神经振荡:窥探句法解析的时间进程
2025-02-14戚睿盈封叶司富珍


摘" 要" 在语言表型与神经机制之间寻找对应关系, 即所谓的映射问题(the mapping problem), 是当前研究的一大热点。其中, 句法解析的神经机制尤具挑战性, 这涉及到如何在神经活动中识别出对应于句法结构构建的过程, 是人类语言能力之谜破题的关键。近期神经振荡活动的相关研究不仅为句法解析过程中句法加工的心理现实性提供了有力证据, 也展示了利用神经振荡来阐释句法解析过程的神经编码活动的可行性。而理论语言学最简方案有关句法计算的理论模型可以与神经科学中有关神经振荡的实验研究相互印证, 通过此类研究可以窥探句法构建的时间进程。未来研究可集中于四方面:神经振荡与句法加工的更细粒度对齐; 神经振荡的发生机制及其生物学意义; 儿童语言发展过程中神经振荡的变化模式; 语言障碍神经生理基础及其康复应用。
关键词" 神经振荡, 语言理解, 句法解析, 最简方案, 增量转换
分类号 "B842; B845
1" 引言
David Poeppel (2012)提出了语言学与神经科学的映射问题:语言学与神经科学之间本体论结构间具有怎样的联系?这一问题的核心在于, 如
何以一种诠释性的框架将语言加工的不同方面与大脑中的神经活动直接关联起来, 从而揭示自然语言加工的具体神经机制。而要回答这一问题, 最大的挑战源于粒度不匹配问题(granularity mismatch problem), 即理论语言学倾向于以细粒度的视角对语言本体进行描写与研究, 而神经科学研究往往聚焦于较为宏观的、粗粒度的语言行为。这种视角上的差异导致很难在语言表征与神经活动之间建构起对应关系, 这使得语言与人脑相关性的研究容易浮于表面, 难以找到语言现象真实对应的神经机制。映射问题的解决是探究人类语言能力的关键, 因此, 学界开始对句法、语义等语言构建过程中的神经编码活动进行研究(Hale et al., 2022)。
目前, 有越来越多的研究揭示了以低频神经振荡(low frequency neural oscillation)作为句法层级结构构建指标的可行性, 使其成为阐述句法解析相关神经机制的有力候选者。特别是, 这些研究发现了外源性刺激时间与内源性振荡表征时间之间的非同构映射关系, 以及语法知识与语义统计线索、词汇属性的分离, 揭示了人脑专司句法构建的先验语法知识1的存在, 为句法加工的心理现实性提供了证据。此外, 神经振荡的相位相干性(phase coherence)与不同语言层级结构之间的潜在联系也逐渐得到关注, 相关领域涌现出了具有发展潜力的理论模型。
2 "句法解析管窥:序列到层级的增量转换
句法结构的层级性是自然语言区分于其它交际系统的重要特征, 它赋予了人类用有限手段生成无限的语言表达的能力(Chomsky, 1957, 1965)。句法解析(syntactic parsing), 即语言实现从线性语音序列到抽象句法层级结构增量转换的过程, 是语言学及认知科学内广受关注的议题(Traxler, 2014)。在映射问题的框架内, 语言学理论与神经科学研究都对增量转换的句法解析过程有了一定的探索与描述。
2.1" 语言本体理论中的句法解析:句法对象的合并操作
最简方案(the Minimalist Program)是Noam Chomsky在语言学中新近提出的一种理论框架, 旨在寻求语言的结构和解释方式的最简化形式(Chomsky, 1995)。根据最简方案理论, 句法解析实质上是以词汇项(lexical items)为基本原子单位, 通过合并(merge)操作将工作空间(workspace)中的可操作句法对象(accessible syntactic object)构建为二分叉的层级组织集合的过程。以下为合并的数学表达式:(其中, WS =工作空间; P/Q =句法对象; X =附加元素; { } 用于标记短语结构; [ ] 用于标记特征) (Chomsky et al., 2023; Marcolli, Chomsky, amp; Berwick, 2023)。
""""""""""""""""""""""""" (1)
(2)
外合并(直接从词库调用词汇项以生成基础的论元结构等)可表示为:
""""" (3)
内合并(即‘移位’, 从已构建的结构成分内部选取成分移位到新的合并位置, 以核查与话语等信息结构安排所需要的特征)可表示为:
(4)
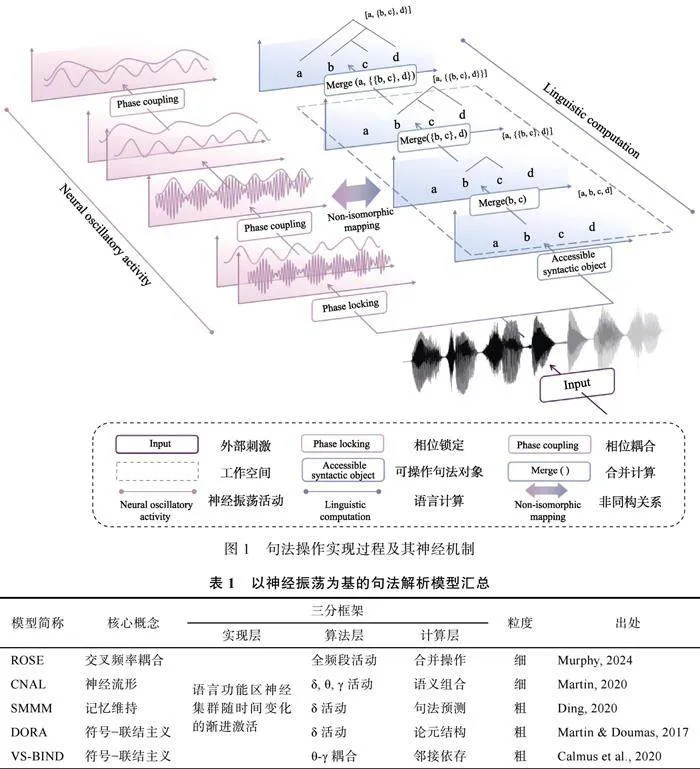
简单来说, 为实现线性序列到层级结构的增量转换, 需要考量以下三个方面:(1)可供合并操作的句法对象的表征如何形成, 即大脑如何收集感觉信息形成词汇表征; (2)工作空间如何动态更新实现对句法对象的存储与预测; (3)合并操作如何将工作空间内的词表征构建为层级结构(Chomsky, 1995)。合并操作最终生成的结果在技术上通常用句法树(syntax tree)来表示, 根据线性对应公理(Linear Correspondence Axiom), 句法树的层级结构映射在线性序列上形成终端节点为标识的语音线性序列, 它以一种可视化的方式展示了句法结构的层级化构建的布局和程序。通过合并操作, 低层级的句法单元被整合在更高层级的句法表征中, 实现了从线性序列到层级结构的增量转换。从心理语言学视角可将其描述为:在线性的语音序列输入的过程中, 人脑整合各类信息在词汇通达(lexical access)后形成词表征(句法对象), 词表征存储入工作记忆(工作空间)后在推断(合并)的过程中被提取与运用, 形成具有垂直层级关系的树形结构(tree notation), 最终实现对句子的理解。
2.2" 神经科学研究中的句法解析:神经振荡与时间结构
研究句法解析的神经机制需要诉诸对人脑活动的观测。如前文所述, 在语言本体理论的框架下, 句法解析是一种将线性序列构建为层级结构的过程。人脑中的神经活动如何表征这一过程, 便是句法解析的神经机制问题。传统上, 学界对于句法解析神经机制的探讨主要集中于事件相关电位(Event Related Potentials, ERPs), 比如与语义违例相关的N400 (Kutas amp; Federmeier, 2000, 2011), 与句法违例相关的P600 (Gouvea et al., 2010, p. 600), 以及与韵律边界识别相关的闭合正漂移(Closure Positive Shift, CPS) (Steinhauer et al., 1999)等。事件相关电位的相关研究已取得重大成效, 作为更敏感的脑电指标, 可以揭露行为表现中无法呈现的心理过程。然而, 通过诱发电位对句法加工过程进行探究具有一定的局限性:事件相关电位的诱发对违例性刺激材料的依赖性较强, 其实验设计在可操作性定义及变量控制上难免失之于主观, 导致其诱发机制解读的重大争议; 且相关研究难以直接提供层级结构加工的时间信息, 难以展现句法解析神经活动的全貌。近年来, 有越来越多的研究发现了神经振荡(neural oscillation)与句法解析之间的潜在联系, 并开始着手研究用神经振荡解释句法解析时不同的心理过程(Bastiaansen et al., 2011), 为句法解析的研究提供另一种可行的路径。
神经振荡是大脑皮层神经集群在时空尺度上表现出的节律性活动。这些活动通常被分为5个主要频段:delta波段(δ: 0.5~4 Hz), theta波段(θ: 4~8 Hz), alpha波段(α: 8~12 Hz), beta波段(β: 12~30 Hz), 低gamma波段(γ: 30~60 Hz)和高gamma波段(60~200 Hz) (Prystauka amp; Lewis, 2019)。根据其频率, 神经振荡又可被分为低频振荡(涵盖δ和θ波段)以及高频振荡(涵盖α、β和γ波段)。作为一种周期性的神经活动, 神经振荡编码信号的一个显著特征便是其时间尺度, 不同频段的神经振荡与不同时间尺度的信息处理相关联。因此, 神经振荡的时空属性使其天然成为编码具有显著时间结构特征信息的候选机制。具体来说, 低频神经振荡因其周期较长, 因此与较大时间窗口的信息交流潜在相关; 相对地, 高频神经振荡由于周期较短, 可能与较小时窗内的信息交流存在联系(张力新 等, 2017)。这一特性表明神经振荡的时间结构与自然语言加工中不同层次结构(词、短语和句子)构建的时间进程存在一定关联, 这意味着神经振荡与不同尺度的句法解析有着潜在联系(胡瑞晨 等, 2019)。此外, 神经振荡可以为大脑系统内部系统的信息处理和传递提供相对精确的时间特征, 这与进行鲁棒性句法解析的需求相符合。
语言学理论已对句法解析的过程做出了丰富的探讨, 而神经科学研究则表明人脑的神经振荡活动与句法解析的时间进程可能存在符合直觉的关联。弥合映射问题之间的鸿沟需要将这两个学科的理论以及研究范式结合起来, 事实上也确实有越来越多的新近研究综合语言学理论与神经科学研究范式, 利用神经振荡相关指标探索句法解析心理过程, 更进一步地为句法层级结构构建过程的心理现实性提供强有力的解释, 并揭示了句法解析潜在的神经编码活动, 推动映射问题的解决。
3" 句法解析探幽:先验语法知识的心理现实性
合并操作形成的语言层级结构是自然语言的核心特征, 在探讨其具体的增量转换过程前, 需要明确语言层级生成过程中句法加工的心理现实性。语言层级结构的生成需要倚赖先验语法知识的表征, 而非单纯是韵律、语义等模块作用的结果。在此视域下, 自然语言的层级结构生成基于先验语法知识[2], 即听者依靠已有语法知识对外来刺激进行自上而下的解析的过程是句法解析的必要条件。从句法解析的角度看, 如果有证据表明建构层级结构的“结构依存原则” (Principle of Structural Dependency)确实存在, 即句子在线加工过程层级结构的表征的句法因素相对独立于韵律、语义、词汇等因素的作用, 那么就可以认为句法加工的心理现实性得到了支持[3]。
低频神经振荡的相关研究对此做出了重要讨论。人们能够根据不同的信息和线索进行有效的语言理解, 这一过程中观测到的低频振荡活动[4]与句法解析任务显示出显著的相关性。然而, 对于低频神经振荡活动具体表征着句子加工中的哪个心理过程, 学界仍存在很大争议。一方面, 一些学者认为低频振荡活动所反映的是内源性(endogenous)神经振荡活动对抽象语言层级结构的表征, 是一个相对独立的、由个体先验知识驱动的过程(Ding, 2023; Giraud, 2020; Haegens, 2020; Kandylaki amp; Kotz, 2020; Klimovich-Gray amp; Molinaro, 2020; Lewis, 2020), 这样的观点被称为层级结构构建假说(Hierarchical Structure Building, HSB)。另一方面, 也有研究认为低频振荡活动源于对外在节律的追踪(Keitel et al., 2018)、统计线索的预测处理(Frank amp; Christiansen, 2018), 以及词汇表征(Frank amp; Yang, 2018)等因素。尽管在实际的言语感知过程中, 低频振荡活动是由不同信息交互作用的共同结果(陈梁杰 等, 2022), 但通过实验设计以及变量控制, 研究者仍然能够将句法结构的表征与其他心理过程进行区分, 从而找到低频振荡活动与句法层级结构的相关性证据。
3.1" 语法知识与韵律信息
由于句法结构与韵律信息在宏观时间尺度上的推进大致相似, 二者引发的神经活动时常容易被混淆。有研究认为低频振荡活动主要是通过追踪语音包络等韵律信息以支持言语感知和信息提取的(Poeppel amp; Assaneo, 2020)。而Meyer等人(2020a, 2020b)则提出不同观点, 认为这种低频活动实质上反应的是对语言结构本身的推断与追踪, 使得外源输入不断与先验语法知识进行内在同步化(intrinsic synchronization)的过程。事实上, 越来越多的证据表明, 仅仅将低频振荡活动归因于对外在节律的追踪并不能充分解释句法表征的成因, 低频振荡与句法计算程序之间的一致性更加显著, 其根本上更可能是对语法知识本身的内在表征。
就韵律−句法界面本身而言, 句法结构与韵律信息存在着非同构关系(non-isomorphic relationship):首先, 语言外化是一种“拼盘式”实现方式(Si, 2016; 司富珍, 2024), 因为虽然“言语产出会使用到内部语言, 但并不能与之等同。关于语言的可计算理论使得我们能够将内化的语言知识系统与访问它的过程区分开来, 而这是一个非常重要的区别” (Chomsky, 2017, p.2)。其次, 自然语言本身的呈现频率并非是固定的。尽管有研究表明自然语音是准节律性的(quasi-rhythmic), 但这并不意味着可以忽略不同自然语音之间的节律差异(Kazanina amp; Tavano, 2023)。另外, 自然语音中韵律边界与句法成分边界的联系相对松散, 二者间并非严格对应(马宝鹏, 庄会彬, 2022), 即对一段自然语音中不同成分的切分与提取难以完全单独依靠声学线索进行。句法范畴本身不由声学边界定义, 而句子本身语义的组合性(compositionality)则需依靠句法成分间的相互关系来构建, 无法仅通过对语音包络的追踪完成对句子的理解(Meyer et al., 2020b)。
正是由于句法结构与外在节律间的非同构关系, 人类在缺乏明确的声学线索或韵律边界的情况下, 仍然能够凭借内在语法知识成功地理解句子。Ding等(2016)通过频率标记范式(frequency tagging paradigm)将不同层级的语言结构(词、词组、句子)以不同的刺激频率呈现, 发现在人工移除句子韵律边界的条件下, 仍能成功分别观测到1、2、4 Hz左右的δ振荡响应; 而相同汉语材料在英语母语被试中却只能观测到4 Hz响应, 说明低频振荡活动实质上是对可理解的语言层级结构的表征, 而非对刺激材料节律的追踪。其他研究也发现, 在语言刺激材料以非周期性频率呈现的情况下, 研究者仍然能观测到表征句法结构加工的1、2、4 Hz的响应(Ding et al., 2016; Jin et al., 2018)。此外, 甚至是在无外在节律刺激的情况下, 如视觉呈现刺激(Henke et al., 2023)、想象言语(Lu et al., 2021)等, 也有与正常句法解析相似的低频神经振荡响应。相反, 在外在节律正常, 而句法信息难以通达的情况下, 如无句法结构的词汇表(word list)条件(Kaufeld et al., 2020)、非母语材料感知(Ding et al., 2016)以及倒放语条件(Gross et"al., 2013; Mai et al., 2016)等情况下, 相应的神经振荡指标都有衰减。
以上实证研究均成功将神经振荡对外在节律刺激的表征与内在语法知识的表征相分离:在外在节律无法提供充足声学线索的情况下, 研究者仍然能观测到与句子层级相关的低频振荡活动; 而在外在节律线索丰富但缺乏语法知识通达的情况下, 则不一定能观察到相应的振荡响应。这表明了低频神经振荡所反映的是内在语法知识的推断过程而非简单地、自下而上地对外在节律的感知。
3.2" 句法规则与语义统计线索
广泛的研究表明, 人类可以通过语义统计线索来预测言语感知的内容(Dikker et al., 2010; Poeppel et al., 2008)。例如, 词频(word frequency)显著影响单个单词的识别时间(Gardner et al., 1987)。词间转移概率(transitional probability)与单词在句中的阅读时间也有很强的相关性(Smith amp; Levy, 2013)。在噪声环境下, 词与词之间转移概率较高的句子比转移概率较低的句子识别效果更好(Miller et al., 1951)。基于概率线索在言语加工中的作用, 有学者认为句法解析的过程实质上是基于语义分析及统计概率预测的产物。相比于句法层级结构, 线性序列本身传递的统计信息对语言理解来说更重要。因此, 低频神经振荡的响应模式在仅针对线性序列关系的统计模型中也可实现(Frank amp; Christiansen, 2018)。然而, 尽管统计概率能辅助和补偿语言理解, 本文认为基于规则的句法层级构建是语言理解的主要驱动力, 语义统计概率并不足以完全说明语言理解过程。
自然语言句法本身的层级结构性无法仅通过统计概率解释。统计概率本质上是基于简单线性序列的信息, 而自然语言中的词组之间并非简单的邻接依存关系(adjacent dependency), 而是存在复杂的长距离依存结构(long-distance dependency) (Berwick et al., 2013)。这表明句法解析的过程需要句法规则来进行层级结构的构建, 仅依靠线性概率无法生成具有递归性质的自然语言(Ding et"al., 2017)。事实上, 有研究通过将句法组块与具体词汇语义分离, 发现低频神经振荡更可能是句法层级构筑的结果, 而非对词汇语义及概率分析的产物(Jin et al., 2020; Lu et al., 2023)。
此外, 研究表明人类可以在低程度依赖转移概率的情况下实现对语言的理解, 但却无法在不依赖句法层级结构的情况下理解句子。Ding等(2016)在低频振荡的实验中通过构建马尔可夫句集(Markovian Sentence Set, MSS)对实验句的转移概率的控制, 最终发现转移概率的高低对δ振荡响应强度的调控作用并不显著。此外, 句法结构本身也可对与统计预测产生调控作用。使用概率线索进行预测在很多时候需要依赖句法的结构层级。Slaats等(2023)通过对前人研究的MEG数据集进行分析, 发现语境驱动的概率线索的影响仅在句子条件下显著, 而在词汇表条件下则不然。这表明在言语感知过程中的低频振荡的响应是由句法结构与语义共同调制的, 而非仅仅受到词汇语义间转移概率差异的影响。概率线索的运用本身也需要以句法层级结构为条件。相似的, Rafferty等(2023)的研究通过最小短语范式(minimal phrase paradigm)以及Jabberwocky范式, 发现短语层的低频神经振荡响应(0.5Hz)主要与句法组合有关而并不与具体的词汇语义性质相关联。除了低频振荡的相关研究之外, 其他心理语言学实验也区分了句法与语义加工的神经机制(Brennan amp; Hale, 2019; Pallier et al., 2011), 支持了两者的相对独立性。
3.3" 句法层级与词汇属性
除了排除韵律、语义模块的相关性之外, 针对低频振荡在句法解析过程中的频段响应模式, Frank和Yang (2018)提出词汇表征说(lexical representation account), 主张实验中观测到的频段响应并非是与句法层级结构有关, 而是与刺激材料中特定句法范畴的词汇呈现频率相关。也就是说, 1 Hz低频神经振荡响应并非是对句子层级本身的响应, 而是对着句中某个呈现频率为1 Hz的词汇属性的反应。如“老牛吃草”这一句子中, 动词(“吃”)的出现次数为1次, 名词(“牛” “草”)的出现次数为2次, 单个字表征(“老” “牛” “吃” “草”)的出现次数为4次, 分别对应着1、2、4 Hz的呈现频率。这类理论虽将低频振荡活动的响应归因为词汇句法范畴, 但其并不承认句法结构层级在句法解析过程中的作用。
对此, Jin (2020)通过组块任务证明低频神经振荡响应的产生是基于句法规则的组块过程产生的, 并非是词汇属性组合的副产品。还有研究利用反转词汇(reversed phrase), 保留基线句中原有的词汇属性的频率与分布, 而去除原有的句法层级结构信息, 最终在相应条件下未观测到相应的1 Hz神经指标(Lo et al., 2022)。这些结果与上述词汇表征说相悖, 证实了层级结构对于低频神经振荡响应的必要性。
综上, 本文认为, 低频神经振荡活动所反映的更可能是专司句法结构的神经机制, 其本身表征的语言层级结构计算的“结构依存原则”的确存在, 并不是语音加工、概率预测等活动过程的副产品。毋庸讳言, 在实际言语感知的过程中, 句法解析的过程是内源性振荡与外在节律、概率预测、词汇语义等共同调控的产物, 在某些时刻低频振荡相关活动所对应的神经回路也很难从解剖结构上严格区分(Meyer et al., 2020a), 但这并不意味着这种不同表征的分离是没有价值的。相反, 这能够帮助我们分离语言层级结构表征中的不同模块, 为句法解析过程中句法加工的心理现实性提供解释。
4" 句法解析溯因:具体神经编码过程
正如前文所述, 研究者可以通过神经振荡分离句法解析中的多重信息表征。其中大多数相关实验都是通过观测不同神经振荡的响应强度(power)或幅值(amplitude)以分离不同模块线索表征。但要进一步解释句法解析的神经编码过程, 将神经振荡活动与语言层级结构相对其, 就必须对神经振荡更具有时间属性的物理性质有更深入的了解。在句法加工心理现实性存在的前提下, 句法解析过程中不同信息的整合如何通过神经振荡之间的协调来实现?
近来, 有越来越多的理论模型及实证开始利用神经振荡的相位相干性(phase coherence)探究句法解析的相关过程(Bai et al., 2022; Brennan amp; Martin, 2019; Calmus et al., 2020; Ding, 2020; Flanagan amp; Goswami, 2018; Ghitza, 2011; Martin, 2020; Martin amp; Doumas, 2017; Murphy, 2024)。这使得神经振荡的信号变换与语言层级结构构建的时间进程相对齐, 是对句法解析神经编码活动的更深入探讨。研究表明, 神经集群对句法层级结构的表征可能是通过同一频段或不同频段振荡的相位同步与去同步化过程来完成的。也就是说, 不同神经集群会随着加工时间变化逐步激活或抑制, 并产生不同相位属性的振荡信号, 对不同时间尺度的感觉信息进行存储、维持与提取, 最终实现从结构层级构建的动态编码。
4.1" 相位相干性
神经振荡空间分布的显著特性主要依赖其相位(phase)属性。相位, 指的是振荡信号在周期性波形中的位置。相位相干性, 指的是不同神经元之间振荡活动的相位同步(phase synchronization)程度, 是衡量神经元之间协调活动的重要指标。相位同步有不同的表现形式, 如同一频段不同脑区间的相位同步(Fries, 2005), 不同频段振荡的间相位相关的交叉节律耦合(Cross-Frequency Coupling, CFC)等。在句法解析的过程中, 同一频段内与不同频段间的神经振荡会表现出不同程度的相位相干性。因此, 相位相干性被认为反映了语言不同层级的表征在不同时间尺度上整合信息的过程(Giraud amp; Poeppel, 2012)。
具体来说, 有研究利用CFC等相关指标发现, 在神经振荡对语言结构进行追踪的过程中, 会系统性地展现出频段内与频段间的相位同步现象, 包括低频耦合(如δ-θ耦合), 以及低频高频耦合(如θ-γ耦合)等形式(Bai et al., 2022; Brennan amp; Martin, 2019)。这种耦合过程中, 高频神经振荡携带的低层级感官信息与低频神经振荡表征的高层级语言结构进行匹配, 是实现语言线性序列合并为层级结构的关键一步。尽管目前的研究暂未发现特定语言层级与显示出同步化的频率带之间严格的对齐关系, 但仍然可以基于现有的成果对句法解析的过程做出更进一步的假设。如图1所示, 神经振荡的相位耦合信号模式与句法树符号的横向表示和纵向结构相匹配(参见Marcolli, et al., 2023a, 2023b), 且与神经振荡的相位属性存在着非同构映射(non-isomorphic mapping)关系, 即神经振荡活动信号与句法结构的对应可能是非线性的, 但两者可经过一定的数学变换进行转换(参见Murphy, 2024; Coopmans et al., 2023; Kazanina amp; Tavano, 2023)。
4.2" 理论模型
目前已有部分理论模型以相位相干性为基, 对神经振荡编码句法解析提供了不同视域下的细致描述(见表"1)。其中, ROSE (Representations, Operations, Structures, Encoding)模型围绕交叉频率耦合等神经信号变换模式, 侧重于在句法操作与神经活动之间建立联系(Murphy, 2024); CNAL (Compositional Neural Architecture for Language)模型以神经流形(neural manifolds)为基础, 为语义组合性实现过程中语音、词汇、句法、语义等不同模态间的相互转化提供了解释性机制; SMMM (Structure-based Memory Maintenance Model)作为经典的心理学模型, 其主要特点在于清晰地具象化句法长短时工作记忆在维持结构表征时的作用(Ding, 2020); DORA (Discovery of Relations by Analogy)作为符号−联结主义模型, 提出了论元结构的一种可能的生成与预测机制(Martin amp; Doumas, 2017); VS-BIND (Vector- symbolic Sequencing of Binding INstantiating Dependencies)使用向量的圆周卷积运算(circular convolution, 运算符“⊗”)模拟θ-γ振荡耦合, 以此来表征句法依存关系(Calmus et al., 2020)。这些模型从不同维度对句法解析及其神经编码活动提出了假设, 在其适用范围内均具有较强的解释力, 并且得到了一定的实证数据支持。然而, 由于学科视角和研究范式不同, 这些模型不具有直接可比性。为更深入全面地介绍各个模型的特征与异同, 本文尝试在David Marr提出的三分框架内对它们进行系统性的评述。
David Marr (1982)[5]关于人脑信息处理机制的三层次理论为相关认知科学研究提供了重要参考框架, 这三个层次分别是:计算层(computational level)、算法层(algorithmic level)及实现层(implementational level)。在本文的语境中, 这三个层次分别关注:(1)语言的基本计算功能, 如语法规则的使用; (2)神经振荡活动, 探讨其在语言处理中的时间过程和调控; (3)底层的生物学机制, 如基因、蛋白质、神经元和脑回路的运作。依照这种三分框架, 本文将立足于映射问题视角, 从计算层、算法层以及实现层的角度对模型的异同点进行梳理, 以此讨论它们对句法解析过程的阐述程度(见图2)。
在这统一的理论框架下, 上述模型具有三个方面的共性。首先, 从实现层来说, 这些模型在解释句法解析的底层生物学机制上是相似的, 它们都以神经集群在句法加工中逐步的、渐进的激活为前提解释表征句法结构的神经振荡活动。也就是说, 随着外界刺激的不断输入, 不同神经集群会在语言层级结构形成的过程中次第激活, 产生节律性的神经信号, 实现对语言层级的动态表征。值得一提的是, 目前的这5个模型暂未涉及更深入的神经生理与动力学层面的探讨。其次, 从算法层来说, 这些模型都假设语言层级结构表征的关键在于神经信号时间结构的耦合, 通过其相位相干性来说明句法解析的过程。最后, 从计算层来说, 尽管这5个模型基于不同的语言学理论, 对不同结构类型进行了解释, 但它们都肯定了语言表型当中层级结构的存在, 并围绕这一点展开讨论。因此, 这5个模型在总体上都揭示了神经振荡作为句法结构构建发生机制的可行性, 认为神经振荡是句法解析的潜在神经机制。
同时, 上述模型也存在三个方面的差异。最主要的差异首先体现在算法层。这些模型对于不同频段神经振荡与不同语言层级结构的具体联系有不同解读:ROSE将全频段的神经活动与最简方案理论中的句法计算过程进行映射, 假设γ振荡活动表征句法对象(也就是词汇)的形成, θ-γ耦合表征词汇调用过程, δ-θ耦合表征层级结构的合并, α振荡与抑制活动有关, β振荡与预测活动有关; CNAL与ROSE相似, 唯一不同的是未讨论α与β波段在此过程中的作用。SMMM与DORA主要强调了δ振荡在语言层级实现中的核心地位, 而对其它频段的讨论较少; VS-BIND则仅基于θ-γ耦合对句法加工的过程进行解释。
根据目前的实证研究结果, δ频段的相关研究已成功排除了韵律、语义、词汇等因素的影响, 说明δ振荡与句法层级结构构筑之间更相关(Ding et"al., 2016), 而θ振荡更可能与刺激材料的语音包络共变(Chalas et al., 2023), γ振荡则更可能与刺激材料的声学特征结构相关(Xu, Zhao et al., 2023)。这意味着θ-γ耦合可能表征词汇的形成, 这一过程发生在语言层级结构构建之前。从这一角度来说, VS-BIND的可信度弱于其它模型, 而其它四个模型的神经机制均有实证研究结果的支撑。对于α与β振荡, 也有认知功能相关的研究支持其分别与抑制与计时(Klimesch, 2012)及预测活动(Abbasi amp; Gross, 2020)相关。因此, ROSE对句法解析过程的阐述更加全面。此外, 并非所有模型都对这些神经信号的活动模式进行了数学层面的模拟。CNAL利用神经流形这一数据模型, 假设神经集群的信号模式可被降维成某一高维中的内嵌流形, 并以其中子空间(subspace)的坐标系变换(coordination transformation)来解释外界刺激在不同模块之间如何转化表征; VS-BIND则以圆周卷积运算来模拟θ-γ振荡耦合, 以此解释句法成分之间的邻接与非邻接关系; 而ROSE、DORA、SMMM则未利用具体的数学工具对神经振荡表征句法层级的信号模式进行解读。
除算法层方面的差异之外, 这些模型在计算层聚焦的语言本体结构也有所不同。具体而言, ROSE与CNAL较为系统地涉及到了句法解析从原子特征到层级结构构筑的全方面, 对于外界输入在语音、词汇、句法、语义模块的转化都有自己的诠释, 属于更细粒度的模型; 而SMMM、DORA、VS-BIND则侧重于解释句法层级结构形成的过程本身, 探讨了抽象的句法记忆装置是如何实现对层级结构的维持和预测的, 属于相对粗粒度模型, 但更直观地展现了句法层级构筑的心理过程。
从模型本身所属的类别以及核心内容来看, 这些模型的底层运行机制涉及不同学科的研究范式, 并存在较大差别。ROSE基于语言学理论框架, 系统性地将各个频段神经振荡及其之间的活动对应至句法解析过程中的某个步骤, 重在体现语言表型与神经活动之间的关系; CNAL引入了前沿的神经流形概念, 属于计算神经科学研究范式; SMMM则为经典的心理机制模型, 其核心在于具象化了句法单元的维持与表征, 并凸显了句法长短时记忆装置在其中的作用; DORA与VS-BIND均为符号−联结主义(symbolic-connectism)模型, 利用神经网络描述句法解析过程。因此, 这些模型在一定程度上具有不可比性, 它们之间的差异并不体现在其是否相悖, 而体现在从不同的维度对句法解析的过程进行解释。
综上, ROSE和CNAL以细粒度的视角完备地将不同频段的神经振荡活动与句法解析过程相对应, 并且有较多实证研究结果支撑, 能更好地解决映射问题, 具备良好发展潜能。其它三个模型虽然粒度不及前二者, 但在具象化句法层级构建的行为过程方面仍然具备较强解释力, 为后续的研究提供了参考。
综上所述, 相位相干性与句法层级结构的潜在联系已经逐渐得到关注。尽管目前缺乏将神经振荡与句法解析过程相联系的严谨的数学证明及动力学计算模型, 但神经振荡相位性质显著的时间分布特征的确能够为句法解析的神经编码活动提供一种潜在的合理解释。
5" 总结与展望
句法层级结构是自然语言区别于其他交际系统的本质属性, 句法解析的过程需要着重解释语音线性序列与句法层级结构的增量转换。在这一过程中, 尽管韵律、统计概率等线索都会参与加工, 但这些线索对于句法解析来说并不充分, 鲁棒性句法结构表征的形成仍需要倚赖先验语法知识。回顾现有的神经振荡研究, 相关领域已累积丰富的证据支持抽象语言结构的内在句法表征, 并展现出利用神经振荡解释句法解析神经编码过程的巨大潜力。神经振荡已经成为探索语言与神经基元映射问题的重要切入点。在未来研究中, 以下几个方面值得关注。
第一, 探究句法解析的因果性神经编码活动。本文对神经振荡以及语言结构关系的阐述属于一种猜想, 如若要进一步证明两者之间更细粒度的映射关系, 还需进一步探究。具体来说, 首先, 目前研究主要以神经振荡具体频段的幅值和强度作为观测指标, 而缺乏对神经振荡相位属性与语言层级结构之间关系的探究。因此, 大部分的研究结果可能停留于对句法知识的存在性探讨, 而不能直接证明不同频段的神经振荡如何通过信号变换来表征语言层级结构, 实现增量转换。未来研究应更深入地探索神经振荡相位属性与句法解析的具体心理过程之间的联系, 利用相位耦合相关指标(如CFC), 以及可充分分离不同语言层级结构的实验范式(如频率标记范式)进行研究。此外, 目前研究主要依赖传统心理学实验中控制变量的手段来进行研究, 通过直接观察不同条件对比的结果来回答研究问题, 得出的相关结论可能受到语言表层形式的制约。未来研究可将句法结构的特征(如句法树深度, 节点等)编码为抽象的数据结构来进行建模与验证(参考Gwilliams et al., 2024), 以越过表层的语言实现形式, 进一步解释神经振荡与抽象句法结构之间的潜在联系。最后, 本文回顾的研究在实验设计与统计学方法上都只涉及相关性分析而暂未涉及因果推断。在日后实验数据储备允许的情况下, 可以通过更多的数学与统计方法对其进行分析, 推进对句法解析的更细粒度的因果性的解释, 在两者的非严格同构结构间找到映射关系, 并将其与神经振荡的精细活动对齐。目前的语言学理论已经试图用抽象代数将句法计算层的活动进行细致的描绘(Marcolli, Chomksy, amp; Berwick, 2023); 已有的句法解析模型中也有试图将句法层级构建的数学模型与其具体神经生理性相匹配(Kaushik amp; Martin, 2022)。未来研究可以更进一步地在这些理论框架的指导下探索句法解析的神经机制。
第二, 厘清神经振荡的发生机制及其生物学意义。神经振荡活动的实现层机制尚存在争议:首先, 神经振荡与诱发响应存在很大的区分困难。神经振荡本身可能涌现于无数个诱发响应的组合(Ding, 2022), 而诱发响应本身也可能产生于振荡活动的平均调制(Meyer et al., 2020a)。其次, 不同成像技术观测的神经振荡指标存在本质差异, 且由于EEG与MEG空间分辨率的缺陷, 目前研究也难以将振荡活动与宏观或微观的神经结构进行对应[6]。神经生理相关研究表明, 神经振荡信号的周期性可能与神经集群的激活抑制循环机制有关, 本文着重于讨论其作为句法构建的发生原理与阐述机制的可能, 因而没有对这些神经活动本身进行梳理。未来如若要更进一步地探究句法解析过程, 势必需要探究这些神经活动之间的因果, 将神经振荡的空间分布与其时间结构进行联系, 将其与神经集群兴奋传导的具体模式相对应, 以明确其具体神经生理基础, 或者通过基因学研究找到与语言层级结构相关的基因表达, 弥合句法解析算法层以及实现层之间的鸿沟。
第三, 关注语言发展过程中神经振荡模式的变化。从语言习得的角度来说, 儿童句法结构的习得体现为从对外在节律的倚赖到内在语法知识的形成的转变。婴幼儿语言习得早期具有知觉窄化(perceptual narrowing) (Kuhl, 2004)以及发展性权重转移(Seidl, 2007)的现象。这可能是因为随着相关大脑皮层发育, 婴幼儿对语言输入的识别与区分逐渐由依赖声学特征向依靠语法知识转变(Meyer et al., 2020b)。而目前相关的假设与研究都主要从对外在节律的跟踪出发考察神经振荡在儿童语言习得中的作用(Attaheri et al., 2022; Nallet amp; Gervain, 2021), 后续的相关研究可以利用频率标记范式等研究方法进一步分离语法知识与韵律信息以考察两者在语言习得过程中的不同作用。
第四, 明确不同语言障碍群体的神经生理缺陷。从语言蚀失的角度而言, 老年人语言能力的衰退在神经振荡活动中可体现为低层级语言结构(4Hz)振荡响应的增强和高层级语言结构(1、2 Hz)振荡响应的减弱(Xu, Qin et al., 2023), 这体现了抽象语法知识衰退对于低频神经振荡活动的影响。后续研究可以低频神经振荡为切入点探究老年人群语言蚀失的具体神经表现。从特殊人群角度出发, 前人研究发现, 与健康人群相比, 语言障碍人群的神经振荡活动会出现一定的变异(Fridriksson et al., 2015; Meyer et al., 2021; Peter et"al., 2023), 这意味着其内在的语法知识表征可能存在一定的缺陷。尤其是对于神经发育障碍(Neural Developmental Disorder, NDD)人群, 其语言障碍发生机制尚无明确的病理学定义, 往往难以通过高空间分辨率脑成像技术确定其脑损伤病灶(Abbott amp; Love, 2023), 可以考虑利用神经振荡指标来考察其语言行为缺陷。已有理论已试图从内源性振荡活动的角度对发展性失读症(Developmental Dyslexia, DD)语言障碍的神经缺陷进行解释(Goswami, 2011), 未来研究可拓展到更难以界定的发展性语言障碍(Developmental Language Disorder, DLD)群体。对语言障碍群体的研究, 不仅可以为句法解析过程中的神经振荡机制提供病理性证据, 也可进一步克服部分语言障碍群体研究的异质性问题, 为该类疾病的早期预测与临床干预工作提供更为统一的病理学标准。
参考文献
陈梁杰, 刘雷, 葛钟书, 杨晓东, 李量. (2022). 节律在听觉言语理解中的作用. 心理科学进展, 30(8), 1818− 1831.
胡瑞晨, 袁佩君, 蒋毅, 王莹. (2019). 时间结构信息在人类知觉中的作用及其脑机制. 生理学报, 71(1), 105−116.
姜孟. (2009). 句法自治: 争鸣与证据. 外国语文, 25(3), 79−86.
马宝鹏, 庄会彬. (2022). 二十年来韵律-句法接口研究的回顾与启示. 外国语, 45(1), 56−66.
司富珍. (2024). 语言与人脑科学研究中的“伽利略谜题”.外国语, 47(2), 2−9.
杨烈祥. (2012). 唯递归论的跨语言比较述评. 外语教学与研究, 44(1), 54−64+158.
张力新, 王发颀, 王玲, 杨佳佳, 万柏坤. (2017). 认知功能研究中神经振荡交叉节律耦合应用研究进展. 生理学报, 69(6), 805−816.
Abbasi, O., amp; Gross, J. (2020). Beta-band oscillations play an essential role in motor-auditory interactions. Human Brain Mapping, 41(3), 656−665.
Abbott, N., amp; Love, T. (2023). Bridging the divide: Brain and behavior in developmental language disorder. Brain Sciences, 13(11), 1606.
Attaheri, A., Choisdealbha, Á. N., Di Liberto, G. M., Rocha, S., Brusini, P., Mead, N., ... Goswami, U. (2022). Delta- and theta-band cortical tracking and phase-amplitude coupling to sung speech by infants. NeuroImage, 247, 118698.
Bai, F., Meyer, A. S., amp; Martin, A. E. (2022). Neural dynamics differentially encode phrases and sentences during spoken language comprehension. PLoS Biology, 20(7), e3001713. https://doi.org/10.1371/journal.pbio.3001713
Bastiaansen, M., Mazaheri, A., amp; Jensen, O. (2011). Beyond ERPs: Oscillatory neuronal dynamics. In E. S. Kappenman amp; S. J. Luck (Eds.), The Oxford handbook of event- related potential components. Oxford University Press.
Berwick, R. C., Friederici, A. D., Chomsky, N., amp; Bolhuis, J. J. (2013). Evolution, brain, and the nature of language. Trends in Cognitive Sciences, 17(2), 89−98.
Brennan, J. R., amp; Hale, J. T. (2019). Hierarchical structure guides rapid linguistic predictions during naturalistic listening. PloS One, 14(1), e0207741. https://doi.org/10. 1371/journal.pone.0207741
Brennan, J. R., amp; Martin, A. E. (2019). Phase synchronization varies systematically with linguistic structure composition. Philosophical Transactions of the Royal Society B: Biological Sciences, 375(1791), 20190305.
Calmus, R., Wilson, B., Kikuchi, Y., amp; Petkov, C. I. (2020). Structured sequence processing and combinatorial binding: Neurobiologically and computationally informed hypotheses. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, 375(1791), 20190304.
Chalas, N., Daube, C., Kluger, D. S., Abbasi, O., Nitsch, R., amp; Gross, J. (2023). Speech onsets and sustained speech contribute differentially to delta and theta speech tracking in auditory cortex. Cerebral Cortex, 33(10), 6273−6281.
Chomsky, N. (1957). Syntactic structures. De Gruyter Mouton.
Chomsky, N. (1965). Aspects of the theory of syntax. The MIT Press.
Chomsky, N. (1995). The minimalist program. The MIT Press.
Chomsky, N. (2017). The Galilean challenge: Architecture and evolution of language. Journal of Physics: Conference Series, 880, 12−15.
Chomsky, N., amp; Moro, A. (2022). The Secrets of words. The MIT Press.
Chomsky, N., Seely, T. D., Berwick, R. C., Fong, S., Huybregts, M. A. C., Kitahara, H., McInnerney, A., amp; Sugimoto, Y. (2023). Merge and the strong minimalist thesis. Cambridge University Press.
Coopmans, C. W., Mai, A., Slaats, S., Weissbart, H., amp; Martin, A. E. (2023). What oscillations can do for syntax depends on your theory of structure building. Nature Reviews Neuroscience, 24(11), 723−723.
Dikker, S., Rabagliati, H., Farmer, T. A., amp; Pylkkänen, L. (2010). Early occipital sensitivity to syntactic category is based on form typicality. Psychological Science, 21(5), 629−634.
Ding, N. (2020). A structure-based memory maintenance model for neural tracking of linguistic structures. arXiv. https://doi.org/10.48550/arXiv.2002.11870
Ding, N. (2022). The neural correlates of linguistic structure building: Comments on Kazanina amp; Tavano (2022). arXiv. https://doi.org/10.48550/arXiv.2212.04219
Ding, N. (2023). Low-frequency neural parsing of hierarchical linguistic structures. Nature Reviews Neuroscience, 24(12), 792−792.
Ding, N., Melloni, L., Tian, X., amp; Poeppel, D. (2017). Rule-based and word-level statistics-based processing of language: Insights from neuroscience. Language, Cognition and Neuroscience, 32(5), 570−575.
Ding, N., Melloni, L., Zhang, H., Tian, X., amp; Poeppel, D. (2016). Cortical tracking of hierarchical linguistic structures in connected speech. Nature Neuroscience, 19(1), 158−164.
Flanagan, S., amp; Goswami, U. (2018). The role of phase synchronisation between low frequency amplitude modulations in child phonology and morphology speech tasks. The Journal of the Acoustical Society of America, 143(3), 1366−1375.
Frank, S. L., amp; Christiansen, M. H. (2018). Hierarchical and sequential processing of language: A response to: Ding, Melloni, Tian, and Poeppel (2017). Rule-based and word-level statistics-based processing of language: Insights from neuroscience. Language, Cognition and Neuroscience, 33(9), 1213−1218.
Frank, S. L., amp; Yang, J. (2018). Lexical representation explains cortical entrainment during speech comprehension. PLoS One, 13(5), e0197304. https://doi.org/10.1371/ journal.pone.0197304
Fridriksson, J., Basilakos, A., Hickok, G., Bonilha, L., amp; Rorden, C. (2015). Speech entrainment compensates for Broca’s area damage. Cortex, 69, 68−75.
Fries, P. (2005). A mechanism for cognitive dynamics: Neuronal communication through neuronal coherence. Trends in Cognitive Sciences, 9(10), 474−480.
Gardner, M. K., Rothkopf, E. Z., Lapan, R., amp; Lafferty, T. (1987). The word frequency effect in lexical decision: Finding a frequency-based component. Memory amp; Cognition, 15(1), 24−28.
Ghitza, O. (2011). Linking speech perception and neurophysiology: Speech decoding guided by cascaded oscillators locked to the input rhythm. Frontiers in Psychology, 2, 130.
Giraud, A.-L. (2020). Oscillations for all ¯_(ツ)_/¯? A commentary on Meyer, Sun amp; Martin (2020). Language, Cognition and Neuroscience, 35(9), 1106−1113.
Giraud, A.-L., amp; Poeppel, D. (2012). Cortical oscillations and speech processing: Emerging computational principles and operations. Nature Neuroscience, 15(4), 511−517.
Goswami, U. (2011). A temporal sampling framework for developmental dyslexia. Trends in Cognitive Sciences, 15(1), 3−10.
Gouvea, A., Phillips, C., Kazanina, N., amp; Poeppel, D. (2010). The linguistic processes underlying the P600. Language and Cognitive Processes, 25(2), 149−188.
Gross, J., Hoogenboom, N., Thut, G., Schyns, P., Panzeri, S., Belin, P., amp; Garrod, S. (2013). Speech rhythms and multiplexed oscillatory sensory coding in the human brain. PLoS Biology, 11(12), e1001752. https://doi.org/10.1371/ journal.pbio.1001752
Gwilliams, L., Marantz, A., Poeppel, D., amp; King, J. R. (2024). Hierarchical dynamic coding coordinates speech comprehension in the brain. bioRxiv: The preprint server for biology, 2024.04.19.590280.
Haegens, S. (2020). Entrainment revisited: A commentary on Meyer, Sun, and Martin (2020). Language, Cognition and Neuroscience, 35(9), 1119−1123.
Haegens, S., amp; Zion Golumbic, E. (2018). Rhythmic facilitation of sensory processing: A critical review. Neuroscience amp; Biobehavioral Reviews, 86, 150−165.
Hale, J. T., Campanelli, L., Li, J., Bhattasali, S., Pallier, C., amp; Brennan, J. R. (2022). Neurocomputational models of language processing. Annual Review of Linguistics, 8(1), 427−446.
Henke, L., Lewis, A. G., amp; Meyer, L. (2023). Fast and slow rhythms of naturalistic reading revealed by combined eye-tracking and electroencephalography. Journal of Neuroscience, 43(24), 4461−4469.
Jin, P., Lu, Y., amp; Ding, N. (2020). Low-frequency neural activity reflects rule-based chunking during speech listening. eLife, 9, e55613. https://doi.org/10.7554/eLife. 55613
Jin, P., Zou, J., Zhou, T., amp; Ding, N. (2018). Eye activity tracks task-relevant structures during speech and auditory sequence perception. Nature Communications, 9(1), 5374.
Kandylaki, K. D., amp; Kotz, S. A. (2020). Distinct cortical rhythms in speech and language processing and some more: A commentary on Meyer, Sun, amp; Martin (2019). Language, Cognition and Neuroscience, 35(9), 1124− 1128.
Kaufeld, G., Bosker, H. R., Ten Oever, S., Alday, P. M., Meyer, A. S., amp; Martin, A. E. (2020). Linguistic structure and meaning organize neural oscillations into a content- specific hierarchy. Journal of Neuroscience, 40(49), 9467−9475.
Kaushik, K. R., amp; Martin, A. E. (2022). A mathematical neural process model of language comprehension, from syllable to sentence. PsyArXiv. https://doi.org/10.31234/ osf.io/xs5kr.
Kazanina, N., amp; Tavano, A. (2023). What neural oscillations can and cannot do for syntactic structure building. Nature Reviews Neuroscience, 24(2), 113−128.
Keitel, A., Gross, J., amp; Kayser, C. (2018). Perceptually relevant speech tracking in auditory and motor cortex reflects distinct linguistic features. PLoS Biology, 16(3), e2004473. https://doi.org/10.1371/journal.pbio.2004473
Klimesch, W. (2012). α-band oscillations, attention, and controlled access to stored information. Trends in Cognitive Sciences, 16(12), 606−617.
Klimovich-Gray, A., amp; Molinaro, N. (2020). Synchronising internal and external information: A commentary on Meyer, Sun amp; Martin (2020). Language, Cognition and Neuroscience, 35(9), 1129−1132.
Kuhl, P. K. (2004). Early language acquisition: Cracking the speech code. Nature Reviews. Neuroscience, 5(11), 831−843.
Kutas, M., amp; Federmeier, K. D. (2000). Electrophysiology reveals semantic memory use in language comprehension. Trends in Cognitive Sciences, 4(12), 463−470.
Kutas, M., amp; Federmeier, K. D. (2011). Thirty years and counting: Finding meaning in the N400 component of the event-related brain potential (ERP). Annual Review of Psychology, 62, 621−647.
Lewis, A. G. (2020). Balancing exogenous and endogenous cortical rhythms for speech and language requires a lot of entraining: A commentary on Meyer, Sun amp; Martin (2020). Language, Cognition and Neuroscience, 35(9), 1133−1137.
Lo, C.-W., Tung, T.-Y., Ke, A. H., amp; Brennan, J. R. (2022). Hierarchy, not lexical regularity, modulates low-frequency neural synchrony during language comprehension. Neurobiology of Language, 3(4), 538−555.
Lu, L., Sheng, J., Liu, Z., amp; Gao, J.-H. (2021). Neural representations of imagined speech revealed by frequency- tagged magnetoencephalography responses. NeuroImage, 229, 117724.
Lu, Y., Jin, P., Ding, N., amp; Tian, X. (2023). Delta-band neural tracking primarily reflects rule-based chunking instead of semantic relatedness between words. Cerebral Cortex, 33(8), 4448−4458.
Mai, G., Minett, J. W., amp; Wang, W. S.-Y. (2016). Delta, theta, beta, and gamma brain oscillations index levels of auditory sentence processing. NeuroImage, 133, 516−528.
Marcolli, M., Berwick, R. C., amp; Chomsky, N. (2023a). Old and New Minimalism: A Hopf algebra comparison. arXiv. https://doi.org/10.48550/arXiv.2306.10270
Marcolli, M., Berwick, R. C., amp; Chomsky, N. (2023b). Syntax-semantics interface: An algebraic model. arXiv. https://doi.org/10.48550/arXiv.2311.06189
Marcolli, M., Chomsky, N., amp; Berwick, R. (2023). Mathematical structure of syntactic merge. arXiv. https://doi.org/10.48550/arXiv.2305.18278
Marr, D. (1982). Vision: A computational investigation into the human representation and processing of visual information. W.H. Freeman.
Martin, A. E. (2020). A compositional neural architecture for language. Journal of Cognitive Neuroscience, 32(8), 1407−1427.
Martin, A. E., amp; Doumas, L. A. (2017). A mechanism for the cortical computation of hierarchical linguistic structure. PLoS Biology, 15(3), e2000663. https://doi.org/10.1371/ journal.pbio.2000663
Martins, P. T., amp; Boeckx, C. (2019). Language evolution and complexity considerations: The no half-Merge fallacy. PLoS Biology, 17(11), e3000389. https://doi.org/10.1371/ journal.pbio.3000389
McClamrock, R. (1991). Marr's three levels: A re-evaluation. Minds and Machines, 1, 185−196.
Meyer, L., Lakatos, P., amp; He, Y. (2021). Language dysfunction in Schizophrenia: Assessing neural tracking to characterize the underlying disorder (s)? Frontiers in Neuroscience, 15, 640502.
Meyer, L., Sun, Y., amp; Martin, A. E. (2020a). “Entraining” to speech, generating language? Language, Cognition and Neuroscience, 35(9), 1138−1148.
Meyer, L., Sun, Y., amp; Martin, A. E. (2020b). Synchronous, but not entrained: Exogenous and endogenous cortical rhythms of speech and language processing. Language, Cognition and Neuroscience, 35(9), 1089−1099.
Miller, G. A., Heise, G. A., amp; Lichten, W. (1951). The intelligibility of speech as a function of the context of the test materials. Journal of Experimental Psychology, 41(5), 329−335.
Murphy, E. (2024). ROSE: A neurocomputational architecture for syntax. Journal of Neurolinguistics, 70, 101180.
Nallet, C., amp; Gervain, J. (2021). Neurodevelopmental preparedness for language in the neonatal brain. Annual Review of Developmental Psychology, 3, 41−58.
Obleser, J., amp; Kayser, C. (2019). Neural entrainment and attentional selection in the listening brain. Trends in Cognitive Sciences, 23(11), 913−926.
Pallier, C., Devauchelle, A.-D., amp; Dehaene, S. (2011). Cortical representation of the constituent structure of sentences. Proceedings of the National Academy of Sciences, 108(6), 2522−2527.
Peter, V., Goswami, U., Burnham, D., amp; Kalashnikova, M. (2023). Impaired neural entrainment to low frequency amplitude modulations in English-speaking children with dyslexia or dyslexia and DLD. Brain and Language, 236, 105217.
Poeppel, D. (2012). The maps problem and the mapping problem: Two challenges for a cognitive neuroscience of speech and language. Cognitive Neuropsychology, 29(1−2), 34−55.
Poeppel, D., amp; Assaneo, M. F. (2020). Speech rhythms and their neural foundations. Nature Reviews Neuroscience, 21(6), 322−334.
Poeppel, D., Idsardi, W. J., amp; van Wassenhove, V. (2008). Speech perception at the interface of neurobiology and linguistics. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, 363(1493), 1071−1086.
Prystauka, Y., amp; Lewis, A. G. (2019). The power of neural oscillations to inform sentence comprehension: A linguistic perspective. Language and Linguistics Compass, 13(9), e12347. https://doi.org/10.1111/lnc3.12347
Rafferty, M. B., Saltuklaroglu, T., Reilly, K., Paek, E. J., amp; Casenhiser, D. M. (2023). Neural synchrony reflects closure of jabberwocky noun phrases but not predictable pseudoword sequences. The European Journal of Neuroscience, 57(11), 1834−1847.
Si, F. (2016). Paralinguistic features and non-verbal elements in human communication. The 7th International Conference on Formal Linguistics, China, Tianjin.
Seidl, A. (2007). Infants’ use and weighting of prosodic cues in clause segmentation. Journal of Memory and Language, 57(1), 24−48.
Slaats, S., Weissbart, H., Schoffelen, J.-M., Meyer, A. S., amp; Martin, A. E. (2023). Delta-band neural responses to individual words are modulated by sentence processing. Journal of Neuroscience, 43(26), 4867−4883.
Smith, N. J., amp; Levy, R. (2013). The effect of word predictability on reading time is logarithmic. Cognition, 128(3), 302−319.
Steinhauer, K., Alter, K., amp; Friederici, A. D. (1999). Brain potentials indicate immediate use of prosodic cues in natural speech processing. Nature Neuroscience, 2(2), 191−196.
Traxler, M. J. (2014). Trends in syntactic parsing: Anticipation, Bayesian estimation, and good-enough parsing. Trends in Cognitive Sciences, 18(11), 605−611.
Xu, N., Qin, X., Zhou, Z., Shan, W., Ren, J., Yang, C., Lu, L., amp; Wang, Q. (2023). Age differentially modulates the cortical tracking of the lower and higher level linguistic structures during speech comprehension. Cerebral Cortex, 33(19), 10463−10474.
Xu, N., Zhao, B., Luo, L., Zhang, K., Shao, X., Luan, G., Wang, Q., Hu, W., amp; Wang, Q. (2023). Two stages of speech envelope tracking in human auditory cortex modulated by speech intelligibility. Cerebral Cortex, 33(5), 2215−2228.
Neural oscillations: Exploring the temporal dynamics ofsyntactic parsing
QI Ruiying, FENG Ye, SI Fuzhen
(Department of Linguistics/ Lab of Biolinguistics and Brain Sciences,Beijing Language and Culture University, Beijing 100083, China)
Abstract: The mapping problem, which aims to establish correspondences between linguistic phenotypes and neural mechanisms, is a central focus of contemporary research. Within this domain, the neural mechanisms underlying syntactic parsing are particularly challenging, as they involve identifying neural activities corresponding to the processes of syntactic structure construction. This endeavor is crucial for deciphering the enigmas of human linguistic capability. Recent investigations into neural oscillatory activity not only furnish compelling evidence for the “autonomy of syntax” of language functions in the human brain but also illustrate the feasibility of employing neural oscillations to explicate the neural coding activities underpinning syntactic parsing. The theoretical models of syntactic computation within the minimalist framework of theoretical linguistics can be corroborated by experimental research on neural oscillations in neuroscience. Such interdisciplinary studies have the potential to elucidate the temporal dynamics inherent in autonomous syntactic structure building. Future research should focus on four key areas: the fine-grained alignment between neural oscillations and syntactic processing, the mechanisms and biological significance of neural oscillations, the developmental trajectory of neural oscillations during language acquisition in children, and the neurophysiological foundations of language disorders alongside their rehabilitative applications.
Keywords: neural oscillation, language comprehension, syntactic parsing, the minimalist program, incremental conversion
* 教育部人文社会科学研究一般项目“汉语婴幼儿声调感知的大脑偏侧化模式及其对语言发展的影响” (23YJC740011)、北京语言大学校级科研项目“音乐节奏对汉语发展性语言障碍儿童语法加工的潜在影响及其神经机制研究” (24QN07)、北京语言大学院级科研项目“汉语儿童关系从句的加工模式探究” (23YJ120001)资助。
通信作者:封叶, E-mail: echofy@blcu.edu.cn
1 本文对于“先验语法知识”的工作定义, 指与后天经验相对的, 人生来具备的构建句法的知识或能力。这里的“知识”也有别于一般日常生活中所指的通过学习和积累而获得的知识, 而是指一种由生物遗传等先天因素所决定的不需要学习就拥有的一种智能。而在其他认知科学语境下先验的定义略有些许不同, 主要指与后验知识相对的, 不依赖具体数据而凭借已有的期望与知识进行的数据加工方式, 类似于(但不完全等同)知觉加工方式中的概念驱动加工方式。
[2] 理论语言学历来存在句法自治与句法涌现之争。句法自治也称“句法中心”论, 指句法作为一个相对独立的模块与通过接口(interface)与语音、韵律、语义等模块进行联系(姜孟, 2009), 即语言层级结构的生成会由于语音、语义等模块的作用而发生调整, 但其本质上由句法主导; 与句法自治理论相对, 一些学者认为句法并非独立的模块, 其结构特征是其他模块信息或一般认知能力的产物, 是基于用法的涌现(杨烈祥, 2012)。尽管目前实证研究的结果尚不能直接证明任何一方观点, 但已为先验语法知识的存在提供了重要证据。
[3] 学界这方面经典的成功实验案例可参考Chomsky和Moro (2022)中有关“结构依存”的两组实验的介绍。
[4] 在讨论神经振荡相关活动时, 不少研究者会以夹带作用(entrainment)为切入点, 但夹带作用这个语本身的定义及其机制在很多时刻都并未明晰(Obleser amp; Kayser, 2019)。一些学者对夹带作用的定义有较为严格的限制(Haegens amp; Zion Golumbic, 2018); 而另一些学者则默认夹带作用概念较为宽泛, 与外在节律有关的追踪与同步活动都可称为夹带作用。 本文旨在将语法知识与其他心理过程进行分离, 而非对概念本身进行讨论。因此, 为避免因措辞而产生歧义, 本文于此不援引夹带作用来指称相关活动。
[5] David Marr的三分框架存在一定争议(如McClamrock, 1991)。尽管如此, 它仍是非常经典的对人脑运作机制进行探讨的研究框架。
[6] 此处参考审稿专家意见进行补充说明, 感谢匿名评审专家所提出的问题。
