珙桐MADS-box基因家族的鉴定、表达及调控分析
2025-02-10刘晋芸胡小盼朱大海叶天意李培瑶魏炜


关键词: 珙桐; MADS-box 基因家族; 花瓣状苞片; 基因异位表达
1 引言
花是被子植物特有的生殖器官,其大小、形状和颜色因物种而异. 植物开花的过程由一系列基因构成一个复杂的基因调控网络响应外界环境及内源的开花信号而操纵. 编码一系列转录因子的MADS-box 基因家族在其中发挥着重要的作用,参与了从响应开花信号到花原基启动再到花器官身份确立以及花发育的整个开花过程.
在植物中具有Ⅰ类和Ⅱ类两种MADS-box 基因,Ⅰ类MADS-box 基因也称M 型基因,编码的蛋白只有SRF-like MADS-box 结构域,根据其C 末端区域的保守基序可分为Mα、Mβ 和Mγ 3 类[1]. 植物Ⅱ 类MADS-box 基因也称MIKC 型基因,其编码的蛋白除MADS-box 结构域外,还具有I(intervening)、K-box(keratin-like)和C(C-terminal)3 个结构域[2]. MIKC 型基因又可进一步分为MIKCC 型和MIKC*型基因,MIKCC 型基因编码的蛋白保留了原始的“经典结构”, 而MIKC*型蛋白的I 结构域明显长于MIKCC蛋白,K-box 结构域在序列及长度上也与MIKCC 蛋白存在差异[3]. MIKCC 型基因根据系统发育情况可划分为AG, AGL12, AGL15,AGL17, AGL2/SEP, AGL6, FLC, GGM13,PI/AP3, STMADS11, SQUA, TM3 共12 个不同的进化枝,每个进化枝的基因具有相似的表达模式和高度相关的功能.
MADS-box 基因编码的蛋白在响应开花信号调控开花时间,花器官身份决定,种子及果实发育以及根的发育等多方面发挥重要的调控作用,其在花器官身份决定中的作用尤其受到关注. 一朵典型的花通常由四轮花器官按同心圆的形式排列,从外到里分别为萼片、花瓣、雄蕊和心皮. 研究人员们通过突变体植株中花器官的同源转化发现花器官身份的确立由一系列花器官特征基因指定[4]. 并基于这些基因的突变体表型及遗传相互作用,经过不断发展、补充和修订,最终建立了花器官身份决定的“ABCDE”模型,将这些花器官特征基因分为5 类并对它们的功能进行了描述. 根据“ABCDE”模型:A 类基因指定萼片的形成;B 类基因和A 类基因共同指定花瓣的形成;B 类基因和C类基因共同指定雄蕊的形成;雌蕊的形成由C 类基因单独指定;D 类基因调控胚珠的发育;E 类基因参与每一轮花器官的形成[5-8]. 目前已鉴定的所有花器官特征基因中除了 A 功能基因 APETALA2(AP2) 外都编码 MIKCC 型MADS-box 转录因子[9, 10]
在花器官身份建成时,花器官特征基因的表达被限制在特定的器官轮里. 而花器官特征基因在其他的花器官区域发生异位表达,能够将其影响扩大到不同的花器官轮,从而导致花器官的同源转化[10, 11]. 在矮牵牛和番茄叶中证明了转基因植株异位表达AG 基因可以将萼片转化为心皮,花瓣转化为雄蕊. AP2 基因在第三轮花器官的表达导致雄蕊转变为花瓣[12-14]. 在一些花被未分化的单子叶植物中,呈现两轮一样的花瓣状花被,例如郁金香[11]和百合[15],在它们的两轮花被中都检测到了B 类基因的表达. 在双子叶植物中也有这样的例子,例如枇杷[16]和小花瓜莲[17]. 除了花器官外,一些物种在苞片中也表现出花瓣的特征,珙桐(Davidia involucrata)的花尤其特别,其花被完全缺失,且花序轴极度缩短形成一个球形的头状花序,使整个花序看起来像一朵花. 花序外侧生长的一对(少数为单瓣或三瓣)白色苞片取代花瓣,以吸引传粉者和保护花粉免受雨水的侵袭. 并且随着花序的生长,苞片有明显的由叶状向花瓣状过渡的过程. 这些特征表明,珙桐具有较高的科学价值,是研究花器官特性在花外建立和花瓣状苞片发育的理想材料.
研究人员已经认识到MADS-box 家族基因在珙桐苞片发育中的重要性,并对其部分基因进行了研究,但仅局限于少数几个基因的克隆及表达定量[18, 19]. 目前仍需要对珙桐的MADS-box 基因家族进行更全面、系统的研究,以了解其对其花器官特性和花瓣状苞片发育的影响. 本研究将基于Chen 等[20]发布的珙桐基因组组装,在全基因组范围内对珙桐MADX-box 家族基因进行鉴定,并通过与近缘物种的比较探索该基因家族的进化. 进一步,通过转录组测序分析MADS-box 在苞片发育过程中的表达模式并预测其潜在的靶基因,探究MADS-box 基因家族对珙桐花瓣状苞片形成的调控机制,对被子植物开花调控网络进行进一步的补充和完善.
2 材料与方法
2. 1 材料
2. 1. 1 植物材料 珙桐苞片及叶片材料取于四川省成都市都江堰龙溪-虹口国家级自然保护区.分别于2018 年4 月15 日、4 月22 日、5 月7 日采集开花早、中、晚期的珙桐苞片和叶片(对应的苞片分别为绿色、淡绿色和白色). 根据时间由早到晚,将3 个时期的苞片和叶片分别标记为B-1、B-2、B-3和L-1、L-2、L-3. 每组苞片和叶片各取3 个生物重复.
2. 1. 2 数据来源 本研究使用的珙桐及近缘物种基因组及注释来源:珙桐全基因组序列和注释从国家基因组科学数据中心(National GenomeData Center,https://ngdc. cncb. ac. cn/)获取,登录号为GWHABJS00000000[20]. 近缘物种喜树(Camptotheca acuminata)染色体规模基因组组装及注释数据从Figshare 数据库获得[基因组组装:https://doi. org/10. 6084/m9. figshare. 12570599;注释:https://doi. org/10. 6084/m9. figshare. 12570614][21]. 蓝果树(Nyssa sinensis)[22]和山茱萸(Cornus florida)的全基因组信息均从NCBI 数据库下载,登录号分别为:GCA_008638375. 1 和GCF_030987335. 1.
2. 2 方法
2. 2. 1 珙桐MADS-box 家族基因鉴定 以拟南芥MADS-box 蛋白序列为参考序列,使用Ge‑MoMa-1. 9[23]软件,通过tblastn 策略在珙桐基因组范围内鉴定可能的MADS-box 基因座位. 基于与Swisse-Prot 数据库的比对结果将新基因座位与基因组注释去冗余获得候选基因. 将全部候选基因编码的氨基酸序列使用Interproscan[23],利用Pfam数据库(http://pfam. xfam. org/)进行注释. 提取注释结果中包含PF00319(SRF-box)和PF01486(MIKCC 型 K-box)结构域的序列为珙桐MADSbox蛋白序列.
2. 2. 2 珙桐MADS-box 基因系统发育分析及分类 利用鉴定出来的珙桐MADS-box 基因编码的最长氨基酸序列进行系统发育分析. 长度大于100aa 的序列与拟南芥MADS-box 基因编码的蛋白质序列一起,利用MAFFT 软件[24]进行多序列比对. 使用ProtTest 3[25]软件检测序列建树的最佳模型,然后使用RAxML[26]基于最大似然法构建ML 进化树,并根据分析结果对MADS-box 基因进行分类. 长度小于100aa 的序列与NR 数据库进行比对,参考比对结果进行分类.
2. 2. 3 山茱萸目物种MADS-box 基因家族同源性研究 对喜树、蓝果树、山茱萸使用与珙桐相同的方法分别鉴定物种中的MADS-box 基因并分类. 并将在珙桐及近缘物种中鉴定出来的MADSbox家族基因使用OrthoFinder-v2. 4. 0[27]进行同源性分析.
2. 2. 4 珙桐MADS-box 基因家族特征分析 将鉴定出来的珙桐MADS-box 蛋白氨基酸序列利用MEME[28]进行motif 预测. 利用Expasy[29]的Protparam工具(https://web. expasy. org/protparam/)分析编码蛋白的理化性质. 另外分别使用TMHMM2. 0[30]和SignalP-6. 0[31]软件对蛋白质氨基酸序列的跨膜结构域和信号肽结构进行预测.2. 2. 5 珙桐MADS-box 基因家族进化分析 使用MCScanX[32]基于默认参数进行物种内共线性分析及复制类型分类. 随后根据物种内共线性分析得出的共线性基因对,利用基因的CDS 序列对共线性基因对之间的碱基非同义替换率(Ka)和同义替换率(Ks)进行计算. 最后对M type, MIKC*和 MIKCC 三种类型的MADS-box 基因的Ka/Ks进行计算,分析珙桐不同类型MADS-box 基因所受到的选择压力.
2. 2. 6 珙桐MADS-box 基因在苞片发育过程中的表达模式分析 从三个发育时期的18 例苞片及叶片样本中提取总RNA 进行转录组测序. 转录组文库构建和测序服务由华大基因提供,使用BGISEQ-500 测序平台进行. 测序原始数据已上传至国家基因组科学数据中心GSA 数据库(CRA013964),可通过https://ngdc. cncb. ac. cn/gsa 公开访问. 使用SOAPnuke 短序列过滤软件和Trimmomatic[33]对原始数据进行过滤获得Cleanreads. 使用HISAT2[34]将Clean reads 比对到珙桐基因组. 利用Stringtie 软件[35]对不同发育阶段的苞片和叶片中的基因表达进行定量分析. 采用DEseq2 软件[36]进行差异表达分析,差异表达基因筛选标准为log2FC 绝对值大于等于1 且FDR 小于0. 01.
2. 2. 7 基因表达模式的qRT-PCR 验证 qRT-PCR 使用Hieff UNICON® Universal Blue qPCRSYBR Green Master Mix (Yeasen; Shanghai,China)基于CFX Connect Real-Time PCR DetectSystem 平台进行. 扩增程序为:95 ℃预变性2 min,95 ℃变性10 s,60 ℃退火/延伸30 s,40 个循环. 使用珙桐β-DiACTIN 基因作为内参基因,发育早期的叶片(L-1)作为对照组,使用2−ΔΔCt法计算基因的相对表达量. qRT-PCR 使用的引物见表1.2. 2. 8 珙桐MADS-box 转录因子靶基因预测 利用 JASPAR 数据库( https://jaspar. elixir. no/)中拟南芥SVP, AP3, AP1 和SEP3 转录因子结合位点的motif 分别在基因转录起始位点上游2000 bp范围内识别潜在的结合位点. 然后在转录调控区具有结合位点的基因中根据pearson 相关性系数,筛选与对应珙桐同源基因表达量相关性gt;0. 70或lt;-0. 70 且Plt;0. 01 的基因作为珙桐同源转录因子的潜在靶基因.
3 结果
3. 1 珙桐MADS-box 基因鉴定及分类
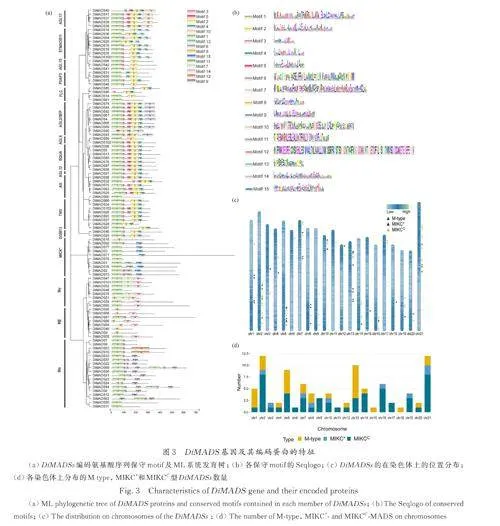
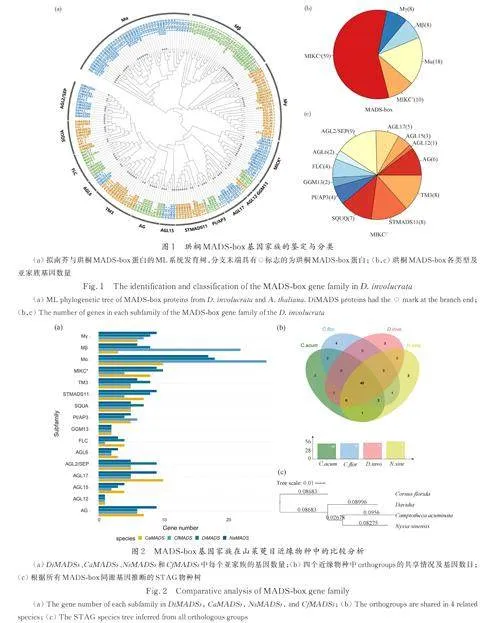
在珙桐基因组中共鉴定出103 个MADS-box家族基因,将这些基因分别命名为DiMADS1-103,其编码的蛋白质分别命名为DiMADS1-103.根据系统发育树的结果( 图1a),参考拟南芥MADS-box 基因家族对103 个DiMADSs 进行了分类. 结果表明珙桐含有34 个M type(Ⅰ类)和69 个MIKC(Ⅱ 类)基因(图1b). 其中Ⅰ 类MADS 基因中包含18 个Mα、8 个Mβ 和8 个Mγ 分支基因. Ⅱ类MADS 包括10 个MIKC*类基因和59 个MIKCC类基因,每个MIKCC亚家族有1~9 个成员(图1c).可以发现珙桐在AGL2/SEP 亚家族、SQUA 亚家族以及STMADS11 亚家族中的基因数量远超拟南芥,说明这几个MADS-box 基因家族的分支可能在珙桐的进化过程中发生了明显的扩张,并且有可能导致这些基因的功能在珙桐中的进一步分化. DiMADS76 由于序列较短,未用于构建系统发育树. 根据与Nr 数据库的比对结果将其归类为MIKC*型基因.
3. 2 山茱萸目植物MADS-box 基因家族比较分析
为了进一步分析珙桐MADS-box 家族基因的进化,以及该基因家族在珙桐及其近缘物种中的变化. 使用了同样方法分别在喜树、蓝果树、山茱萸中鉴定出85、102、103 个MADS-box 家族基因并命名为CaMADS1-85、NsMADS1-102 和Cf⁃MADS1-103. 如亚家族基因数目统计结果所示,4个物种基因数目的主要差异来源于Ⅰ类MADS 基因的差异. 值得注意的是,珙桐的AGL2/SEP 亚家族和SQUA 亚家族的基因数目除了之前发现与拟南芥相比有显著扩张之外,与它的近缘物种相比也存在明显的扩张(图2a).
对4 个物种的MADS-box 基因家族进行同源性分析. 共鉴定出63 个orthogroups,其中有40 个在4 个近缘物种中共享,包含68 个DiMADS、72 个CaMADS、56 个CfMADS 和62 个NsMADS. 珙桐、喜树、蓝果树、山茱萸各有24、4、18、38 个物种特异性基因(图2b). 说明MADS-box 基因家族在这四个近缘物种中可能具有不同的进化历史,在功能上也可能存在差异.
根据所有MADS-box 同源基因构建的STAG物种树显示,在蓝果树科中喜树与蓝果树的亲缘关系比跟珙桐更近. 而属于另一个科的物种山茱萸与其他3 个物种的亲缘关系最远(图2c). 这与此前Qi 等[37]和Li 等[38]利用叶绿体基因组构建的山茱萸目和蓝果树科的物种系统发育树相符合,可以为珙桐的系统发育分类提供一份来自核基因的证据.
在DiMADSs 编码的氨基酸序列中共鉴定了15个保守的 moti(f 图 3a). 其中 99个 DiMADSs氨基酸序列含有motif 1 或motif 8. 通过SMART[39]进行评估,motif 1 和motif 8 都与SRF 结构域高度相关. 结合这两个motif 在序列上的位置和motif的Seqlogo(图3b)进行推断,motif 8 可能是MADS结构域的核心区域. 同样根据SMART 评估,Motif2、4、5 与K-box 结构域高度相关,其中motif 2 为核心区域. 共有57 条MIKCC 型序列(57/59)包含K-box 结构域. 另外,MIKC*类中的motif 15 也与K-box 结构域相关,但是与motif 2 不同. 并且MIKC*类的SRF 结构域和K-box 结构域之间的插入区段明显比MIKCC 类更长. 这两点均符合二者的特征差异. 此外,部分进化支(或亚家族)还具有其特征性的保守基序. 综上所述, SRF 结构域是MADS-box 基因家族中最保守的结构域,而K-box是Ⅱ型基因的保守结构域. 序列C 端的保守性在总体上较低,但在其亚家族内也有一定程度的保守性.
除了DiMADS76 以外,DiMADSs 的编码序列(CDS)长度范围为327 bp 至2070 bp,编码蛋白的氨基酸残基数介于108 至698 之间,分子量介于12883. 13 至77751. 06 kDa 之间. 理论等电点(pI)介于4. 36 至10. 36 之间. 所有DiMADSs 均为亲水性蛋白,无信号肽,多数为不稳定蛋白. 仅有3个DiMADS 蛋白含有1 到2 个跨膜结构.
DiMADS 基因的染色体定位如图3c、3d 所示. 103 个基因中,有95 个基因定位于珙桐基因组组装的21 条染色体上,8 个基因定位于未组装到染色体的片段上. Chr2、Chr 5、Chr13、Chr21 分布了9 个及以上的MADS-box 基因,其余染色体上皆分布有1~6 个MADS-box 基因. MIKCC 类基因分布最为广泛,除Chr15 和Chr18 外,每条染色体上均有MIKCC类基因的分布.
3. 4 珙桐MADS-box 基因家族进化分析
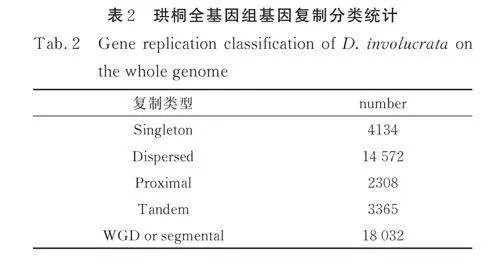
为了进一步研究发生在珙桐MADS-box 基因家族中的复制事件,对珙桐基因组进行了基因复制及种内共线性分析. 如表2 所示,基因被分为五类. 在珙桐103 个MADS-box 家族基因中,最多的是由全基因组或片段复制相关的基因. 珙桐MADS-box 基因家族中没有基因被分类为单拷贝基因,这可能与MADS-box 基因家族的起源与扩张有关(表3).
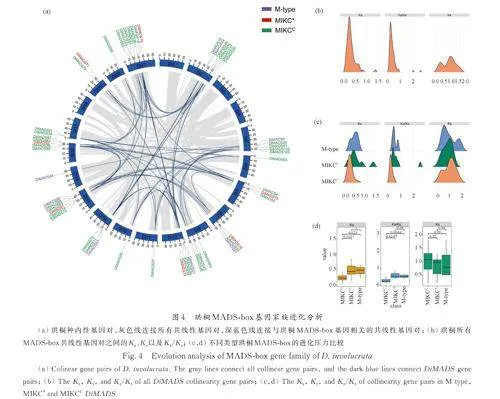
55 个珙桐MADS-box 家族基因关联于48 个共线性块中的66 个共线性基因对之中(图4a),说明某些珙桐MADS-box 基因在进化过程中发生了多次的复制. 大多数共线性基因对中的DiMADSs属于同一个亚家族,在一定程度上说明了亚家族基因结构域保守性的形成原因.
对DiMADSs 的进化压力进行分析发现,仅MIKC* 亚家族的DiMADS71-DiMADS15 和Di⁃MADS71-Dinv44740. t1( 非MADS)两对基因的Ka/Ks 大于2,基因受正选择,可能正在发生功能上的分化. 其余基因对之间的Ka/Ks 均小于1,且大多数基因对的Ka/Ks 小于0. 5. 证明绝大多数珙桐MADS-box 基因受到纯化选择. 对比Ⅰ型M type基因和Ⅱ 型基因中的MIKC*和MIKCC 类MADSbox基因. M type 类基因和MIKC*类基因的Ka、Ks以及Ka/Ks 均无显著性差异. MIKCC 类基因的Ka以及Ka/Ks 值显著低于M type 基因和MIKC*类基因,说明MIKCC 类基因受到更为强烈的纯化选择(图4b~4d). 这应当与MIKCC 类基因在高等植物发育过程中的重要作用有着密切关联.
3. 5 DiMADSs 在苞片发育过程中的表达模式
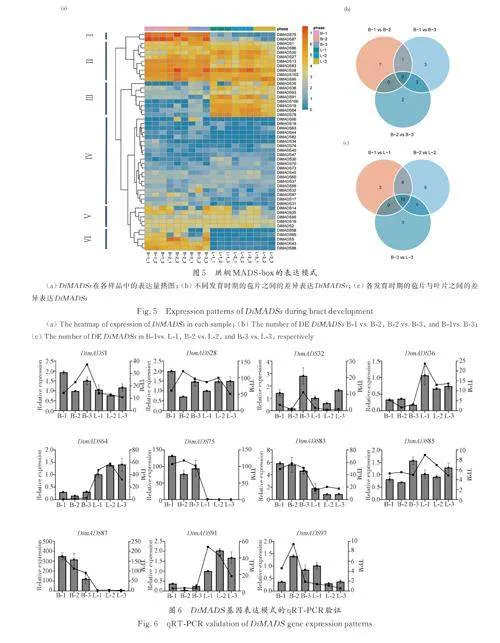
苞片是珙桐生殖发育过程中的一种特殊结构. 在生长初期,苞片呈绿色,形状和颜色与同时期的叶片相似,二者主要根据着生位置区分. 随着发育的进行,两个器官的形态差异逐渐增大. 为了探究MADS-box 基因在苞片变化中的作用,本研究通过转录组测序检测了这些基因在花期早、中、晚期的苞片和叶片中的表达模式. 根据聚类结果可以将DiMADSs 的表达谱分为6 类. 类别Ⅰ和Ⅵ包括2 个AGL2/SEP 亚家族基因、4 个SQUA 亚家族基因和1 个Mβ 亚家族基因,这些基因主要在苞片中表达,在叶片中低表达或不表达. Ⅲ类基因在叶片发育的不同阶段都有较高水平的表达,但在苞片发育过程中受到抑制. 在Ⅲ类的8 个基因中,有6 个是拟南芥SVP 的同源基因,SVP 在拟南芥中具有抵抗营养生长向生殖生长转变的功能. 另外2 个为拟南芥TM3 类基因AGL42、AGL71 和AGL72 的同源基因,这些基因在拟南芥中与开花整合因子密切相关,参与花转化(图5a).
对不同发育时期的苞片进行比较,在B-1vs.B-2、B-2vs. B-3 以及 B-1vs. B-3 中分别鉴定出8 个、5 个以及7 个差异表达的DiMADSs. 这一结果暗示了不同的DiMADS 基因可能在苞片发育的特定阶段发挥着重要的调控功能. 进一步对各发育阶段的苞片和叶片的差异表达基因进行分析.在B-1vs. L-1、B-2vs. L-2 和B-3vs. L-3 中分别鉴定出19 个、22 个和14 个差异表达的DiMADS. 似乎有更多的DiMADS 在发育的早期和中期决定苞片和叶片的差异. 10 个基因在3 个发育时期的苞片和叶片中均存在差异表达,其中属于SQUA 亚家族的DiMADS5、DiMADS13、DiMADS75、Di⁃MADS83、DiMADS87 和DiMADS88 以及属于AGL2/SEP 亚家族的DiMADS43 始终在苞片中高表达. 而属于STMADS11 亚家族的Di⁃MADS19、DiMADS64、DiMADS78 始终在叶片中高表达.
3. 6 DiMADSs 基因表达模式的qRT-PCR 验证
随机选择了11 个具有不同表达模式的Di⁃MADS 基因,以珙桐β-DiACTIN 基因作为内参基因,以L-1 叶片样本为对照组计算了基因的相对表达量. 结果如图6 所示,直方图表示qRT-PCR 检测的相对表达量,折线图表示转录组定量中TPM数据. 二者之间虽然存在一定的差异,但是考虑到用于测序的cDNA 和用于qRT-PCR 的cDNA 可能存在批次效应带来的差异,而且这些基因在苞片和叶片中的表达变化趋势与转录组数据基本一致,表明基于转录组测序的定量结果是基本可靠的.
3. 7 DiMADS 转录因子的靶基因预测
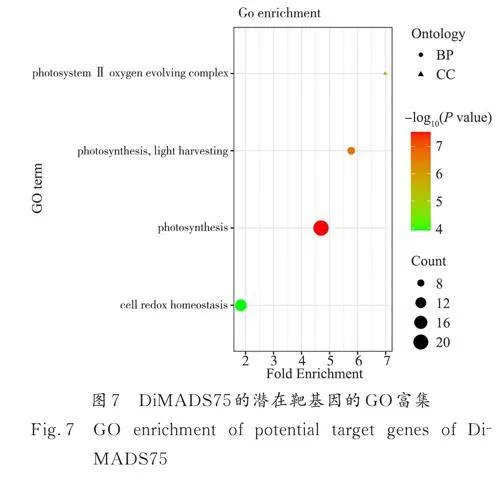
在珙桐基因组中分别预测了201 个珙桐AP3-like 转录因子DiMADS31 的潜在靶标,1100 个珙桐AP1-like 转录因子DiMADS75 的潜在靶基因,182 个珙桐SEP-like 转录因子DiMADS43 的潜在靶基因,以及672 个SVP-like 转录因子DiMADS36的潜在靶基因. DiMADS31、DiMADS43 以及Di‑MADS36 潜在靶基因没有明显的功能富集,但Di‑MADS75 的潜在靶基因在光合作用以及细胞氧化还原稳态的生物学过程(BP)有所富集(图7). 并且与富集到的光合相关基因均有极强的负相关关系.
此外,一些DiMADS 基因也是这些转录因子的潜在靶基因. 另外,DiMADS36 在其自身的转录调控区域也存在结合位点. 这进一步说明了珙桐开花的基因调控相当复杂,这些调控基因之间可能形成相互的反馈网络,在不同的发育阶段控制着基因表达的平衡.
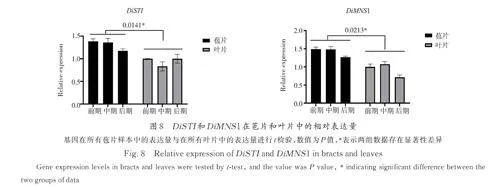
3. 8 潜在靶基因DiSTI 和DiMNS1 的表达模式
根据花器官建立的ABCDE 模型,花瓣的性状由A、B 以及E 类基因共同控制. 并且根据花的四因子模型. AP3、PI 以及AP1、SEP 将形成转录因子四聚体结合两个CArG box 介导DNA 环化并调控转录(AP3-PI 形成异源二聚体结合其中一个CArG box). 在预测的靶基因中有两个基因同时是DiMADS31、DiMADS75、DiMADS43 的潜在靶标,并且SEP3 和AP1 与AP3 在转录调控区有不同的结合位点,说明AP3-PI-AP1-SEP 转录因子四聚体有可能结合到这两个基因的转录调控区域并调控其表达. 这两个基因编码的蛋白分别是STICHEL(STI)蛋白与Mannosyl-oligosaccharide1,2-alpha-mannosidase MNS1( α-1,2-甘露糖苷酶1)的同源物,因此将它们分别命名为DiSTI 和DiMNS1. 以L-1 时期为对照组,利用qRT-PCR 检测了这两个基因在苞片及叶片中的相对表达量.结果表明,这两个基因在苞片中的表达量显著高于其在叶片中的表达量( 图8). DiMADS31,Di‑MADS75,DiMADS43 可能促进了这两个基因在苞片中的表达.
4 讨论
目前,MADS-box 基因家族在大量开花物种中的鉴定和克隆以及功能研究证明了其在植物生长发育及繁殖中的重要调控作用[14, 40, 41]. 目前关于被子植物开花调控的研究虽然已经有很多,但开花过程中的复杂基因调控网络还没有完全清晰,目前对花器官特征基因的功能描述也不足以解释所有自然界中形态万千的花. 珙桐作为第三纪古热带孑遗植物,尚有丰富的遗传资源等待挖掘[42]. 其特殊的花器官结构及苞片结构表明珙桐是研究花器官身份决定的良好材料. 珙桐MADSbox基因家族进行鉴定并对其在苞片发育过程中的表达模式进行研究,将有助于加深对被子植物开花调控网络的理解. 此外对珙桐遗传资源的保护、开发和利用也具有重要的意义.
在本研究中,成功鉴定了103 个珙桐 MADSbox基因,并分析了这些基因及其编码蛋白的特征. MADS-box 蛋白作为转录因子,通过与特定的顺式作用元件结合来调控下游基因的表达. 对于结构域的功能,SRF-box(MADS 结构域) 决定了转录因子与DNA 序列的结合,而K-box 促进MADS-box 转录因子发挥功能所必需的二聚化[3].二者的缺失可能导致基因功能的改变或丧失. 在本研究中发现FLC 亚家族中的DiMADS85 和Di⁃MADS86 缺失MADS 结构域,并且在不同时期的苞片和叶片中显著表达. 研究表明,过表达截短的转录因子可能通过竞争性结合有功能的转录因子形成无功能的异二聚体而干扰正常功能[43]. FLC亚家族基因是控制开花时间的抑制因子,通过FT间接抑制下游花器官识别基因AP1 的表达[44]. 因此推测FLC-like 截短基因的表达可能是影响珙桐特殊花结构的潜在因素. 但其是否参与以及具体如何调控珙桐开花还有待进一步研究.
在基因组水平上比较了4 个物种的MADSbox基因家族. 除在物种间具有同源性的基因外,每个物种还存在多个物种特异性的MADS-box 基因. 这意味着MADS-box 基因家族在这四个近缘物种的进化过程中出现了分歧. 值得注意的是,珙桐的AGL2/SEP 和SQUA 亚家族的基因数目与拟南芥和近缘物种相比都有显著的增加. 这两个亚家族的成员在拟南芥和其他许多物种中都已被证明在植物开花调控中发挥着极其重要的作用.在拟南芥中,SQUA 亚家族包括4 个成员AP1、CAL、FUL 和AGL79,前三者在调控花分生组织决定中具有冗余的功能[45, 46]. 而AP1 更是调控萼片和花瓣发育所必需的基因[47]. 该亚家族的另一个同源基因AGL79 也被证明可以通过抑制TFL1的表达正向调控开花[48]. AGL2/SEP 亚家族的成员是花器官身份决定的E 类基因,在所有器官轮中发挥功能,其他花器官特征基因编码的转录因子需要与SEP 蛋白结合形成高阶蛋白质复合物才能正确激活下游开花基因的表达[7, 49]. 珙桐AGL2/SEP 亚家族和SQUA 亚家族的扩张,说明与模式种及其近缘物种相比,珙桐对花分生组织和花器官身份建立可能存在更为复杂的调控模式. 后续可以通过转基因实验来验证这些复制产生的旁系同源基因的功能是否发生了亚功能化,是否具有功能冗余.
本研究通过转录组测序和qRT-PCR 验证,发现AP3(B 类)、AP1(A 类)以及SEP(E 类)的同源基因在苞片中表现出较高的表达水平. P(I B类)的同源基因在苞片和叶片中持续表达. 而已有研究表明AP3、PI 与AP1 或SEP3 的异源表达可以将植物营养叶转化为花瓣状器官[6, 49, 50]. 据此推测,珙桐花瓣状苞片的形成可能是由AP1-like、AP3-like 和SEP-like 的同源基因在苞片中的异位表达所导致.
进一步对这珙桐这三类转录因子的靶基因进行预测并筛选到两个可能符合四因子模型[50]调控的基因DiSTI 和DiMNS1. 通过qRT-PCR 检测发现它们在苞片中的表达显著高于叶片,说明AP1-like、AP3-like 和SEP-like 的同源转录因子可能促进了它们在苞片中的表达. DiSTI 编码的STICHEL 蛋白在拟南芥中被证明与毛状体分支有关[51],DiMNS1 编码的α-1,2-甘露糖苷酶则是天冬酰胺(N)- 聚糖的生物合成中的一种关键酶,N-聚糖在植物开花、种子形成以及胁迫响应等方面发挥着重要的作用[52]. 而二者在珙桐苞片发育中的作用还有待进一步探索. 此外,通过靶基因的功能富集发现珙桐AP1-like 转录因子DiMADS75可能作为直接的抑制因子,参与调控部分光合作用相关基因的表达,抑制了苞片的光合功能. 同时参与了苞片与叶片发育细胞氧化还原稳态相关基因的表达调控.
