珍稀濒危植物天台鹅耳枥营养器官的解剖学研究
2010-06-04陈模舜柯世省杨勇宇颜倩倩
陈模舜,柯世省,杨勇宇,颜倩倩
(台州学院 生命科学学院,浙江 临海 317000)
天台鹅耳枥(Carpinus tientaiensis)属桦木科鹅耳枥属,只分布在浙江省天台县华顶山西茅棚,仅存13株母株,被列为国家二级重点保护野生植物[1~2]。目前生境的严重破碎化直接影响到小种群植物的生存,天台鹅耳枥在研究植物地理、植物区系和生物多样性等方面均有较高的科研价值[3~4]。目前国内外有关天台鹅耳枥保护生物学的研究甚少,特别是对其解剖学方面的研究尚未见报道。本文对天台鹅耳枥的芽、根、茎和叶进行了解剖学特征的观察和研究,希望为研究天台鹅耳枥在其生长环境中的适应情况及其系统演化和区系分类提供解剖学资料,在此基础上提出科学的优先保护序列并制定合理的保护策略,以期能更好地保护天台鹅耳枥。
1 材料与方法
1.1 材料
所用材料为野生天台鹅耳枥,采自浙江天台县华顶山(29º 15′ N、121 º 06′ E),海拔900 m处。2008年5月采集天台鹅耳枥根、茎和芽,根、茎切成1 cm左右的小段,根、茎和芽均用FAA固定48 h以上,根、茎固定后需用软化剂(95%乙醇:甘油 = 1:1)软化1 ~ 2个月才可使用。2008年7月采集3个生境(林窗、林缘和林下)的天台鹅耳枥叶,林窗日最大PPFD约1 120 μmol·m-2·s-1,林缘日最大PPFD约480 μmol·m-2·s-1,林下日最大PPFD约45 μmol·m-2·s-1,采用顶芽以下第4片成熟的叶片,采样后剪取沿中脉两侧约1.5 cm×1.5 cm的小块,用FAA固定48 h以上。以上材料采用Rotary Microtome HM 315切片机石蜡切片,切片厚8 µm,酒精脱水,二甲苯透明,固绿番红对染,中性树胶封片[5]。叶表皮采用铬酸、硝酸离析法(Geffrey)(10%铬酸:10%硝酸= 1:1)得到。
1.2 方法
对本实验(解离法和石蜡切片法)做成的装片于Leica DFC 320显微镜下观察并拍照,照片留在计算机硬盘上存档。利用Leica QWin V3计算机显微图像分析系统进行图像分析,解剖结构测量取每试验材料装片5张,每观察取10个视野,数值为平均值±标准误,显微图像经Adobe Photoshop 10.0图像处理系统制版。
2 结果与分析
2.1 天台鹅耳枥营养器官的解剖结构
2.1.1 天台鹅耳枥芽的解剖结构 天台鹅耳枥芽外面有鳞片包被,芽由顶端分生组织、叶原基、腋芽原基和幼叶构成(图1-1)。顶端分生组织位于芽的顶端,具原套—原体结构[6],原套具1 ~ 3层细胞,只进行垂周分裂,使表面连续生长;原体原始细胞位于原套下方,它的衍生细胞形成肋状分生组织和周围分生组织,将来分化出茎皮层和茎的维管组织。在顶端分生组织下面的一些突起为叶原基,叶原基是叶的原始体,叶原基愈向下愈明显,较下的已发育成幼叶,这些幼叶包被着芽。腋芽原基是芽的每个叶原基的腋部的小型突起物,它们是腋芽的原始体,腋芽展开后长成侧枝。
2.1.2 天台鹅耳枥根的解剖结构 天台鹅耳枥主根发达,属直根系;根的横切面近似圆形,主要由周皮和次生维管组织组成,属根的次生结构(图1-2)。根的周皮厚,外层表皮多枯死脱落,周皮具有5 ~ 7层木栓层,细胞扁平,长方形,1层木栓形成层和2 ~ 3层栓内层细胞。皮层位于周皮之内,由3 ~ 5层薄壁细胞组成,薄壁细胞长×宽 =(15.30±2.37)μm×(5.67±0.34)μm。
次生维管组织位于皮层以内,它由次生韧皮部和次生木质部组成,次生木质部居内,次生韧皮部在外,相对排列。次生韧皮部所占比例较小,韧皮纤维较发达;次生木质部极为发达,导管数量多,导管平均切向径(31.36±3.50)μm,呈分散排列,导管间由大量木纤维组成,中间有薄壁细胞组成放射状木射线,根为四或五原型,中央无髓部。
2.1.3 天台鹅耳枥侧根的解剖结构 天台鹅耳枥主根上产生各级侧根。观察侧根的结构特征,其木质部为四或五原型,侧根原基发生于木质部放射角所对的中柱鞘细胞(图1-3)。从侧根横切面观察,侧根已产生的次生结构,周皮的木栓层4 ~ 6层,栓质化程度很高,染色很深;韧皮部中韧皮纤维较发达;次生木质部发达,导管数量多,木射线发达,排列呈十分规则的放射状。
2.1.4 天台鹅耳枥茎的解剖结构 天台鹅耳枥茎灰褐色有突起的皮孔,茎的横切面近圆形,具典型双子叶植物茎的结构特点,茎由外向内为周皮、皮层、韧皮部、形成层、次生木质部、初生木质部及髓组成(图 1-4,图1-5)。
茎次生构造形成时,表皮下皮层细胞产生木栓形成层,木栓形成层向外形成木栓层,其细胞3 ~ 4层排列扁化,壁栓质化,具有防止水分过多蒸发的作用;向内产生2 ~ 3层栓内层细胞。皮层位于周皮之内,具厚角组织3 ~ 5层,薄壁细胞8 ~ 10层,薄壁细胞长×宽 =(27.55±3.50)μm×(17.96±1.28)μm,薄壁细胞含晶体(图1-5)。
茎的次生维管组织是维管形成层平周分裂的结果,形成层细胞3 ~ 5层,为扁长形细胞,向外分化成次生韧皮部,向内分化成次生木质部。
次生韧皮部位于初生韧皮部内,由筛管、伴胞、韧皮纤维、韧皮薄壁细胞和韧皮射线组成,在次生结构中所占比例较少。在次生韧皮部外侧,有成团的韧皮纤维,韧皮纤维厚度为(22.24±2.32)μm,由厚壁细胞组成,排成一圈;韧皮射线呈直线排列,贯穿次生韧皮部。
次生木质部位于初生木质部外侧,所占面积较大,是茎的次生结构的主要部分。它的组成和初生木质部相似,主要由导管和木纤维组成,导管横径较大,导管直径为(22.86±3.58)μm,次生木质部显然要比初生木质部显得浓密得多,且木质化明显,在横切面上有木射线,显示较强的输导水分能力。

图1 天台鹅耳枥芽、根、茎和叶的解剖结构Figure 1 The anatomical structures of bud, root, stem and leaf of C. tientaiensis
茎的中心部分是髓(图1-6),髓的周围部分有紧密排列的厚壁小细胞组成的环髓带,髓部由薄壁细胞组成,细胞较大,薄壁细胞长(41.90±1.39)μm×宽(30.65±2.24)μm,可见簇晶。
2.1.5 天台鹅耳枥叶的解剖结构 天台鹅耳枥单叶互生,叶椭圆形或卵形,叶具网状脉序,边缘具重锯齿,叶正面近无毛,背面沿脉疏被长柔毛、脉腋间有簇生的髯毛,其余无毛。叶为完全叶,托叶早落,叶柄长6 ~ 12 mm,上面沟槽内密被长柔毛。
天台鹅耳枥的叶片为典型的异面叶,从叶片的横切面上看,由表皮、叶肉和叶脉所构成,叶厚度为(165.90±20.50)μm。表皮细胞排列紧密整齐,细胞多为无规则型,垂周壁深波状弯曲,上表皮的细胞较下表皮的细胞大;气孔集中在下表皮,为不规则型,由两个半月形的保卫细胞组成,下表皮气孔器密度平均(28.82±2.50)个/mm2(图1-7、图1-8)。叶片的上、下表皮各为1层(图1-9、图1-10),上、下表皮的细胞都角质化,形成连续的角质层,上表皮厚度为(12.12±0.45)μm,下表皮厚度为(10.50±2.35)μm。叶肉由栅栏组织和海绵组织构成(图1-9),栅栏组织厚度为(70.60±10.45)μm,栅栏组织细胞为1层排列整齐的长柱形细胞,薄壁细胞含晶体,细胞内含丰富的叶绿体,是天台鹅耳枥进行光合作用的主要场所;海绵组织由3 ~ 6层细胞构成,海绵组织厚度为(66.50±12.20)μm,细胞形状不规则,排列疏松,细胞间空隙大,邻近的细胞之间为水平连接,成网状,细胞所含叶绿体较栅栏组织的少。
观察中脉横切面(图1-11),中脉维管束较发达,近轴面三个维管束呈扇形排列,远轴面一个大维管束近圆形;其上、下部皆有数层厚角组织,中脉两面均突出形成脊。维管束的木质部在近轴面,而韧皮部在远轴面,并有形成层;木质部所占面积较大,而韧皮部则较小;维管束外面围绕着一圈由2 ~ 5层厚壁组织细胞(纤维)构成的维管束鞘,维管束鞘细胞木质化加厚,相当发达。
小型叶脉只含有一个维管束(图1-9,图1-12),其韧皮部有显著的薄壁组织细胞。维管束外面围绕着一层紧密排列的维管束鞘,由薄壁细胞组成。维管束鞘并不直接与表皮层相连,而是通过薄壁组织细胞与表皮层连接,这部分的细胞区域称为维管束鞘延伸区[6],这些薄壁细胞都不含有叶绿体。
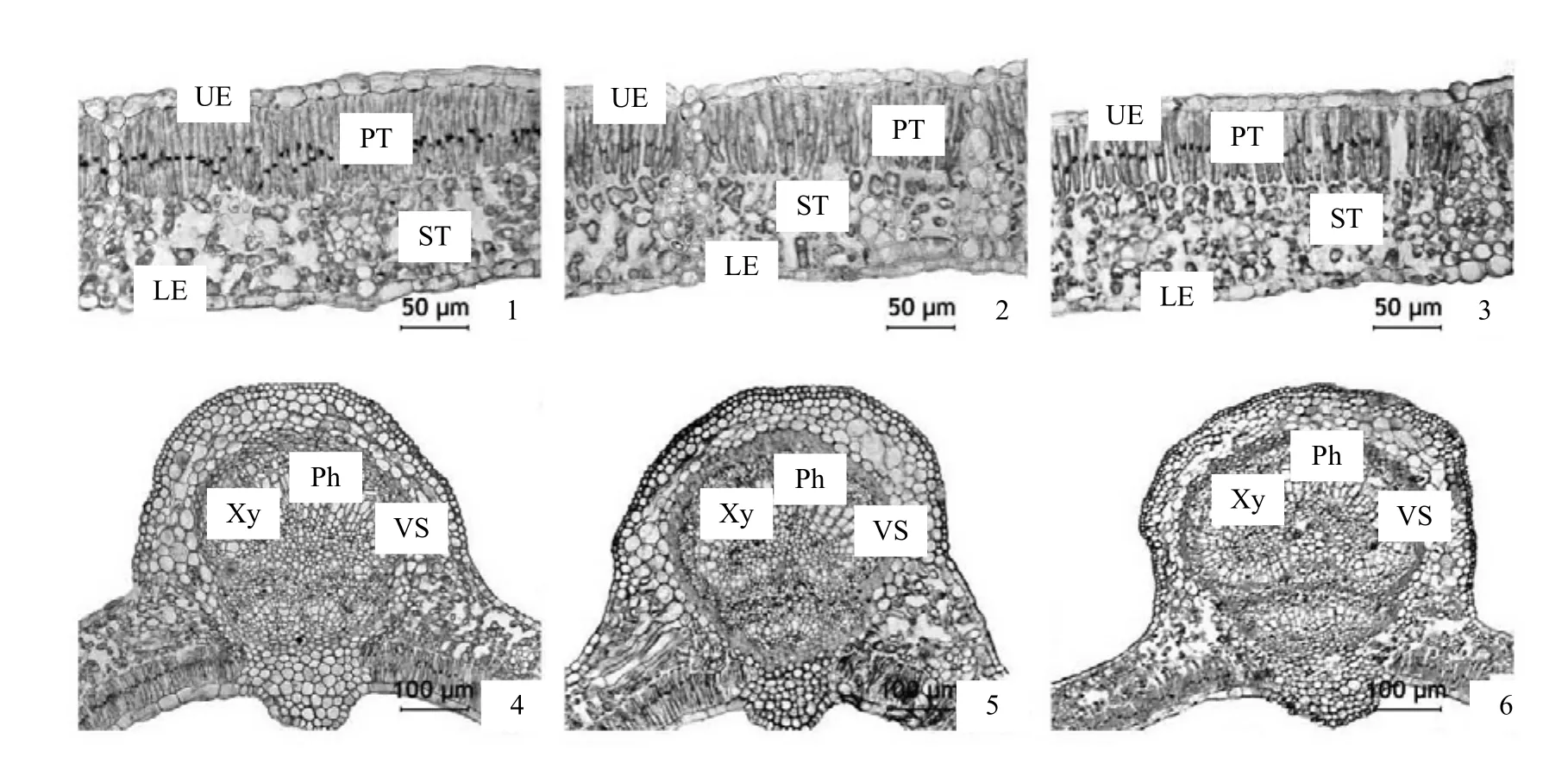
图2 生长环境光强对天台鹅耳枥叶的结构和主脉结构的影响Figure 2 Effects of the environmental light intensities on leaf and main vein microstructure of C. tientaiensis
2.2 生长环境光强对天台鹅耳枥叶解剖学特征的影响
天台鹅耳枥叶片的结构特征表现为典型的异面叶,由表皮、叶肉和叶脉所构成,叶脉横切面维管束呈圆状,主脉由1 ~ 4个维管束构成。
2.2.1 生长环境光强对天台鹅耳枥叶表皮组织结构的影响 天台鹅耳枥叶有表皮毛和角质膜,表皮细胞排列较为平整,大小也较为一致;生长在林缘和林下环境,天台鹅耳枥叶的表皮细胞形状及排列方式没有改变,但其厚度均变薄,其中林下环境其表皮细胞变薄更为明显(图2-1、图2-2、图2-3和表1),这与有利于吸收更多的光照以适应弱光环境有关。
2.2.2 生长环境光强对天台鹅耳枥叶肉组织结构的影响 低、弱光环境对天台鹅耳枥叶的结构影响较大。天台鹅耳枥在林窗较高光强下叶小而厚,叶肉组织分化明显,含有较多的细胞,叶片横切结构有栅栏组织1层,细胞均比较细长,呈长条形或圆柱形,海绵组织4 ~ 6层排列疏松(图2-1);在林缘和林下较低光强下天台鹅耳枥叶肉栅栏细胞排列疏松,海绵组织细胞间隙变大,细胞短而圆,叶片也较薄(图2-2、图2-3)。林窗天台鹅耳枥叶的主脉维管束发达,近轴面二个维管束呈扇形排列,远轴面一个大维管束近圆形,维管束鞘细胞由2 ~ 4层厚壁组织细胞构成(图2-4);林缘叶的主脉维管束较发达,近轴面有一大一小维管束呈扇形排列,远轴面有一个大维管束(图2-5);林下叶的主脉维管束近轴面有一个小维管束,远轴面有一个大维管束(图2-6),林下弱光环境叶脉维管束不发达。
就叶片厚度而言,低、弱光环境下天台鹅耳枥的叶片厚度明显变薄,林窗叶片厚度是林下叶片厚度的 1.27倍左右,其栅栏组织与海绵组织厚度之比均大于 1,海绵组织排列都较为疏松,不同生长环境光强叶片的栅栏组织与海绵组织厚度比值存在差异(表1)。

表1 生长环境光强对天台鹅耳枥叶的解剖学特征的影响Table 1 Effects of the environmental light intensities on anatomical characteristics of C. tientaiensis leaves
3 讨论
天台鹅耳枥生 长 于天台县华顶山溪谷两侧和山区林中,分布在海拔860 ~ 920 m处,年降水量1 700 mm,平均相对湿度85%以上,年平均气温 1 3℃,无霜期230 d,属中亚热带气候,温暖潮湿。山地土壤系水成 岩及火成的花岗岩母质上发育的山地黄壤土。本区特有的物种有天台鹅耳枥、华顶杜鹃、七子花等,并在浙江范围内以此区为主要分布区[2]。
植物与其生长的环境是一个整体,环境对植物的生长作用影响了植物的形态构成,营养器官的形态结构特征最能体现植物与生境的相互作用关系[7~8]。
天台鹅耳枥叶 肉组织和茎的皮层薄壁细胞含有晶体,茎的髓部薄壁细胞含簇晶,细胞中晶体的存在,可以改变细胞渗透压,提高吸水和持水力,同时也在植物组织中起到了支持作用[9]。
叶片是植物 进 化过程中对环境变化 较 敏感且可 塑 性较大的器官, 在不同选择 压力下已经形 成 各种适应类型,其结构特征最能体现环境因子的影响或植物对环境的适应[10~11]。光是植物进行光合作用的基本能源,同时影响植物光形态建成。叶片在 较高光强下加 厚 ,叶肉组织 细胞数量增加 ; 在弱光下 叶肉组织细胞 排 列疏松,叶片变薄[12];高至低光环境下 (林窗、林缘至林 下 ), 栅栏组织与海绵组织 厚 度之比为1.13 ~ 1.03,栅栏组织与叶片厚度之比为0.47 ~ 0.41。天台鹅耳枥叶的 主脉维管束发达,叶片具有 发达的网状叶脉和维管束鞘 细胞,可增强从叶基部向叶肉 细胞的水分传输,而机 械 组织的增强 则能在逆境 中 抵御物理损伤 和因失水萎 蔫 而造成的不良影响[13]。由此说明,天台鹅耳枥属耐荫植物类型,这与天台鹅耳枥分布范围极其狭窄,一般分布于溪谷林下环境相适应。另一方面,天台鹅耳枥的气孔主要集中在下表皮,叶表皮细胞角质化,根和茎均具有明显的角质层,芽有覆瓦状鳞片包被,使得它的蒸腾作用减弱,说明其保水能力比较好,能适应山区林中环境。
天台鹅耳枥的营养器官结构表现出较耐荫的特征,在进行天台鹅耳枥生物多样性的迁地保护中[14],幼苗的管理应给予适当的光照条件,注重其保水器官的保护,引种栽培天台鹅耳枥时最好选择在溪谷林下以促进其正常生长,确保物种繁衍。
[1]章绍尧,丁炳扬. 《浙江植物志》总论卷[M]. 杭州:浙江科学技术出版社,1993. 246-251.
[2] 金则新,林苹,张美娟. 台州珍稀濒危植物区系分析[J]. 浙江林业科技,2005(6):55-58.
[3]胡绍庆,丁炳扬,陈征海. 浙江省珍稀濒危植物物种多样性保护的关键区域[J]. 生物多样性,2002,10(1):15-23.
[4] 陈之端. 桦木科植物的系统发育和地理分布[J]. 植物分类学报,1994,32(1):1-31.
[5] 林加涵,魏文铃,彭宣宪. 现代生物学实验(上册)[M]. 北京:高等教育出版社.
[6]刘穆. 种子植物形态解剖学导论[M]. 北京:科学出版社,2001. 164-176.
[7] 王勋陵,马骥. 从旱生植物叶结构探讨其生态适应的多样性[J]. 生态学报,1999,19(6):787-792.
[8] Klich M G. Leaf variations in Elaeagnus angustifolia related to environment heterogeneity[J]. Environ Exp Bot,2000,44(3):171-183.
[9] 周智彬,李培军. 我国旱生植物的形态解剖学研究[J]. 干旱区研究,2002,19(1):35-40.
[10] 李芳兰,包维楷. 植物叶片形态解剖结构对环境变化的响应与适应[J]. 植物学通报,2005,22(S1):118-127.
[11] Castro-Diez P, Puyravaud J P, Cornelissen J H C. Leaf structure and anatomy as related to leaf mass per area variation in seedlings of wide range of woody plant species and types[J]. Oecologia,2000(124):476-486.
[12]Hans Lambers, F.Stuart Chapin Ⅲ, Thijs L.Pons. Plant Physiological Ecology[M]. New York:Springer Press,1998. 8-57.
[13] 王勋陵,王静. 植物形态结构与环境[M]. 兰州:兰州大学出版社,1989. 105-138.
[14] 张文辉,祖元刚,刘国彬. 十种濒危植物的种群生态学特征极致危因素分析[J]. 生态学报,2002,22(9):1 512-1520.
