声音在耳内的信号转导及其分子生物学机制(4)
2010-01-25汤浩石丽娟于利曹宇
汤浩 石丽娟 于利 曹宇
2 毛细胞信号转导的几个特点
2.1信号转导过程中无第二信使参与 已知毛细胞的直接的机械-电转导是很迅速的,原因在于毛细胞的信号转导是无第二信使干涉的。与此相反,许多其它的感觉感受器,比如视觉感受器和嗅觉神经元,在转导过程中必须有环状核苷酸或者其它的第二信使参与。这一转导过程的优越性在于它的酶器,并且在代谢通路中产生反馈,因此,允许这些感觉感受器具有快速的适应或减感的控制性。
那么无第二信使干涉的转导的选择性优势是什么呢?答案就在反应的速度上。毛细胞的控制要比脊椎神经系统的其它感觉感受器快得多,并且比神经的本身还要快。为了适应生物性相关刺激的频率,毛细胞的转导速度必须很迅速。对空气中声音的运动和收发声音的器官(如鼓膜等)研究表明,最理想的听觉转导发生在0.1~100 kHz的频率范围内。过高的频率在空气中传播的效果较差;过低的频率是无效率的传播,并且只能被大小适宜的动物所捕获。
声源的位置更严格地限定了转导速度。如果一个声源直接位于一个动物的一侧,发出的声波到达近侧耳将比到达远侧耳更快一些。对于人类,这一延迟至多大约是700 μs。人和猫头鹰可以在更小的一过性延迟时间(大约是10 μs)内确定声源的位置。由此可见,毛细胞必须能够以微秒水平的速度探测到声波的波形。毛细胞对高频刺激的反应充分表明其转导通道的开闭是非常快的。由于哺乳动物能够对大于100 kHz的频率作出反应,因此毛细胞很可能更快地控制等级状次序的速度。但是,低等动物却对相对较低频率的声波敏感。在24℃时,对于适宜强度的刺激的反应的时间常数只有80 μs。
毛细胞的一过性反应决定了它们对刺激的敏感性。但毛细胞的反应并不是不变的,其机械敏感性有着不同的方式,一个给定的细胞对与之相适应的刺激反应最强。但一定范围的适当刺激(忽略低频输入),毛细胞有一个独特的适应方式来承受刺激。许多毛细胞能探测摆动的刺激并产生电共振现象,最后使得每个细胞调整到一个特定的频率。
2.2毛细胞可适应持久的刺激 不论它如何精密地生长,具有高度机械敏感性的纤毛束转导器都不可能始终平衡地保持在原位。一些机制必须用来均衡发育的不规则以及环境的变化,通过重新设置门控弹簧来适应纤毛束的静息位置。一个连续的调节过程校准了纤毛束的机械敏感性范围,对于瞬时的刺激在抵制大约一百万倍的静电输入的同时,它仍可以维持较高的敏感性。
调节作用表明,在拖延纤毛束感受器偏离的过程中感受器电位是逐步减弱的。这一过程同感受器持续敏感性的减感作用是不同的。但是,在延长的刺激作用下,毛细胞的机械敏感范围从纤毛束的静息位置移动到受刺激维持的偏移位置。适应出现在三等级的对称,要比机械电转导慢。当内淋巴浸润着纤毛束时,适应的时间常数接近或等于25 ms。适应的速度和程度随着接触到细胞顶部的液体中的Ca2+浓度的增加而增加[3,11]。
适应又是怎样发生的呢?由于在适应的过程中纤毛束的硬度会发生变化,因此这一过程明显包括通过门控弹簧对压力负担的调整。固定每个顶端连接的上部的插入斑在由活性分子发动机的调节下重新配置。纤毛束包括几个作为该角色的候补物质——不同形式的球蛋白,和沿着肌动蛋白丝并且与运动有关的分子。例如,免疫组织化学研究表明肌球蛋白Ⅰβ在立体纤毛顶端附近丛生,很可能是插入斑处。肌球蛋白Ⅵ和Ⅶa也可以出现在纤毛束中。这些同工酶的ATP酶活性受到许多相关的调钙素的调节,而它们的功能至今仍未被发现。许多这样的肌球蛋白分子可能通过升高细胞骨架肌动蛋白纤维,牵引顶端连接与它们的附着点维持了每个顶端连接的压力(图7)。
图8上方曲线显示纤毛束在受到正向的或反向的持续很长时间的偏移时的电学反应。一个阳性(正向)刺激的电学反应表现为开始是去极化,紧接着逐渐降低到一个平台期,在刺激停止时出现一个负尖峰。阴性(反向)刺激时表现出一个补偿性反应,在刺激过程中,感受器电位大大地减弱,但在末尾出现一个反弹。下面的模型图表示,立体纤毛偏移与门控弹簧的变化关系。阳性(正向箭头)刺激引起的纤毛束的移动增加了顶端连接的张力,从而开放了转导通道,正离子进入,对应于上面的去极化反应。然而,当刺激持续时,顶端连接的较高一侧的插入点沿着静纤毛向下移动,表明在适应过程中可以使每一个通道都能够关闭。而在阴性刺激过程中,通过顶端连接较高一侧的插入点又主动上移(见最右侧图再转到最左侧初始图),张力重新储存到初始的松弛状态的顶端连接中(又回到静息状态)。可见,当一个刺激逐步增加门控弹簧上的压力时,相应的转导通道开放,允许正离子流入。当Ca2+在立体纤毛细胞浆中聚集时,它们同调钙素相互作用,引起相应的肌球蛋白Ⅰβ分子产生更小的向上的力。因此,门控弹簧可以降低肌球蛋白分子向上的力,并最终使之缩短。当弹簧达到它的静息压力时,转导通道的关闭减少了Ca2+的流入,直至恢复到原来的水平,亦即使肌球蛋白产生的向上的力和弹簧使之产生的向下的力之间达到平衡。
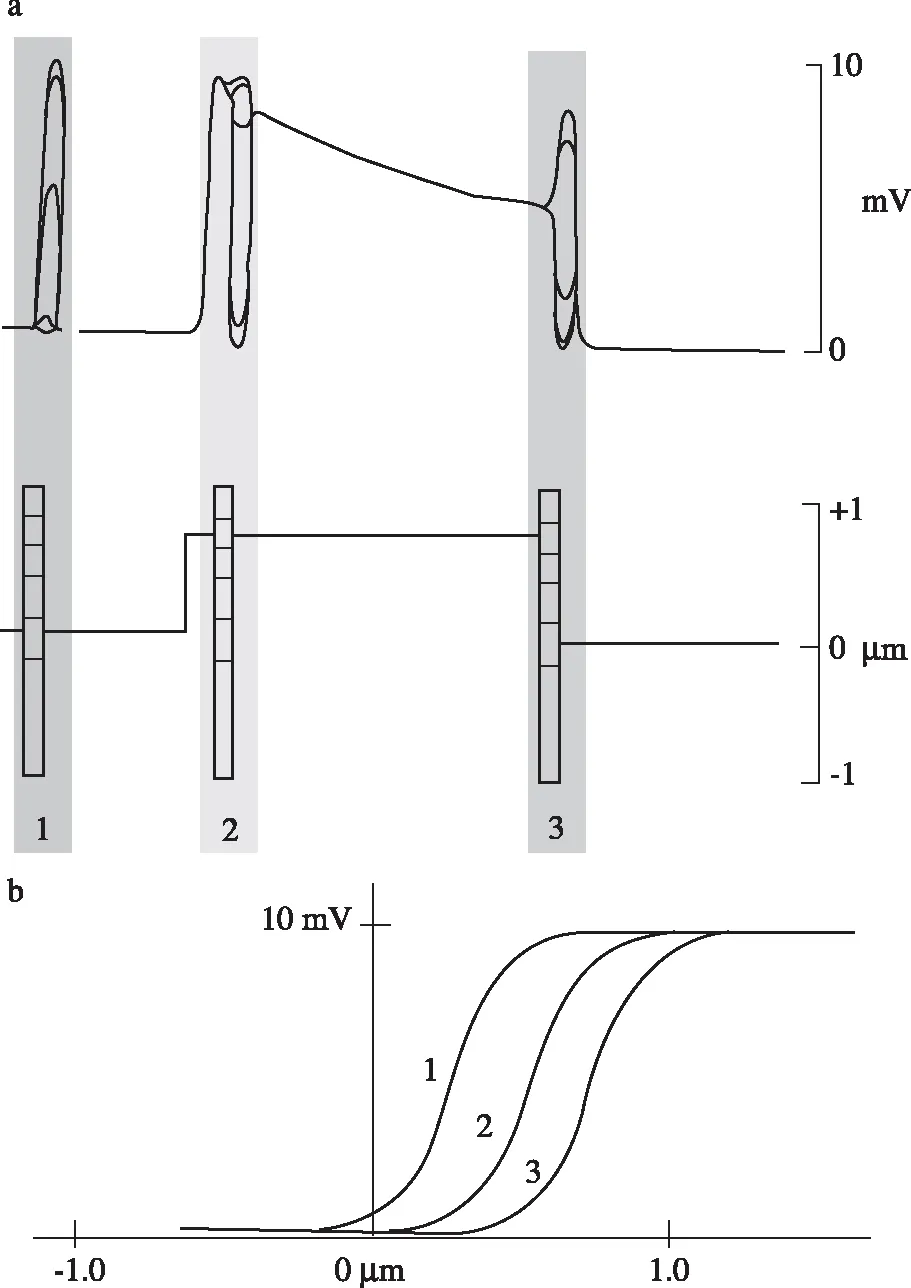
图7毛细胞的机械-电转导作用的适应现象
a.在一系列实验中,使纤毛束受到同属一组的偏移刺激及其产生的感受器电位。在(1)之前施加了不同强度的偏移刺激,而在纤毛束偏移过程中以两倍的强度维持偏移100 ms(2)~(3)。可见这一组的感受器电位显示在开始阶段出现一个快速的去极化,紧接着在维持刺激过程中感受器电位逐渐降低到一个平台期
b.在偏移前及偏移过程中,纤毛束的位移与电学反应间的关系。当适应现象开始时,纤毛束的位移与感受器电位间的“S”形的关系沿着横坐标变化,但曲线的形状和幅度没有实质性的改变。这个结果提示,在适应一个延长的刺激时,纤毛束的机械敏感性的迁移范围是朝向维持纤毛束举起时的机械敏感性位点的
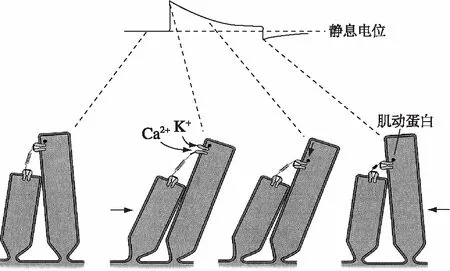
图8毛细胞的适应现象的模型
上方曲线为,纤毛束在受到正向的或反向的持续很长时间的偏移时的电学反应。下面的模型图表示立体纤毛偏移与门控弹簧的变化关系
2.3毛细胞调谐于特定的刺激频率 毛细胞必须与具有很低能量的听觉刺激竞争,如果刺激由一个周期性的信号组成,例如,纯音的正弦曲线压力,一个探测系统可以通过选择性地放大相应频率的反应加强它的信噪比(SNR)。目前已知,至少有两种细胞机制作用于这一任务的完成。
首先,纤毛束的机械特性使得它调整到一个特定的频率。这一普通的纤毛束频率依赖于它的机械性质,这一性质正如音叉的共鸣频率取决于它的弹性和音叉的质量。使得纤毛束恢复到静息位置的弹性物质包括门控弹簧和立体纤毛基部肌动蛋白支根。由于纤毛束运动通过一个粘性的介质,其粘性也使得运动严重受阻。此外,纤毛束调谐的相应的物质包括随纤毛束移动而摆动的内淋巴流。尽管纤毛束最佳反应是在一个特定的频率,但这与音叉在刺激停止后不能持续振动是不同的。
在不同动物听觉器官中,纤毛束的长度在沿着器官频率轴的方向上有系统的区别,对听觉的、振动的、加速的低频刺激作出反应的毛细胞具有最长的纤毛束,而感受最高频听觉信号的毛细胞具有最短的纤毛束,例如,在人耳蜗中,一个对20 kHz有反应的内毛细胞拥有4 μm的纤毛束。相对地,一个对20 Hz的刺激敏感的毛细胞拥有超过7 μm高的纤毛束。这种解剖学位置和细胞最敏感的频率之间的系统性的关系被称为张力映射或声调谐映射(tonotopic mapping)[3]。这一现象在听觉系统十分普遍。
电脑模型显示音叉机制使用不与盖膜接触的自立细胞来调整毛细胞。这些细胞包括人耳蜗的内毛细胞,它们是提供通过耳蜗神经传递大部分信息的感受器。立体纤毛的长度可能也影响纤毛束插入盖膜的那些毛细胞的调谐,这是由于这些细胞中存在一种纤毛束长度和特征性频率之间的反比关系。
其次,调节个体细胞到特定频率的第二种机制在本质上是电的,并且发生在机械-电转导之后,即膜电位。在鱼类、两栖类、爬行类和鸟类的毛细胞膜电位在一个特殊的频率处产生共鸣。包括人类在内的哺乳动物的耳中,电共鸣是否应用于频率调谐仍不明确。电共鸣是通过两栖类实验得到证明的[12]:将脉冲电流刺激海龟毛细胞时,膜电位形成一种按指数规律的、阻尼的、正弦曲线的振动(图9a)。当毛细胞受到具有不变振幅的机械刺激时,它的转导器会在一个较宽的频率范围内产生反应。当毛细胞受到电流刺激后,使膜电位共鸣的特定频率的刺激可以引起最大的感受器电位。在这种动物里,毛细胞对于特定频率的回应的分布一般符合张力映射(或声调谐映射)。例如,对于爬行类和鸟类的听觉感受器官,在基底部的乳突上存在一个沿着器官的,毛细胞与其调谐的频率的连续转变。因此认为,毛细胞基底外侧的突触递质释放是由膜电位控制的。而突触后神经纤维支配的毛细胞是对特定频率刺激反应最强的。
电共鸣的起源是通过使用电压钳技术对孤立的毛细胞进行记录而确定的(图9b)。振动的去极化是由通过电压敏感性Ca2+通道进入毛细胞的电流驱动的;而复极化的现象是由Ca2+敏感性K+通道向外流出的电流引起的。许多因素构成了共振的频率和阻尼:如膜电容,Ca2+、K+通道的数量和动力学性质以及Ca2+扩散的时程等。
