胞浆中有一个突触前致密小体(dense body),它是一个球形的或卵圆形的、直径大约400 nm的纤维状嗜锇性结构。尽管它的生化成分还不清楚,但致密小体同视觉感受器细胞的突触带很相似,并且可能是对发现于神经肌肉接头和中枢神经系统突触中的更小的突触前致密点进行了加工。突触前致密小体由直径大约35 nm的核心清楚的突触小泡围绕着,有时这些小泡会被很细的细丝黏附在致密小体上。在致密小体和突触前浆膜之间有一个显著的致密点,它通常由许多短小的绒毛状的物质构成。在质膜内,成排的膜内粒子同突触前致密点对齐。这些粒子包括与递质释放有关的Ca2+通道和参与电共鸣的K+通道。很少有的关于哺乳动物的毛细胞的传入突触的生理学研究显示,哺乳动物突触同其它脊椎动物是一致的。因此,从模型中得出的结论也可以应用于哺乳动物的突触。而对非哺乳类的脊椎动物的研究表明,同其它的突触一样(如神经突触),毛细胞的递质释放是由突触前去极化引起的,并且需要胞外介质中Ca2+的存在。突触后记录显示毛细胞突触递质的释放是量子式的;这些突触的统计学性质同神经肌肉接头相似,传入神经递质的特性是有争议的。
毛细胞传入突触有许多的特征,而这些特征就成了毛细胞具有专门发射信号的能力的基础。首先,许多毛细胞在静息时释放递质。在突触传递过程中,所释放递质的数量和由其所致的传入神经纤维活性分别依赖于毛细胞从静息电位的去极化或是复极化。同这一现象一致,毛细胞Ca2+通道在静息时被激活,形成稳定的Ca2+渗漏,引起未受刺激细胞的递质释放。其次,同视觉感受器一样,它们必须能够释放神经递质对只有100μV或更大的阈值感受器电位做出回应,从而使突触前Ca2+通道在静息电位时被激活,最终毛细胞能够对高频刺激产生反应,尤其是那些哺乳动物的耳蜗,必须能够高速会聚突触小泡,并且速度要大到足以确实发出信号。尽管人们还不知道突触前致密小体的作用,但是作为主细胞中的结构突出物,致密小体同突触前小泡释放器的关系十分紧密,表明,它与最小限度刺激的神经递质异常快速释放有关。
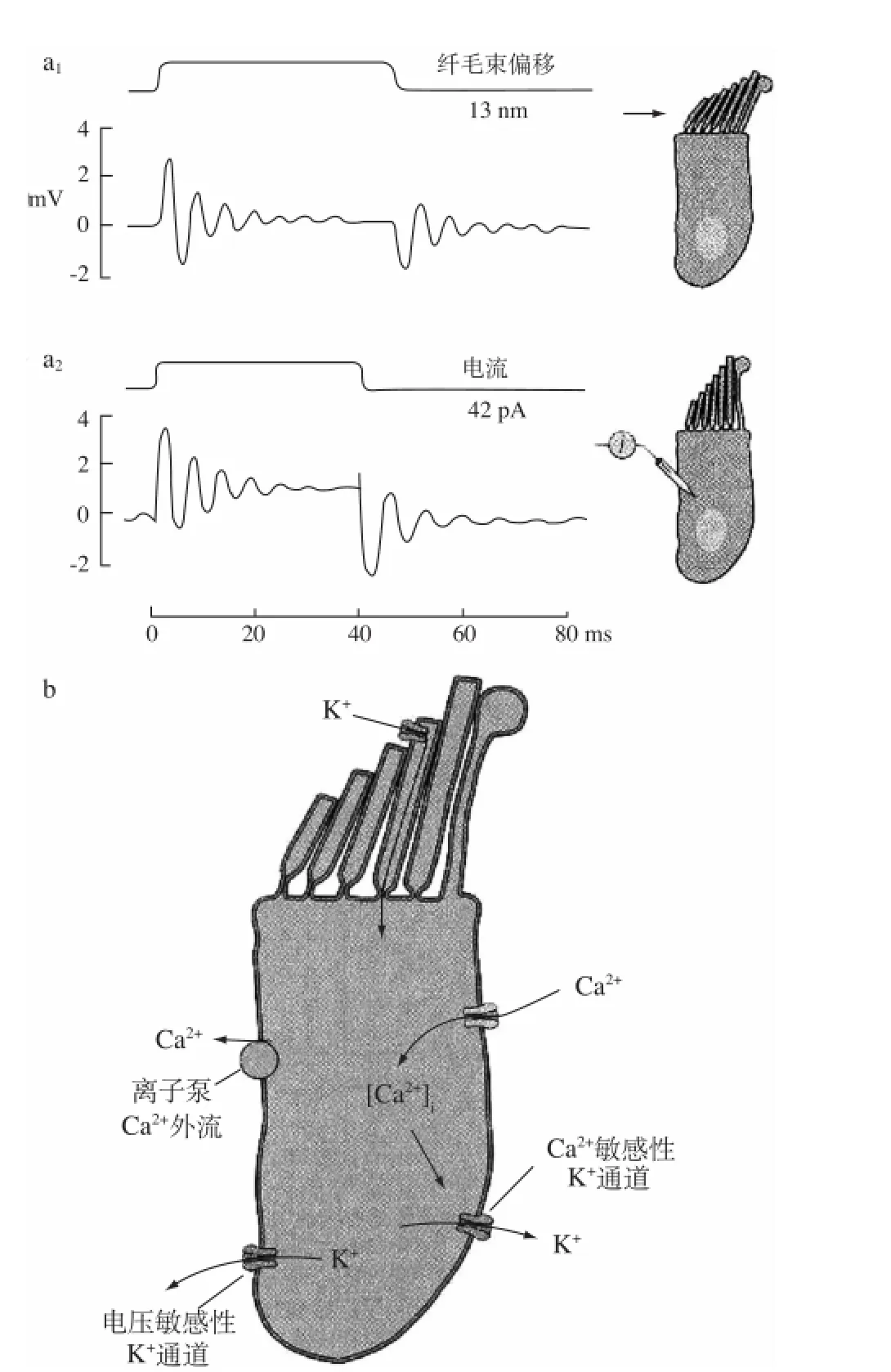
图9 海龟内耳毛细胞的共振电反应(引自Crawford and Fettiplace,1985)[12]
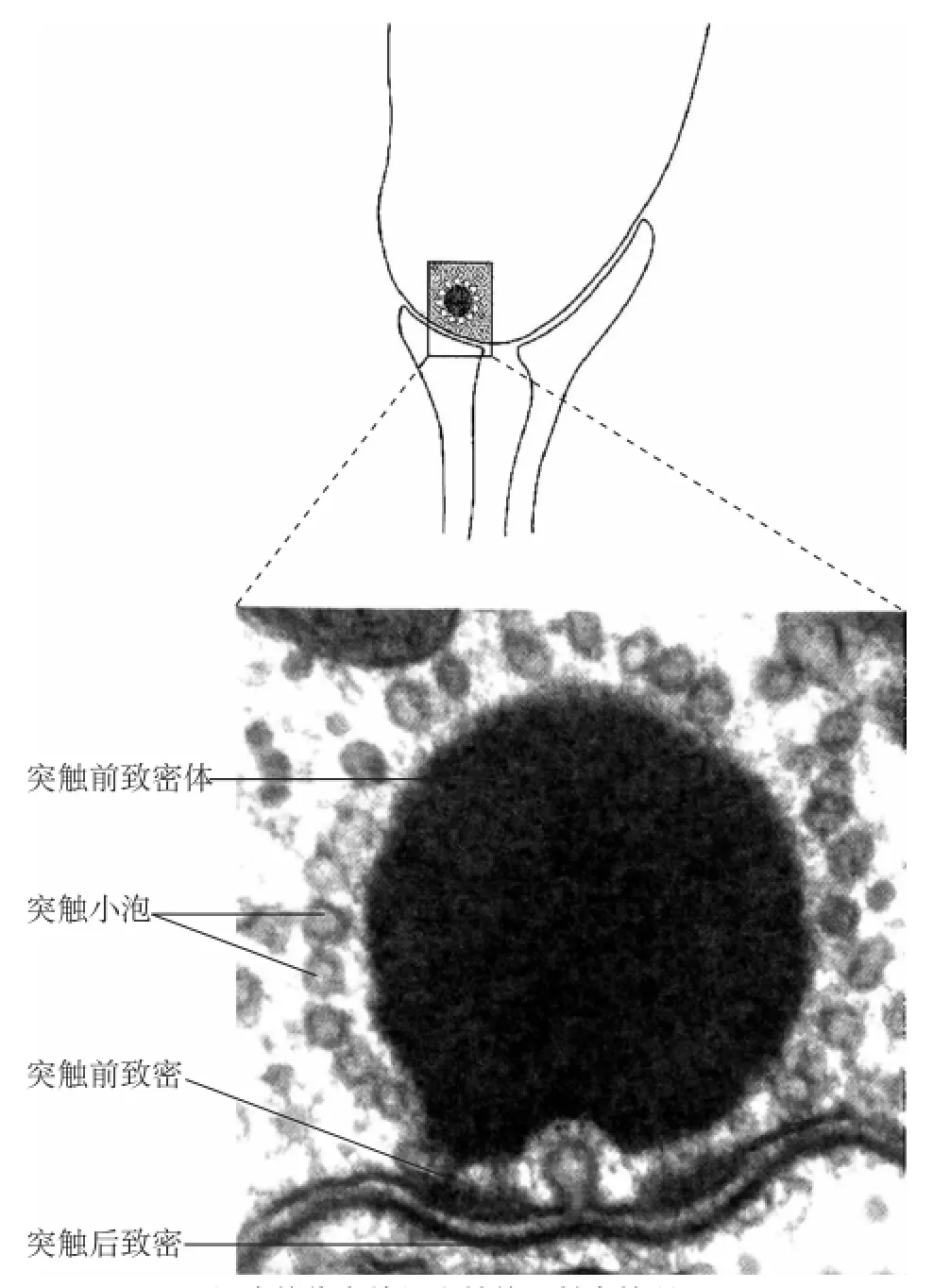
图10 毛细胞的传出神经突触的透射电镜所见
大部分毛细胞从脑干部的神经元接收传出突触输入信号,传出终端是一种典型的外周突触,传出轴在毛细胞表面相对较大的小结处终结[13]。突触前胞浆含有大量的清晰的直径约为50 nm的突触小泡和少量较大的、核心清晰的小泡。最重要的递质是乙酰胆碱(ACh)。但是,降钙素基因相关肽(CGRP)也出现在传出终端并且可能同ACh一起释放。在每个传出终端的下面,毛细胞的突触后胞浆形成一个滑面内质网池,作为对传出刺激的反应,这一结构可能同Ca2+的释放和重吸收有关。
在不同的毛细胞中,传出刺激有不同的效果。在前庭器,它能降低或增高与靶毛细胞相连的传入纤维的神经活性。在使用电共鸣来进行频率调谐的毛细胞中传出神经支配的作用是最易理解的。一方面当传出神经受到刺激时,传出递质使得靶细胞超极化。更重要的是,递质诱导膜电导增强打乱了毛细胞膜上的已调谐的共鸣回路。这样,频率选择性的清晰度和电放大的增进都减少了。研究表明,在外界噪音刺激或其他精神因素影响下,动物脑干电位的记录波形受到干扰,而在人的实验中,表现为无法听清人们的语言交流或无法享受美妙的音乐,均为传出神经受到刺激而对毛细胞的调谐功能产生影响的结果。
目前临床上使用的耳声发射仪的原理可能就是基于外毛细胞能够加强耳蜗敏感性和频率选择性的缘故,而且认为外毛细胞有放大作用,故可作为放大的能量来源。实验用电刺激一个单离的外毛细胞所表现的活性是:去极化时胞体缩短,超极化时伸长,运动的能量来源于电刺激,而不是ATP等能源介质。因此认为可能是当外毛细胞把纤毛上的机械刺激转化为感受器电位的同时发生了放大作用,与细胞体变化相应的细胞膜电位的变化可能会放大基底膜的运动。
1 于利,汤浩.哺乳动物耳蜗外毛细胞的马达蛋白:Prestin[J].生理科学进展,2008,39:1.
2 Brownell WE,Bader CR,Bertrand D,et al.Evoked mechanical responses of isolated cochlear outer hair cells[J].Science,1985,227:194.
3 Kandel ER,Schartz JH,Jessel TM.Principles of neural science[M].4th ed.New York:McGraw-Hill Medical,2000.591~612,614~624.
4 范少光,汤浩,主编.人体生理学[M].第三版.北京:北京大学出版社,2005.416~420.
5 Miller JM,Towe AL.Audition:Structural and acoustical properties[M].In:Ruch T,Patton HD,eds.Physiology and Biophysics,Vol 1.The Brain and Neural Function.20th ed.Philadelphia:Saunder,1979.339~375.
6 Pickles JO.An Introduction to the physiology of hearing[M].2nd ed.New York:Academic,1988.203~235.
7 Howard J,Hudspeth AJ.'Compliance of the hair bundle Associated with gating of mechanoelectrical transduction channels in the Bullfrog's saccular hair cell'[J].Neuron,1988,1:189.
8 Hudspeth AJ.How the ear's works work[J].Nature,1989,341:397.
9 Jaramillo F,Hudspeth AJ.Localization of the hair cell's transduction channels at the hair bundle's top by iontophoretic application of a channel blocker[J].Neuron,1991,7:409.
10 Hudspeth AJ.Hair-bundle mechanics and a model for mechanoelectrical transduction by hair cells[J].Society of General Physiologists Series,1992,47:357.
11 Hudspeth AJ,Gillespie PG.Pulling springs to tune transduction:Adaptation by hair cells[J].Neuron,1994,12:1.
12 Crawford AC,Fettiplace R.The mechanical properties of ciliary bundles of turtle cochlear hair cells[J].J Physiol,1985,364:359.
13 Jacobs RA,Hudspeth AJ.Ultrastructural correlates of mechanoelectrical transduction in hair cells of the bullfrog's internal ear[J].Cold Spring Harbor Symposia on Quantitative Biology,1990,55:547.
(全文完)
(2009-02-11收稿)
(本文编辑 李翠娥)
10.3969/j.issn.1006-7299.2010.04.032
R339.16
E
1006-7299(2010)04-0411-02
1 中国医科大学基础医学院生理教研室(沈阳 110001)
