植物生长调节剂对青钱柳光合积累及黄酮合成的影响
2024-12-27赵睿智徐道炜何中声刘金福郭相亿李伟森张朝鹏








关键词:青钱柳;植物生长调节剂;叶绿素含量;光合积累;黄酮
青钱柳Cyclocaryapaliurus为胡桃科Juglandaceae青钱柳属Cyclocarya的单种属植物,是一种兼具生态效益与经济效益的重要树种[1]。青钱柳叶片味道甘甜,含有黄酮、三萜、多糖等多种活性成分,能够增强人体免疫力,减轻“三高”(高血压、高血糖、高血脂)对人体的危害,具有较高的食用及药用价值[2-3]。青钱柳具有速生特性,普遍存在树高徒长、侧枝萌发率低等问题,导致采摘难度增加和成本上升,同时叶片产量不高,这些因素限制了我国青钱柳产业的规模化发展[4]。采用人工矮化技术进行生长调控是解决上述问题的有效方法。
植物生长调节剂是人工合成的(或从微生物中提取的)一类植物激素,通过模拟或调节植物内源激素,能够有效削弱植株顶端优势,促进侧枝萌发,进而达到矮化植株、增加分枝密度等目的[5-6]。苏宇霞等[7]发现施用多效唑1500mg/L、矮壮素500mg/L及缩节胺1000mg/L,能够有效抑制无花果树体及叶片的营养生长。张钰晗等[8]发现喷施三碘苯甲酸能够显著抑制‘骨里红’梅花的苗高增长并增加新梢生长量,但会对其光合作用产生不同程度的抑制。管婧等[9]对姜黄品种‘桂热1号’喷施0.1mg/L芸苔素内酯叶面肥后,发现姜黄总黄酮含量显著提高,相比对照处理提高了128.99%。在农业生产上,植物生长调节剂在多种经济林(如苹果[10],油茶[11]等)的矮化栽培中已得到广泛应用,是实现青钱柳矮化的理想途径。
在此背景下,深入研究植物生长调节剂对青钱柳光合积累过程的影响尤为重要。光合积累不仅关乎植物的生长发育和干物质积累[12],还直接影响目标活性成分(如黄酮)的合成与累积[13]。探究植物生长调节剂对青钱柳光合积累的影响,不仅可评估矮化效果,还可揭示其在促进植株矮化的同时能否保持或增加叶片中有效成分的含量。另外,研究植物生长调节剂对黄酮合成关键酶苯丙氨酸解氨酶(phenylalaninammonia-lyase,PAL)和查尔酮合成酶(chalconesynthase,CHS)活性的调节作用,可以进一步揭示其对青钱柳次生代谢通路的深层影响机制。谢寅峰等[14]发现在培养基中添加适量的烯效唑能够显著提高青钱柳试管苗的叶绿素与可溶性糖含量,郭楠等[15]发现对青钱柳幼苗喷施适宜浓度的多效唑叶面肥,不仅提高了可溶性糖含量,还提高了PAL活性,促进了有效成分黄酮的合成。然而,关于植物生长调节剂处理下青钱柳光合积累及黄酮合成相关指标变化规律的研究鲜见报道。
本研究选取3种常用植物生长调节剂(矮壮素、三碘苯甲酸、芸苔素内酯),分别设置4个浓度水平,研究其对1年生青钱柳幼苗光合积累和黄酮合成的影响,旨在揭示青钱柳幼苗在植物生长调节剂作用下的生理响应机制,为建立青钱柳矮化经济林高效培育技术体系提供参考。
1材料与方法
1.1研究地概况
试验地位于福建省福州市福建农林大学林学院试验田(119°13′42″E,26°05′15″N)。试验区域属亚热带季风气候,地势平坦,无霜期326d,年平均日照时长1700~1980h,年平均降水量900~2100mm,年均相对湿度约77%。试验期间最低气温17℃,最高气温38℃,平均气温24℃[16],适宜青钱柳幼苗生长。在试验田内专用的金属苗床摆放育苗盆,苗床上方安装有遮光率为50%的遮阳网。
1.2试验材料
1年生青钱柳幼苗来源于江西省九江市青钱柳种植基地。育苗盆规格为24cm×20cm×27cm,培养基质主要原料为珍珠岩、黄土和泥炭土的混合物(体积比1∶1∶2),基质pH值为4.56,电导率为0.077S/m,全氮含量为0.921g/kg,全磷含量为0.463g/kg。
1.3试验设计
2022年4月中旬,将青钱柳幼苗移栽至育苗盆内,为保证幼苗生长条件和初始指标的一致性,缓苗3个月,缓苗和试验期间保证良好的通风和光照条件,每2天浇水1次。选用矮壮素溶液、三碘苯甲酸溶液、芸苔素内酯溶液作为叶面肥进行喷施(表1),以清水(CK)作为对照处理,共设置13个处理。每个处理设置3次重复,每个重复处理10株青钱柳幼苗。喷施植物生长调节剂溶液时以叶面布满水珠而不滴水为宜,每10d喷1次,共喷施3次。在试验期间统一进行正常的栽培管理,保证所有青钱柳幼苗生长条件一致,于2022年12月中旬结束试验。
1.4指标测量
1.4.1叶绿素含量测定
2022年12月中旬,选取每株幼苗中间位置的3片叶,使用便携式叶绿素测定仪(SPAD-502PLUS,KonicaMinoltaHoldingsInc,日本)测定叶绿素相对含量(SPAD值),取平均值作为此株幼苗的叶绿素相对含量。
1.4.2能量积累指标测定
采集每株幼苗的成熟功能叶,用混合采样法将每个重复中幼苗的叶片混合为1个样品,共采集样品39份。将样品带回实验室清洗、保存。可溶性糖含量(solublesugarcontent,SSC)和可溶性淀粉(starchcontent,SC)含量均采用蒽酮比色法测定,非结构性碳水化合物(non-structuralcarbohydratecontent,NSC)含量为SSC与SC的总含量[17],每个样品重复测定3次,取平均值。
1.4.3生物量指标测定
2022年12月中旬,在每个重复中挑选3株长势基本一致且无病虫害的幼苗进行收获,将根、茎和叶附着的杂质清理干净并分装于塑封袋中,带回实验室用去离子水清洗,再使用滤纸吸干表面水分。置于105℃烘箱中杀青30min,在65℃条件下烘干至恒定质量。使用电子天平(精度0.0001g)称量并记录根、茎、叶、枝的生物量,计算光合与非光合组织生物量的比值(即叶与根、茎、枝生物量的比值)。
1.4.4叶片总黄酮含量测定
精确称取叶片粉末样品0.3g,加入70%乙醇20mL,摇匀封口,在50℃条件下超声波(300W,33kHz)辅助提取60min,用微孔滤膜过滤掉残渣,取上清液置于4℃冰箱待用。总黄酮含量采用比色法测定[18]。
1.4.5叶片黄酮合成酶活性测定
精确称取叶片粉末样品0.5g,研磨至匀浆,加入9mL缓冲液,在4℃、1000r/min条件下离心20min,取上清液置于4℃冰箱待用。PAL和CHS活性分别使用植物苯丙氨酸解氨酶ELISA检测试剂盒(CB10016-Pt,科艾博生物,上海)、植物查尔酮合成酶ELISA检测试剂盒(CB10144-Pt,科艾博生物,上海)测定,每处理均设置3次重复。
1.5数据处理
采用R4.3.2软件进行ANOVA方差分析、LSD多重比较以及主成分分析,通过KMO和Bartlett球形检验确保主成分分析结果的有效性,采用Origin2022软件绘图。
2结果与分析
2.1植物生长调节剂对幼苗叶绿素含量的影响
由图1可知,在苗期喷施植物生长调节剂会对青钱柳幼苗叶片叶绿素含量产生一定的影响。矮壮素处理中,叶绿素含量在C1处理下达到最高,且显著高于CK处理,叶绿素含量比CK处理增加了35.5%;三碘苯甲酸处理中,叶绿素含量随三碘苯甲酸溶液质量浓度增加呈现上升趋势,在T4处理下达到最高,相较于CK处理增加了24.5%;芸苔素内酯处理中,幼苗叶绿素含量均高于CK处理,但处理间差异均不显著。
总体上,3种植物生长调节剂均提高了青钱柳幼苗叶片的叶绿素含量,其中矮壮素对叶绿素含量的促进效果最为显著。
2.2植物生长调节剂对幼苗能量积累指标的影响
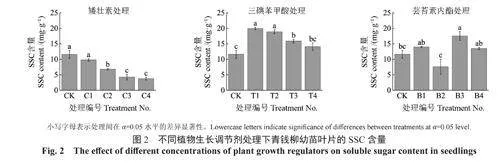
由图2可知,幼苗叶片SSC含量随着矮壮素和三碘苯甲酸质量浓度的增加呈现下降趋势,但矮壮素处理下幼苗叶片SSC含量均低于CK处理,在三碘苯甲酸处理下叶片SSC含量均高于CK处理。芸苔素内酯处理的幼苗叶片SSC含量仅B3处理显著高于CK处理。在所有处理中,T1处理的幼苗叶片SSC含量最高,相较于CK处理增加了71.6%;C4处理含量最低,相较于CK处理减少了67.5%。
由图3可知,矮壮素处理的幼苗叶片SC含量随着矮壮素质量浓度的增加呈现下降趋势,且均显著低于CK处理。三碘苯甲酸处理的幼苗叶片SC含量均显著高于CK处理,在T3处理达到最大。芸苔素内酯处理中除B2处理的叶片SC含量显著低于CK处理,其他处理与CK处理无显著差异。所有处理中,T3处理的幼苗叶片SC含量最高,相较于CK处理增加了21.7%。
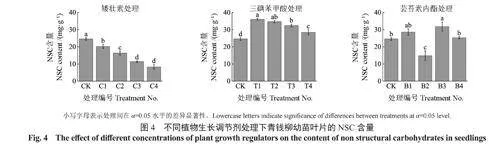
由图4可知,矮壮素处理和三碘苯甲酸处理的幼苗叶片NSC含量随着喷施溶液质量浓度增加呈现下降趋势,矮壮素处理的幼苗叶片NSC含量均显著低于CK处理,而三碘苯甲酸处理的幼苗叶片NSC含量均显著高于CK处理。芸苔素内酯处理的幼苗叶片NSC含量随溶液质量浓度的增加呈现波动趋势,在B3处理达最大,显著高于CK处理。在所有处理中,T1处理的幼苗叶片NSC含量最高,相较于CK处理增加了29.1%;C4处理的幼苗叶片NSC含量最低,相较于CK处理减少了66.1%。
总体上,喷施适当质量浓度的三碘苯甲酸和芸苔素内酯会促进青钱柳幼苗的能量积累,相较于芸苔素内酯处理下能量指标的波动现象,三碘苯甲酸对能量积累的促进效果更加稳定,不同质量浓度矮壮素溶液对青钱柳幼苗的能量积累均起到了抑制作用。
2.3植物生长调节剂对幼苗光合与非光合组织生物量分配的影响
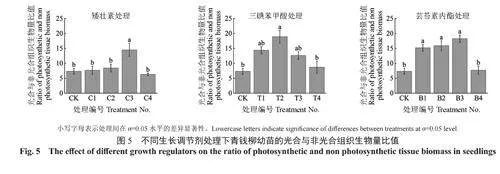
从图5可以看出,矮壮素处理的幼苗光合与非光合组织生物量比值仅在C3处理下显著高于CK处理,比CK处理提高了97.6%;随喷施溶液质量浓度上升,三碘苯甲酸处理的幼苗光合与非光合组织生物量比值呈现抛物线趋势,仅在T2处理下显著高于CK处理,相较于CK处理提高了157%;随喷施溶液质量浓度上升,芸苔素内酯处理的幼苗光合与非光合组织生物量比值呈现先增后减的趋势,在B3处理下达到最高,相较于CK处理提高了148.3%。
总体上,3种植物生长调节剂不同程度地影响了青钱柳幼苗的光合与非光合组织生物量分配,进而影响了其光合效率。相较于矮壮素,三碘苯甲酸(50、100、150mg/L)和芸苔素内酯(0.05、0.10、0.15mg/L),在提高幼苗光合与非光合组织生物量比值方面的效果更加显著且稳定。
2.4植物生长调节剂对幼苗叶片总黄酮含量的影响
由图6可知,3种植物生长调节剂均能提高幼苗叶片的黄酮含量。矮壮素处理的幼苗叶片总黄酮含量随矮壮素质量浓度上升呈现先增后减的趋势,且均显著高于CK处理,在C3处理达到最高,相较于CK处理提高了33.2%;三碘苯甲酸和芸苔素内酯处理的幼苗叶片总黄酮含量随喷施溶液质量浓度上升呈现波动趋势,分别在T2和B4处理下达到最高,相较于CK处理分别增加了34.9%和30.3%。
总体上,3种植物生长调节剂均在一定程度上提高了青钱柳幼苗叶片中的总黄酮含量,尽管影响存在差异,但整体来看3种植物生长调节剂对叶片总黄酮含量的提升效果相近。
2.5植物生长调节剂对幼苗PAL和CHS活性的影响
由图7可知,除T3处理外,所有植物生长调节剂处理的幼苗叶片PAL活性均显著高于CK处理。矮壮素处理的幼苗叶片PAL活性随着浓度的增加呈先增后减的趋势,在C3处理达到最高,相较于CK处理增加172.3%;三碘苯甲酸和芸苔素内酯处理的幼苗叶片PAL活性随浓度上升呈现递增和波动趋势,分别在T4和B2处理达到最高,相较于CK处理分别增加125.2%和93.3%。
如图8所示,3种植物生长调节剂处理的幼苗叶片CHS活性均显著高于CK处理。矮壮素处理中C1处理幼苗CHS活性最高,相较于CK处理增加230.1%。三碘苯甲酸处理的幼苗叶片CHS活性随着喷施溶液质量浓度的增加呈现先增后减的趋势,在T2处理达最大值,相较于CK处理提高了194.1%。芸苔素内酯处理的幼苗叶片CHS活性随着喷施溶液质量浓度的增加呈现递增趋势,在B4处理达最大值,相较于CK处理提高了310.3%。在所有处理中,B4处理的幼苗叶片CHS活性显著高于其他处理。
总体上,3种植物生长调节剂均在一定程度上提高了青钱柳幼苗叶片的PAL和CHS活性,进而促进了黄酮的合成。矮壮素处理提升PAL活性的效果最为显著,芸苔素内酯处理提升CHS活性的效果最为显著。
2.6植物生长调节剂对幼苗光合积累和黄酮合成影响的综合评价
对叶绿素含量、SSC含量、SC含量、NSC含量、光合与非光合组织生物量比值、总黄酮含量,PAL活性及CHS活性这8个指标进行主成分分析和KMO、Bartlett球形检验。结果表明,指标间存在强相关性(表2),第1主成分的方差解释率达92.499%,特征根为12.025,可以用单一主成分解释。
利用主成分1载荷系数和特征根计算综合得分系数(表3)。按照综合得分系数由高到低排序,各处理依次为T3、C1、B1、T4、T1、C2、T2、C3、B2、C4、B3、CK、B4。T3处理(150mg/L三碘苯甲酸溶液)促进青钱柳幼苗光合积累和黄酮合成的效果最好,其次是C1处理(500mg/L矮壮素溶液)、B1处理(0.05mg/L芸苔素内酯溶液)。总体来看,不同浓度的三碘苯甲酸处理对于幼苗的光合积累和黄酮合成有较强的正面影响。
3讨论与结论
光合作用是植物生长发育的基础,叶绿素作为光合作用中的关键色素,其含量直接影响到光能的吸收和转化效率[19-20]。本研究结果表明,C1处理(500mg/L矮壮素溶液)显著提升了青钱柳幼苗叶片的叶绿素含量。这一结果与马亚男等[21]关于番茄的研究结果相一致。叶绿素含量的提高增强了植物对光能的利用效率,加快了光合作用进程,也为植物提供了更加充足的同化物基础,从而促进了初级代谢产物的合成[22]。然而,初级代谢产物(如SSC、SC)的合成与叶绿素含量的提高并不呈现线性关系[23]。本研究中,矮壮素处理虽然促进了叶绿素积累,但是不利于碳水化合物的形成,这与徐志勇等[24]的研究结果相一致,即矮壮素通过抑制赤霉素合成,影响了植物的能量积累。矮壮素抑制赤霉素合成,打破了植物体内的激素平衡,进而诱导植物资源向细胞伸长和分生组织活动重新分配,用于碳水化合物积累的养分和能量减少[21]。相反,三碘苯甲酸T1和T3处理下碳水化合物的含量显著增加。特别是T1处理下SSC和NSC含量的提升,这与Li等[25]报道的三碘苯甲酸阻断了糖的代谢并促进糖分积累的结果相一致。同时,施用三碘苯甲酸可能激发植物的逆境响应机制,通过积累SSC来提高细胞的渗透压,增强其对逆境的适应性。施用不同浓度的芸苔素内酯对青钱柳幼苗能量积累的影响呈现波动趋势。在芸苔素内酯较低质量浓度的B1处理条件下,幼苗的糖分积累得以促进,这与Jiang等[26]关于黄瓜糖分累积的研究结果相一致。当芸苔素内酯质量浓度增加至B2处理水平时,幼苗的能量积累受到抑制。然而在更高质量浓度(B3、B4处理)的芸苔素内酯处理情况下,幼苗的能量积累再次被促进。这可能是因为在较高的质量浓度范围内,芸苔素内酯重新激发了相关的代谢过程,导致碳水化合物含量上升,形成了新的平衡点。
本研究中探讨了不同浓度的植物生长调节剂对青钱柳幼苗光合与非光合组织生物量比值及黄酮合成的影响。结果显示,除C4处理外,3种植物生长调节剂均在一定程度上提升了青钱柳幼苗光合与非光合组织生物量的比值,中间水平的植物生长调节剂溶液提升该比值的效果更加明显,这表明可以通过精确调控生长调节剂浓度来优化植物生长资源的分配,促进光合组织发展,进而提升光合效率与生长潜力[27-28]。
施用3种植物生长调节剂均提高了青钱柳叶片的总黄酮含量,其中T2处理(100mg/L三碘苯甲酸溶液)的效果最为显著,这与刘小锐等[29]关于罗布麻的研究结果相一致。同时,PAL和CHS这2种黄酮合成关键酶的活性得到显著增强。黄酮类化合物的合成与苯丙烷类合成途径紧密相关,PAL是苯丙烷类合成途径第一步反应的关键酶和限速酶,CHS是黄酮合成途径中的另一个关键酶,参与查尔酮的生成,进而促进各种黄酮类化合物的进一步合成[30]。这2种酶的活性增强直接促进了黄酮的生物合成,为本研究中施用生长调节剂使幼苗总黄酮含量上升的现象提供了合理的解释。此外,不同植物生长调节剂及不同浓度处理下,PAL和CHS活性的变化趋势不同,如在C3处理下PAL活性达到最高,在B4处理下CHS活性显著高于其他处理。这种差异性表明,每种植物生长调节剂及其特定浓度可能通过不同的分子机制作用于黄酮合成途径的特定阶段,从而影响最终的黄酮产量以及植物的防御响应[31]。综合来看,可以推断植物生长调节剂通过激活PAL和CHS等关键酶,促进了黄酮合成途径中前体物质的转化与下游黄酮化合物的积累。
主成分分析结果揭示了一系列生理生化指标间的高度相关性,说明所选指标与青钱柳幼苗光合积累与黄酮合成过程紧密关联,且单个主成分解释了大部分变异(92.499%)。结果显示,T3处理(150mg/L三碘苯甲酸溶液)对青钱柳幼苗的光合作用和黄酮合成具有最显著的促进作用,这与Qi等[32]的研究结果相似,即三碘苯甲酸通过抑制生长素的极性运输,使更多能量被用于糖分积累与次生代谢产物的合成,同时促进侧根的形成,增强了植物吸收水和养分的能力,进而提升了干物质积累量和总黄酮含量。矮壮素处理提高了青钱柳幼苗叶片的叶绿素与黄酮含量,提升叶绿素含量的效果非常显著,但其会抑制青钱柳幼苗的能量积累,在生产中可根据需求使用。B1处理(0.05mg/L芸苔素内酯溶液)虽然对青钱柳幼苗光合积累及黄酮合成显示出一定的促进效果,但是其综合效果不如三碘苯甲酸和矮壮素显著。作为一种新型植物生长调节物质,芸苔素内酯可以调节植物体内多种内源激素的平衡,提高植物细胞内的叶绿素含量和CO2固定速率,进而提高植物的光合作用效率,并影响次生代谢产物的合成[33]。然而,本研究中芸苔素内酯在促进青钱柳幼苗光合积累及黄酮合成方面未表现出稳定的正面效应,可能是由于青钱柳对芸苔素内酯的响应模式不同于其他植物,其他浓度处理与施用方法等的效果有待进一步研究。
矮壮素、三碘苯甲酸和芸苔素内酯对青钱柳幼苗光合积累及黄酮合成均有一定的调控作用,且大多为促进作用,其中150mg/L三碘苯甲酸溶液的促进效果最好,其次为500mg/L矮壮素溶液和0.05mg/L芸苔素内酯溶液。综合来看,不同质量浓度的三碘苯甲酸溶液均对青钱柳幼苗的光合积累和黄酮合成产生了较为明显的正面影响,更适合应用于青钱柳的矮化栽培。本研究中仅使用3种植物生长调节剂的4种质量浓度梯度进行试验,试验设计存在不足,如低于500mg/L的矮壮素溶液或高于200mg/L的三碘苯甲酸溶液的处理效果有待进一步研究,后续研究中应增加植物生长调节剂施用种类,同时通过更精细的梯度试验确定适宜青钱柳幼苗的最佳质量浓度,综合研究其对叶绿素含量、碳水化合物代谢以及其他药用成分(如三萜和多糖等)合成的促进作用。
