江西齐云山福建柏群落优势种群生态位动态特征
2010-05-15杨清培季春峰杨光耀黄声亮
杨清培,季春峰,杨光耀,刘 骏,黄声亮
(1.江西农业大学 江西省竹子种质资源与利用重点实验室,江西 南昌 330045;2.江西齐云山自然保护区管理局,江西 崇义 341315)
福建柏(Fokieniahodginsii)为柏科福建柏属植物,主要分布于浙江、福建、江西等地[1].它不但在柏科植物系统发育研究方面有重要的科学意义,而且是我国南方优质用材树种和园林树种[1-3].由于长期不合理开发,现天然林资源已近枯竭,已被列为国家二级重点保护植物[2].目前相关学者对其群落结构[4-6]、种间关系[7]、生长规律[8]、引种栽培[9]、人工林生物量[10]、生产力[11]以及碳汇功能[12]等方面研究较多,应用生态位概念更能较好的定量反映生物与环境间的相互作用关系[13-15],然而对其生态位的研究未见报道.本文通过对江西齐云山自然保护区福建柏群落优势种群发展过程中生态位动态特征进行研究,以期揭示其群落结构与功能、群落动态演替、种内种间关系及种群群落地位,为该种质资源保护、栽培利用等提供理论参考.
1 材料与方法
1.1 研究区概况
齐云山自然保护区位于崇义县西北边缘地带,属罗霄山脉南端的诸广山区,地理位置为东经113°55′~114°08′,北纬25°44′~25°54′,西北长22 km,东西宽14 km,总面积为18 870 hm2.地势为西高东低,最高峰(即齐云山顶峰)为2 061.3 m,最低(香炉坝)为450 m.保护区属湿润型季风气候带,常年温暖湿润,雨量充沛,年平匀降雨量为1 750 mm,平均气温17℃,绝对最高气温38.8℃,最低气温-8℃,年平均日照时数1 374.8 h,年有效积温为4 007℃.土壤以山地黄红壤和黄棕壤土为主,植被以常绿阔叶林为主,但嵌有较大面积针叶林,尤其福建柏林数量众多、分布较广,分别达到75.26万株和1 485.0 hm2[8].
1.2 研究方法
1.2.1 样地调查 2007~2008年,在对齐云山森林群落全面踏查的基础上,根据海拔高度、群落类型和立地条件等环境因子,设立8块福建柏群落样地,采用相邻格子法把每个样地分成10 m×10 m的小样方(共计64个),并进行群落调查.所有木本植物进行每木检尺,记录植物种名、胸径(幼树、幼苗测地径)、树高、冠幅、枝下高等.同时测定每块样地的海拔、坡度和坡向等环境因子.
1.2.2 数据处理 将8个样地采用立木级代表年龄的方法,将群落分成4个年龄层片,即Ⅰ-Ⅱ(DBH<2.5 cm幼苗幼树)、Ⅲ(2.5 cm≤DBH<7.5 cm小树)、Ⅳ(7.5 cm≤DBH<22.5 cm壮树)和Ⅴ(DBH≥22.5 cm大树)层片[8,16].先计算各层片中物种重要值,再从各层片中分别选取前15位物种,以调查的10 m×10 m小样方作为资源状态,进行生态位相关指标的计测.
1)重要值 它是计量种群在群落中重要性的常用指标[16],其计算式为:
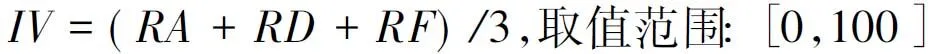
(1)
式(1)中,IV为物种重要值,RA为相对多度,RD为相对显著度,RF为相对频度.
2)生态位宽度 利用物种多度作为物种数据的综合指标.为了考虑每一资源状态的丰富度和可获得性的不同,认为采用Hurlbert指数标准生态位宽度较好[17]:
(2)
(3)
3)生态位重叠 采用Hurlbert指数计算生态位重叠[17]:
(4)
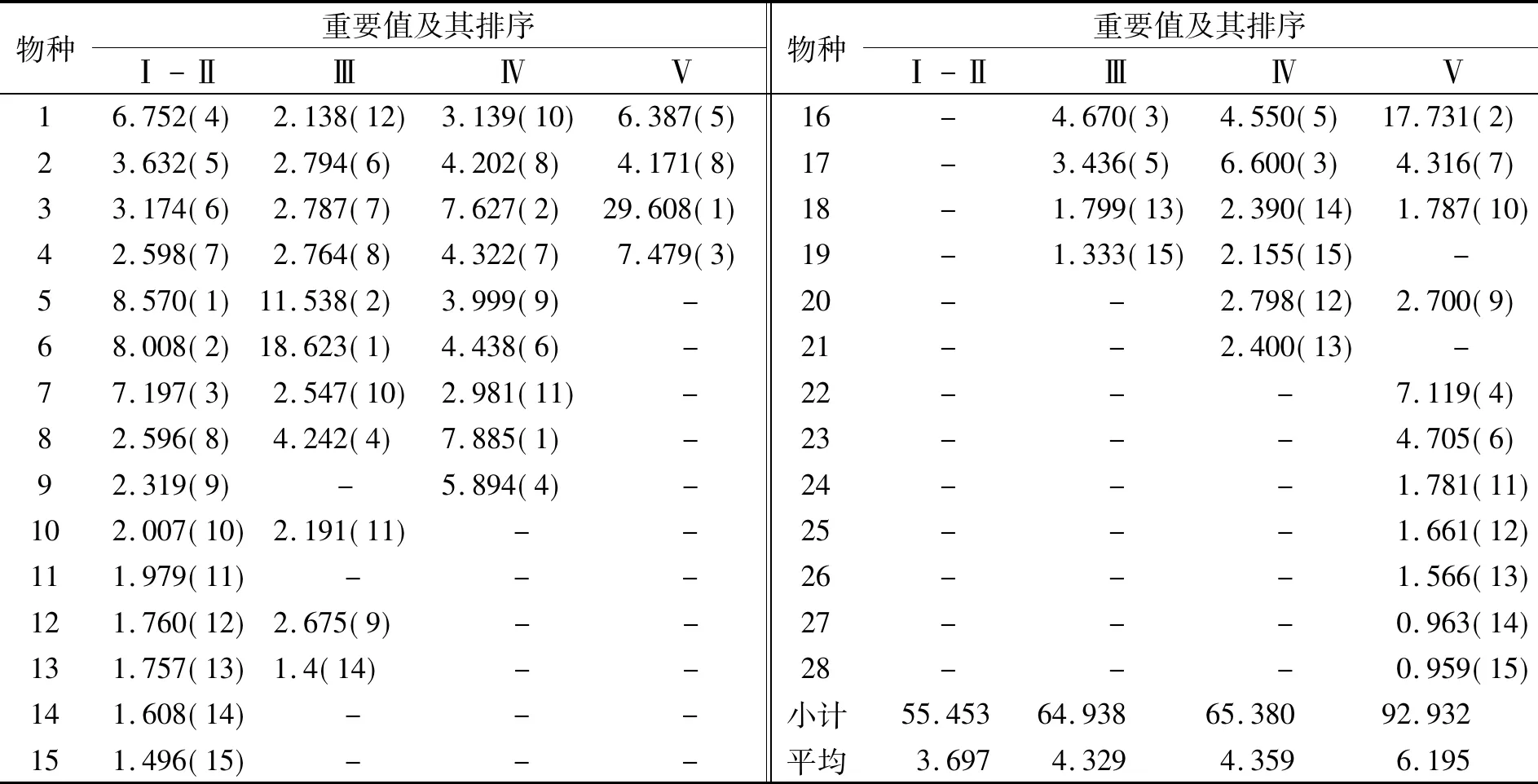
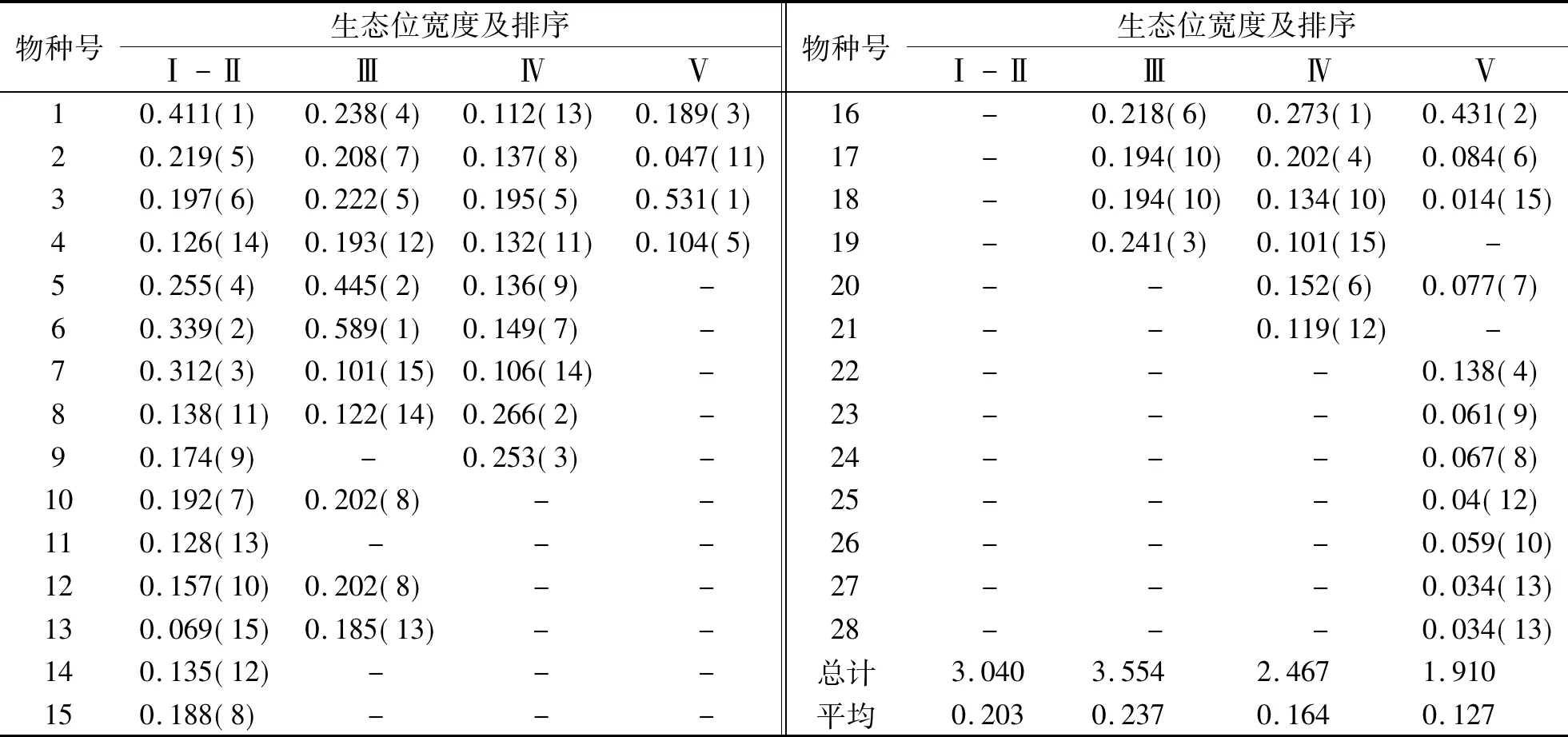
式(4)中L(i,j)为种i对种j的生态位重叠指数,且L(i,j)=L(j,i);Pki,Pkj,ak同式(2).L(i,j)=0时两物种不共有资源状态;L(i,j)=1时,两物种成比例地使用每一资源状态;L(i,j)>1时,两物种集中使用某些资源状态且喜好程度一致.本文将生态位重叠分成高度重叠(L(i,j)≥1)和部分重叠(0 福建柏群落中共调查到木本植物250种,隶属112科156属.第Ⅰ-Ⅱ、Ⅲ、Ⅳ和Ⅴ层片各有物种192、151、78和24种.群落各层片优势物种及其重要值(见表1).由表1可知,随年龄增长,各层片优势物种不断发生变化.第Ⅰ-Ⅱ层片中以马银花(种5)、鹿角杜鹃(种6)和深山含笑(种7)等,其重要值分别为8.570、8.008和7.197;第Ⅲ层片主要鹿角杜鹃18.623、马银花11.538和银木荷(种16)4.670;第Ⅳ层片却主要是假木荷(种8)7.885、甜槠(种3)7.627和黄山松(种17)6.600;发展至第Ⅴ层片,变成以甜槠(种3)29.608、银木荷17.731和红楠(种4)7.479等占有明显优势.同时,优势物种重要值所占比例逐渐增大,且不断向少数物种集中.第Ⅰ-Ⅱ层片15种优势物种重要值之和与最大值分别为55.453和8.570,第Ⅲ层片为64.938和18.625,至第Ⅴ层片时分别增至92.932和29.608.说明物种间经过激烈竞争,此消彼长,少数适应能力强的物种相对数量和生物量占有明显优势,逐渐成为各层片优势种,甚至群落建群种. 随年龄增长,优势物种生态位宽度的消长关系也在不断变化(表2).第Ⅰ-Ⅱ层片福建柏(种1)生态位宽度最大,其值为0.411,其次为鹿角杜鹃0.339、深山含笑0.312、马银花0.255等;第Ⅲ层片,鹿角杜鹃最大,值为0.589,其次是马银花0.445,多脉青冈(种19)0.241、福建柏0.238等;第Ⅳ层片,生态位最宽的依次是银木荷0.273、假木荷0.226、舟柄茶(种9)0.253等;至第Ⅴ层片,甜槠生态位最大0.531,其次是银木荷0.431、福建柏0.189等.生态位宽度的变化表现出大多数增长型种群逐渐减小而衰退型种群逐渐增加的趋势,这也是对不同种群在群落中生态适应能力和资源利用能力的量化反映. 表1 群落各层片优势种重要值及动态变化 同时还发现,随年龄增长,各层片生态位宽度总和(或平均值)呈现出下降趋势.第Ⅰ-Ⅱ与第Ⅲ层片优势种生态位宽度总值分别为3.040和3.554, 但至第Ⅳ、Ⅴ层片时则依次下降至2.467和1.910,相应地,其平均值也由0.203下降至0.164和0.127. 福建柏4层片15优势树种分别构成105种对的半矩阵(见图1)及生态位重叠变化趋势(图2).可知,随年龄增长,生态位重叠种对逐渐减少,生态位重叠均值逐渐降低;相反,生态位分离种对逐渐增加. 第Ⅰ-Ⅱ层片,生态位重叠种对共计100对,占95.238%,其中部分重叠66对(62.857%)、高度重叠34对(32.381%).重叠值较大的种对依次是假木荷-日本杜英(8-12)、长苞铁杉-老鼠矢(2-15)、柃木-日本杜英(11-12)、深山含笑-日本杜英(9-12)、满山红-老鼠矢(13-15).相比较而言,日本杜英、老鼠矢、狭叶香桂(14)与其他物种的重叠度较高,而甜槠、深山含笑与其他物种的重叠度较低.而生态位分离种对5对,占4.762%,仅鹿角杜鹃-深山含笑、红楠-假木荷等少数种对.说明此时大多数物种间关系复杂,种间竞争激烈. 表2 福建柏群落各层片优势种群的生态位宽度 图1 福建柏群落各层片中优势种群生态位重叠特征(a:Ⅰ-Ⅱ层; b:Ⅲ层;c:Ⅳ层;d:Ⅴ层) 第Ⅲ层片,生态位重叠种对有所减少,生态位重叠种对减为94对.其中高度重叠减至25对,减少了9.52%.此层片生态位重叠较大的种对依次是:假木荷-日本杜英、满山红-南方铁杉、长苞铁杉-黄山松、银木荷-黄山松、满山红-多脉青冈、南方铁杉-多脉青冈.相比而言,多脉青冈、南方铁杉、满山红与其他物种的重叠度较高.生态位分离种对有所增加,为11对,占10.476%.此时,生态位重叠平均值也下降至0.857. 第Ⅳ层片,生态位重叠种对已减至77对,与上层比,高度重叠25对不变,但部分重叠52对,减少18对.重叠值最大种对是南方铁杉-多脉青冈,其次为长苞铁杉-深山含笑、银木荷-南方铁杉、假木荷-舟柄茶.南方铁杉、银木荷与其他物种重叠度较高,而深山含笑、舟柄茶与其他物种的重叠度较低.生态位分离种对已升至28对,达26.667%.同时,此层片生态位重叠平均值降至0.665. 图2 福建柏群落优势种群间生态位重叠动态 第Ⅴ层片,生态位重叠种对减少至34对,仅占15.238%,高度重叠与部分重叠种对数分别减至18和16对,分别占17.143%和15.238%.其中蓝果树-乌楣栲(25-27)重叠值最大,为11.900.其次为黄山松-南方铁杉(5.950)、福建柏-大果马蹄荷(1-24)、福建柏-马尾松(1-23)(2.975).生态位分离种对升至71对,已占总种对的67.619%,说明此时多数物种对环境资源利用已发生分化,表现出高度的异质性.同时发现,虽然重叠种对数减少,但重叠种对间的重叠值普遍增加,如蓝果树-乌楣栲,重叠值达11.90,黄山松-南方铁杉为5.950,都要远远超过幼苗幼树、小树时期最大的重叠值,这可能是经过长期激烈竞争后,种间彼此适应,各自从不同角度与侧面利用环境资源[18,19],从而形成相对稳定群落. 随年龄增长,福建柏种群的重要值与生态位宽度都而呈先降后升的“V”形趋势,同时,与越来越多的伴生树种发生生态位分离(表1,表2,图3(a,b)).第Ⅰ-Ⅱ层片,即幼苗幼树时期,福建柏种群重要值为6.752,然后减至第III层片的2.138(列于此层第12位),然后逐渐回升至第V层片的6.387(列第3位).生态位宽度也表现出相同的趋势,第I层为0.411(列第1位),然后递减至第Ⅳ层片的0.112(列此层第13位),最后在第Ⅴ层回升至0.189(列第3位).说明在福建柏种群发展过程中,开始占有相当优势且生态位较广,然后逐渐失去一些资源位,直到在大树时期竞争力才逐渐增强,并在群落中起着一定的主导作用. 图3 福建柏生态位宽度及与伴生种群生态位重叠动态 与时同时,在第Ⅰ-Ⅱ层片中,福建柏与伴生树种都有生态位重叠,表明它们有着较为相似的资源利用方式,对其发展有较强的竞争作用,尤其是与鹿角杜鹃和狭叶香桂(种14)的竞争最大.第Ⅲ层片全部伴生树种都仍与之生态位重叠,而重叠程度进一步加大,高度重叠的物种比例由14.186%增加到28.571%,说明此时福建柏仍未能摆脱伴生树种的竞争;第Ⅳ层片(即壮树时期),福建柏开始与部分伴生树种(如深山含笑、多脉青冈)发生生态位分离,生态位重叠种对比例下降.进入第Ⅴ层片(大树龄级)后,福建柏与更多的树种发生生态位分离,占总物种的50.0%,但仍与部分物种有较大的生态位重叠.这可能是经过长期激烈竞争,福建柏与部分物种(红楠、黄山松、南方铁杉等)在环境资源利用方式出现了完全分化,同时又与另一部分物种(如大果马蹄荷、饭甑青冈、小叶青冈、马尾松)彼此相互适应,形成了相对稳定的种间关系[18]. 群落发展与群落内种群消长互为因果[20].随年龄增长,福建柏群落各层片物种种类、重要值和生态位宽度都发生相应变化.由第Ⅰ-Ⅱ层片至第V层片,优势物种由马银花、鹿角杜鹃、深山含笑等逐渐演变成甜槠、银木荷、红楠等,而且其平均重要值逐渐增大,由3.697增至6.195;平均生态位宽度在逐渐减少,由0.203减小至0.127;同时,优势种作用越来越明显,重要值或生态位宽度值向少数物种集中,如第一重要值由8.570增至29.608,第一生态位宽度占总宽度百分比由13.520 %增至27.801%.这说明随年龄增长,多数物种对环境资源的利用越来越少,逐渐从不适应的环境中退出,并让位给那些适应能力强物种,所以少数物种在群落中的地位和作用越来越明显,成为群落的共优种或建群种. 从理论上说,当不同物种利用或共同占有同一资源时,就会出现生态位重叠现象,而且生态位重叠越大,竞争就越激烈[16,21].但从低维生态位上来说,生态位重叠可能有两种情况:一是可能资源共享而竞争,二是可能是资源互补而共存[22].本研究中,随年龄增长,生态位重叠种对数量逐渐减少,平均重叠程度不断降低;相反,生态分离种对逐渐增加.从第Ⅰ-Ⅱ层片至第Ⅴ层片,生态位重叠种对比例由95.238%依次降为89.524%、73.333% ,最后仅占32.381%;同时其平均重叠值由0.876依次降为0.848、0.665和0.526;生态位分离种对却相应地由4.762%,依次增加到10.476%、26.667%和67.619%.所以从群落发展总体上看,物种间对环境资源利用方式逐渐出现分化.然而,少数物种间在此过程中生态位重叠度越来越大.从第Ⅰ-Ⅱ层片至第Ⅴ层片生态位重叠最大值由3.703(假木荷-日本杜英)依次增至4.541(假木荷-日本杜英)、4.922(南方铁杉-多脉青冈)和11.900(蓝果树-乌楣栲).因为至第V层片,这些林木都是几十年甚至上百年的大树,因此,本文认为尽管它们具有相同或相似的环境需求,但是它们却属于资源利用互补. 就福建柏种群而言,它在群落中的地位和作用在不断发生变化.随年龄增长,其重要值和生态位宽度都基本上呈现“高-低-高”的“V”型趋势,从第Ⅰ-Ⅱ层片至第V层片,其重要值依次为6.752、2.138、3.139和6.387,生态位宽度值依次为0.411、0.238、0.112和0.189.同时,福建柏与越来越少的伴生树种生态位重叠,而生态位分离的物种逐渐增加.这种情况的出现与福建柏的生态习性分不开.福建柏种子传播广、萌发能力强,且幼年耐荫[5],所以第Ⅰ-Ⅱ层片生态位宽度较大、重要值较高,但它毕竟它是中性偏阳喜光、喜水肥的树种[2,4].深处阔叶树下,有众多树种与之争夺光资源,或自身不能长期适应干旱贫瘠的生境,至第Ⅲ、Ⅳ层片便从许多生境退出,表现为生态位宽度和重要值都有所下降.至第Ⅴ层片,某些水肥充足、土层深厚、排水良好生境的福建柏摆脱伴生树种的竞争,而居群落冠层,并与马尾松、大果马蹄荷或小叶青冈等树种资源利用互补,从而形成相对的针阔混交林或小片纯林.这种变化规律与本地福建柏增长型种群特点和发展规律相一致[8].因此,福建柏种群生态位变化能较好地解释群落发展过程中它对环境的响应以及种间关系等生态过程的演变.在福建柏保护实践中,可以利用种群生态位的计测,加强对不同种群在群落中的地位的认识,合理进行人工干扰和抚育,加速群落向稳定性的目标群落演替与发展. 参考文献: [1]侯伯鑫,林峰,余格非,等.福建柏资源分布的研究[J].中国野生植物资源,2005,24(1):58-59,64. [2]林峰,侯伯鑫,杨宗武,等.福建柏属的起源与分布[J].南京林业大学学报:自然科学版,2004,28(5):22-26. [3]陈祖松.福建柏人工林木材物理力学性质的试验研究[J].福建林学院学报,1999,19(3):223-226. [4]邓贤兰,龙婉婉,许东风,等.井冈山自然保护区福建柏群落的研究[J].热带亚热带植物学报,2008,16(2):128-133. [5]侯伯鑫,余格非,林峰,等.福建柏天然林群落类型研究[J].湖南林业科技,2004,31(6):33-35. [6]哀建国,丁炳扬,于明坚.凤阳山白然保护区福建柏种群结构和分布格局研究[J].西部林业科学,2005,34(3):45-49. [7]曹培健,丁炳扬,李伟成,等.凤阳山福建柏群落主要种群种间联结性研究[J].浙江大学学报:理学版,2006,33(6):676-681,687. [8]杨清培,江西齐云山长苞铁杉与福建柏生长规律及种群特征研究[J].江西农业大学学报,2009,31(2):264-269. [9]侯伯鑫,林峰,余格非,等.福建柏地理种源开花与结实变异规律的研究[J].植物遗传资源学报,2005,6(2):163-167. [10]杨宗武,谭芳林,肖祥希.福建柏人工林生物量的研究[J].林业科学,2000,36(专刊1):120-124. [11]廖涵宗.福建柏人工林生态系统生产力研究[J].南京林业大学学报,1995,19(2):59-66. [12]何宗明,李丽红,王义祥,等.33年生福建柏人工林碳库与碳吸存[J].山地学报,2003,21(3):298-303. [13]Silvertown J. Plant coexistence and the niche[J]. Trends in Ecology and Evolution,2004,19(11):605-611. [14]Wissinger SA. Niche overlap and potential for competition and intraguild predation between size structured populations[J]. Ecology,1992,73(4):1 431-1 444. [15]张金屯.数量生态学[M].北京:科学出版社,2004:110-120. [16]王伯荪,余世孝,彭少鳞,等.植物群落学实验手册[M].广州:广东高等教育出版社,1996:107-115. [17]张文军.生态学研究方法[M].广州:中山大学出版社,2007:104-111. [18]闫淑君,洪伟,吴承祯,等.万木林中亚热带常绿阔叶林林隙主要树种的高度生态位[J].应用与环境生物学报,2002,8(6):578-582. [19]赵永华,雷瑞德,何兴元,等.秦岭锐齿栎林种群生态位特征研究[J].应用生态学报,2004,15(6):913-918. [20]祝廷成,钟章成,李建东.植物生态学[M].北京:高等教育出版社,1988:204-213. [21]康冰,刘世荣,史作民,等.南亚热带人工马尾松林下植物组成特征及主要木本种群生态位研究[J].应用生态学报,2005,16(9):1 786-1 790. [22]刘金福,洪伟.格氏栲群落生态学研究——格氏栲林主要种群生态位的研究[J].生态学报,1999,19(3):347-352.2 结果与分析
2.1 优势种群重要值及其动态变化
2.2 优势种群生态位宽度及其动态变化
2.3 优势种群间生态位重叠与动态变化
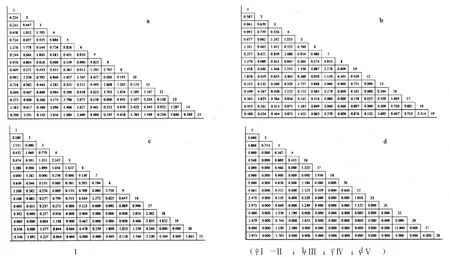
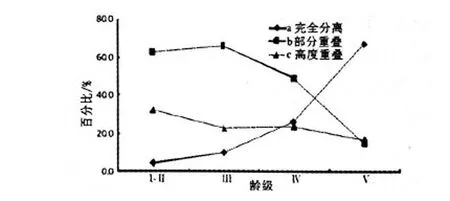
2.4 福建柏与其伴生种群生态位动态

3 小结与讨论
