山桐子芽叶果可溶性糖和可溶性蛋白含量与其大小年的关系
2024-12-27耿晓东张召增朱秀征蔡齐飞李志王艳梅刘震




关键词:山桐子;可溶性糖;可溶性蛋白;枝条类型;大小年结实类型
山桐子Idesiapolycarpa是大风子科Flacourtiaceae山桐子属Idesia落叶乔木,果实含油率30%左右,果实油不饱和脂肪酸含量高达90%,其中亚油酸含量在65%~80%,是重要的新兴木本食用油料树种[1]。其树干通直,树冠分层,花淡黄色且芳香,秋季红果满冠,具有较高的观赏价值,被誉为“美丽的树上油库”[2]。盛果期山桐子单株产果量可达70kg以上,但由于山桐子是雌雄异株,雌株还存在大小年现象,影响着山桐子的丰产性[3]。解决山桐子雌株大小年的问题实现稳产高产具有重要的现实意义。
大小年现象在许多树木中普遍存在[4-7]。开心果Pistaciavera大小年现象明显,大年夏季花芽脱落是翌年减产为小年的直接原因[8-13]。开心果枝条果实发育和花芽分化间存在着营养物质(主要是糖)的竞争,大年导致花芽分化营养不良诱发细胞程序性死亡和早噬而早落,致使第2年小年的发生[10,14]。柑橘Citrusclementina×Citrusunshiu×Citrusnobilis大年以牺牲花芽为代价,使果实积累更多的糖类,获得更多的果实,进而影响下一年营养枝的发育和开花数量,导致小年的出现[15-16]。油橄榄Oleaeuropaea也存在相似的营养调控[17]。“抱子怀胎”的油茶Camelliaoleifera大年结果较多,用于果实发育的营养消耗较多,导致花芽分化减少,甚至没有花芽,形成的花芽也会脱落,隔年就会出现结果数量锐减的现象,成为小年[4,18]。油茶花芽数量与当年春梢数量呈现正相关,与当年的植株产果量、坐果率呈现负相关[19];外施赤霉素和多效唑不仅显著提高了‘湘林27’和‘湘林14’油茶的花芽分化率、花芽饱满度和连续2年果实产量,还增加了不同节位叶片制造的同化物在花芽和果实中的分配量,缓解了油茶的大小年现象[20]。可溶性糖和可溶性蛋白作为营养物质,在树木花芽分化中起着至关重要的作用。在植物成花诱导期,糖类与花芽形成具有正相关性。油桐Verniciafordii的花芽分化可能与蛋白质的积累和消耗有着密切的关系[21];文冠果Xanthocerassorbifolium的芽和叶片营养的相关性最为密切,上部芽的磷和可溶性糖含量与叶片的营养状态有显著相关性,雌芽的分化需要消耗大量的可溶性糖[22];在芽期和花期喷施3%的蔗糖溶液能使元宝枫Acertruncatum果实可溶性糖和蛋白质含量增加,能明显提高元宝枫果实品质[23]。大小年现象的发生在许多经济林树种中的表现不尽相同,但本质上均是大量开花结实对营养的消耗影响到花芽分化(或脱落)造成的。
由于山桐子种子靠鸟传播,地理隔绝有限,在横跨亚热带与暖温带分布区内基因得以广泛交流,致使种源内变异丰富,分化出了不同的大小年类型,同一种源山桐子也有大小年现象发生。目前,山桐子相关研究主要包括营养生长与生殖生长模式[24-25],不同种源开花结实的生理响应[26],果实发育过程中内源激素的动态变化[27],性别分化的表型、解剖构造变化以及转录组和蛋白组分析[28-29],内源激素与大小年的关系[3]等。但山桐子大小年与营养的关系尚不明晰,探讨山桐子大小年形成的营养调控机制具有重要的理论价值。
文中以成都种源7年生山桐子为研究对象,通过对比观测不同大小年类型山桐子结果枝与不结果枝上的叶片、芽和果实的可溶性糖和可溶性蛋白含量的差异,以期探寻山桐子大小年形成的营养机制,为解决山桐子在生产上存在的大小年问题提供理论依据。
1材料与方法
1.1材料
供试材料为种植于河南农业大学林学试验站(113°42′E,34°43′N)内的7年生成都种源山桐子,栽植行距为2m×4m,树势生长良好,已进入稳定结果期,试验期间水肥等管理方式相同。试验地属于暖温带大陆性季风气候,四季分明,年均温14.2℃,年平均降水量650.1mm,无霜期220d,土壤质地为砂壤土,pH7.5,有机质含量7.4g/kg,有效氮含量0.56mg/kg,有效磷含量1.31mg/kg,有效钾含量20.8mg/kg。
1.2方法
根据前期对山桐子开花结实量的调查,把山桐子结实大小年划分为3类:小年产量占大年产量80%以上的无大小年(NAB)、小年产量低于大年产量80%的不完全大小年(IAB)和小年不结实的完全大小年(CAB)。选取不同大小年类型山桐子标准木各3株,分别在2021年果实膨大期(5月30日)、油脂积累期(7月10日)、花芽分化期(7月30日)和果实成熟期(10月10日)采摘分布于树冠中部南向有果枝条和无果枝条上着生的叶片、芽和果实各5个,用液氮速冻后置于-80℃超低温冰箱中,用于可溶性糖含量和可溶性蛋白含量的测定,测定方法分别为蒽酮比色法和考马斯亮蓝G-250染色法[30]。每次取样0.5g,重复3次。
1.3数据处理与分析
利用WPSOffice软件进行数据整理,使用Origin2017软件绘图。使用SPSS24.0软件对不同大小年类型山桐子营养指标进行单因素方差分析,若差异显著,则用邓肯多重比较法在α=0.05水平下进行多重比较,得出不同大小年类型山桐子以及不同类型嫩枝之间各指标的差异性。
2结果与分析
2.1不同大小年类型山桐子可溶性糖含量的差异
2.1.1叶片可溶性糖含量的差异
由表1可知:3种大小年类型山桐子的有果嫩枝和无果嫩枝的叶片可溶性糖含量随着果实发育呈现先降低后升高的趋势,并在花芽分化期达到最小值;同一发育时期内,无大小年山桐子叶片可溶性糖含量最大,不完全大小年样品次之,完全大小年样品该指标最小,在油脂积累期、花芽分化期和果实成熟期,无果嫩枝中三者之间的差异均达到了显著水平(P<0.05);在果实膨大期、油脂积累期和果实成熟期,山桐子有果嫩枝叶片可溶性糖含量显著高于无果嫩枝(P<0.05),但在花芽分化期,除不完全大小年有果嫩枝与无果嫩枝上的叶片可溶性糖含量差异达到显著水平外,其他2种类型差异不显著。
2.1.2芽可溶性糖含量的差异
由表2可知,无大小年山桐子芽可溶性糖含量随着果实发育呈现先升高后降低的趋势,在花芽分化期达到最大值,并达到显著水平(P<0.05)。而不完全大小年和完全大小年山桐子芽的可溶性糖含量随着果实发育呈现逐渐升高的趋势,但完全大小年样品在花芽分化期与果实成熟期之间,不完全大小年样品在果实膨大期与油脂积累期之间、花芽分化期与果实成熟期之间的差异未达显著水平(P>0.05)。相同发育时期内,无大小年山桐子芽可溶性糖含量最高,由高到低排序依次均为无大小年样品、不完全大小年样品、完全大小年样品。除油脂积累期完全大小年无果嫩枝上芽的可溶性糖含量低于有果嫩枝外(P>0.05),其他时期及不同大小年类型均表现为无果嫩枝上芽的可溶性糖含量高于有果嫩枝,并达显著水平(P<0.05)。
2.1.3果实可溶性糖含量的差异
从图1可以看出,山桐子果实可溶性糖含量随着果实发育呈现一直升高的趋势,并在果实成熟期达到最大值;同一发育时期内,完全大小年山桐子果实可溶性糖含量最大,不完全大小年样品次之,无大小年样品最小,在花芽分化期和果实成熟期,三者之间的差异均达到了显著水平,完全大小年山桐子的果实可溶性糖含量显著高于其他2种大小年类型的山桐子。
2.2不同大小年类型山桐子可溶性蛋白含量的差异
2.2.1叶片可溶性蛋白含量的差异
由表3可知,3种大小年类型山桐子有果嫩枝上的叶片可溶性蛋白含量随着果实发育呈现先降低后升高的趋势,并在花芽分化期达到最小值。无大小年和不完全大小年类型有果嫩枝上的叶片可溶性蛋白含量的差异达到显著水平,但从油脂积累期到果实成熟期,完全大小年类型的叶片可溶性蛋白含量的差异未达显著水平,且含量较低。
3种大小年类型山桐子无果嫩枝上的叶片可溶性蛋白含量随着果实发育的变化趋势不一致。无大小年类型是随着果实发育呈现逐渐增加的趋势,但果实膨大期与油脂积累期的含量差异未达显著水平;不完全大小年类型是果实膨大期和油脂积累期的叶片可溶性蛋白含量大于花芽分化期和果实成熟期;完全大小年类型随着果实发育呈现逐渐降低的趋势,除花芽分化期与果实成熟期的含量差异不显著外,其他时期的含量差异达到显著水平。
通过比较同一时期3种大小年类型叶片可溶性蛋白含量的差异发现,不完全大小年类型样品的含量最高,除果实膨大期完全大小年类型样品含量次之,无大小年类型样品含量最小外,其他时期均是无大小年类型样品含量次之,完全大小年类型样品含量最小。除了果实膨大期不完全大小年类型无果嫩枝和完全大小年类型间、油脂积累期无大小年类型无果嫩枝与完全大小年类型间、花芽分化期无大小年类型有果嫩枝与完全大小年类型间、果实成熟期无大小年类型有果嫩枝与不完全大小年类型间的含量差异不显著外,其他时期样品含量的差异均达到了显著水平。
通过比较有果嫩枝和无果嫩枝的含量差异可以看出,无果嫩枝叶片可溶性蛋白含量均高于有果嫩枝,除果实膨大期完全大小年类型和不完全大小年类型,果实成熟期无大小年类型的有果枝条与无果枝条间的可溶性蛋白含量差异未达显著水平外,其他时期样品可溶性蛋白含量均表现为差异显著。
2.2.2芽可溶性蛋白含量的差异
由表4可知,3种大小年类型山桐子的有果嫩枝和无果嫩枝的芽可溶性蛋白含量随着果实发育呈现一直升高的趋势,并在果实成熟期达到最大值。在同一发育期内,无大小年类型山桐子嫩枝上的芽可溶性蛋白含量最大,不完全大小年类型次之,完全大小年类型最小,含量差异达到显著水平(P<0.05),无果嫩枝上的芽可溶性蛋白含量显著高于有果嫩枝(P<0.05)。
2.2.3果实可溶性蛋白含量的差异
从图2可以看出,3种大小年类型山桐子果实可溶性蛋白含量随着果实发育呈现一直升高的趋势,并在果实成熟期达到最大值。完全大小年类型山桐子的果实可溶性蛋白含量最大,不完全大小年类型含量次之,无大小年类型含量最小。在果实膨大期、油脂积累期和果实成熟期,三者之间的差异均达到了显著水平(P<0.05)。在花芽分化期和果实成熟期,完全大小年类型山桐子的果实可溶性蛋白含量显著高于其他2种大小年类型的山桐子(P<0.05)。
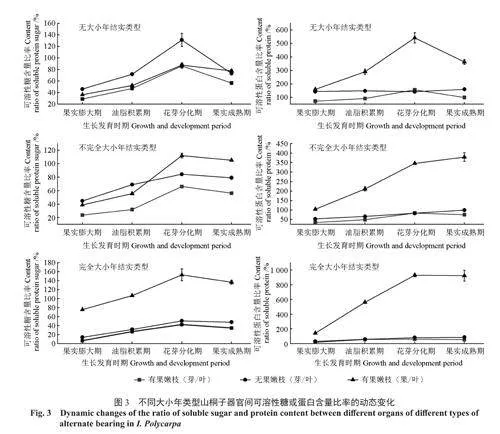
2.3不同大小年类型山桐子器官间可溶性糖和可溶性蛋白分配的变化
2.3.1器官间可溶性糖或可溶性蛋白含量比率的变化
由图3可以看出,山桐子的芽、果可溶性糖含量与叶可溶性糖含量的比率均随着果实发育而升高,到花芽分化期达到最高值,随后有所降低,其中无大小年类型芽与叶可溶性糖含量的比率下降最多。同一发育期内,无大小年类型芽与叶可溶性糖含量比率最高,不完全大小年类型次之,完全大小年类型最小;而果实与叶可溶性糖含量比率相反,完全大小年类型最高,不完全大小年类型次之,无大小年类型最小。另外,有果枝上芽与叶可溶性糖含量的比率均小于同期的无果枝,说明芽与果实对可溶性糖的竞争决定着大小年的发生。
由图3还可以看出,山桐子的芽与果以及芽与叶的可溶性蛋白含量的比率随着果实发育变化不明显,未达显著水平(P<0.05)。而果实与叶的可溶性蛋白含量比率均随着果实发育而升高,在花芽分化期达到最大值,随后无大小年类型该指标下降明显,而不完全大小年类型和完全大小年类型的该指标维持在较高的水平,变化不明显。同一发育期内,完全大小年类型山桐子果实与叶片的可溶性蛋白含量比率最高,无大小年类型次之,不完全大小年类型最小。说明芽与果实对可溶性蛋白也存在着竞争关系,完全大小年类型的山桐子果实对可溶性蛋白的竞争力最强。
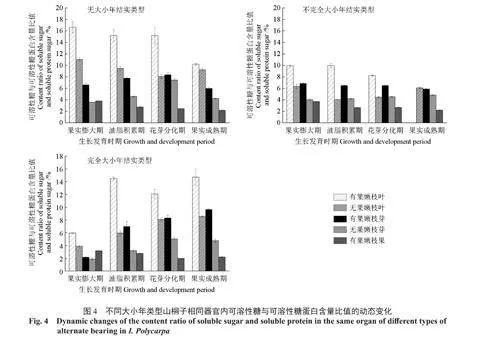
2.3.2器官间可溶性糖与可溶性蛋白含量比值的变化
由图4可以看出,有果嫩枝叶片和芽的可溶性糖含量与可溶性蛋白含量的比值大于无果嫩枝叶片和芽,叶片的可溶性糖含量与可溶性蛋白含量的比值大于芽,果实的可溶性糖含量与可溶性蛋白含量的比值较小。叶片为光合器官,光合初级产物糖类在叶内储存量较高,而芽和果实在发育过程中部分可溶性糖转化为可溶性蛋白,可溶性糖和可溶性蛋白的含量比值降低。无大小年类型和不完全大小年类型山桐子叶片可溶性糖与可溶性蛋白含量的比值随着果实发育有逐渐降低的趋势,而完全大小年类型该比值有逐渐升高的趋势;无大小年和完全大小年类型山桐子芽的该比值随着果实发育先升高,在花芽分化期达到最大,随后又有所降低,而不完全大小年类型该比值的变化不明显;3种大小年类型山桐子果实的该比值均呈现逐渐降低的趋势,在果实成熟期该比值最小。通过比较山桐子叶片和芽的可溶性糖与可溶性蛋白含量的比值,发现果实膨大期有果嫩枝叶3种大小年类型按照可溶性糖与可溶性蛋白含量的比值由高到低排序依次为NAB(16.64)、IAB(9.93)、CAB(5.95),随着果实发育,各大小年类型间的差异变小。说明在果实膨大期(即5月底—6月初)叶片和芽(尤其是叶片)的可溶性糖和可溶性蛋白含量的比值,可能影响着结实大小年的发生。提高果实膨大期叶片的光合产物量,提高叶片可溶性糖与可溶性蛋白含量的比值,对于防止小年发生可能具有重要的意义。
3讨论与结论
山桐子有果枝芽可溶性糖和可溶性蛋白含量显著低于无果枝,不利于花芽分化,说明山桐子同一枝条无法实现连年结果是果实发育与花芽分化竞争营养物质所导致,这与较多其他树种的相关研究结果一致[4,16,18,25]。与营养不足致使开心果花芽脱落不同[8,13],山桐子大小年的发生是营养不足导致花芽分化受限所致。花芽分化不能顺利进行是大小年现象发生的关键性因素[31],营养物质的积累则是影响花芽分化的一种重要因素[32-33]。保证营养供应,促进结果枝花芽分化,是控制山桐子大小年发生的关键。
除花芽分化期差异未达显著水平外,有果嫩枝叶片可溶性糖含量显著大于无果枝叶片,但有果枝芽可溶性糖含量、叶片以及芽可溶性蛋白含量均小于无果枝,说明有果枝叶片对可溶性糖的竞争强于无果枝,以供果实发育的需要,而无果枝叶片及芽可溶性蛋白含量增加,可以保证叶芽向花芽转化的营养供应,促进其花芽分化。同时,在花芽分化期叶片可溶性糖和可溶性蛋白含量最低,但芽的可溶性糖和可溶性蛋白含量最高的为无大小年类型样品,不完全大小年类型样品次之,完全大小年类型样品的含量最低,说明叶片向芽输送营养物质的量决定着大小年的发生。山桐子花芽分化期到果实成熟期,叶片的可溶性糖和可溶性蛋白含量决定着芽的可溶性糖和可溶性蛋白含量,进而决定着花芽分化的进行,最终决定着山桐子结实大小年的发生。所以,保障花芽分化期(7月底)可溶性糖和可溶性蛋白的供应是保障山桐子连年丰产的关键。油茶叶果比的差异并未导致叶片可溶性糖含量差异达显著水平[34],开心果、柑橘、杧果、小檗、葡萄等枝条发育和花芽分化间主要存在着可溶性糖营养物质的竞争[7,10,14,35-36],这些报道与本研究结果不完全一致。可溶性蛋白也是山桐子果实发育和花芽分化竞争的主要物质,这与越南安息香Styraxtonkinensis越冬枝条储藏的可溶性糖和可溶性蛋白含量高则促进其开花的研究结果是一致的[37],也与枇杷Eriobotryajaponica成花诱导要经历叶片可溶性蛋白质含量有一定程度升高的过程一致[38]。
从果实发育时期来看,在花芽分化期之前,有果枝叶片可溶性糖和可溶性蛋白含量逐渐降低,而后增加,但完全大小年类型在油脂积累期到果实成熟期变化未达显著水平,芽的可溶性糖含量在花芽分化期后也未明显增加,说明完全大小年类型的有果枝叶片可溶性糖和可溶性蛋白含量未能明显增加,导致叶片光合产物供应不足,花芽分化受阻。无果枝叶片可溶性糖含量变化趋势与有果枝相同,但叶片可溶性蛋白含量变化在不同大小年类型样品间差异显著。完全大小年类型山桐子叶片可溶性蛋白含量从果实膨大期到花芽分化期逐渐降低,而后维持在较低的水平;不完全大小年类型山桐子叶片可溶性蛋白含量在花芽分化期之前含量最高,在花芽分化期后含量降低,之后仍维持在较高的水平;无大小年类型山桐子叶片可溶性蛋白含量逐渐增加,且花芽分化期后其无果枝的可溶性蛋白含量与其他3个时期的差异达到显著水平。随着果实发育,有果枝及无果枝芽的可溶性糖和可溶性蛋白含量均逐渐增加。到花芽分化期,无大小年类型可溶性糖含量达到最大,随后有所降低,不完全大小年类型可溶性糖含量继续增加,完全大小年类型可溶性糖含量维持在最低的水平不变,而可溶性蛋白含量均是逐渐增加的趋势。上述结果说明,决定大小年的关键因素是芽的可溶性糖和可溶性蛋白含量,尤其在花芽分化期之后可溶性糖和蛋白含量的持续增加或保持较高水平至关重要。同时,无大小年类型山桐子在果实膨大期叶片可溶性糖与可溶性蛋白含量的比值最大,说明果实膨大期叶片营养状况也决定着大小年的发生,这与果实膨大期的叶和芽内ABA与GA3含量比值以及ZR、ABA总含量与GA3含量比值越大越有利于花芽分化[3]的结果一致。果实膨大期的内源激素状态和叶片营养状况可能决定着花芽分化期叶片的可溶性糖和可溶性蛋白含量[39]。上述结果说明5月底的果实膨大期也可能是花芽生理分化期,7月下旬达到花芽形态分化期。从山桐子叶芽生长发育规律来看,5月底山桐子高生长进入缓慢生长期,随后进入腋芽集中萌发形成近轮生枝期[40-41],该时期叶片营养与内源激素状况可能也决定着当年的花芽分化,进而决定着翌年的开花结实。
从果实的可溶性糖和可溶性蛋白含量来看,无大小年类型果实可溶性糖和可溶性蛋白含量最低,不完全大小年类型次之,完全大小年类型最高,尤其是到花芽分化期后完全大小年类型的含量显著高于不完全大小年类型和无大小年类型。该结果说明有大小年的山桐子果实营养含量要高于无大小年的山桐子。如果连续2年内有大小年的果实绝对产量大于无大小年的绝对产量,保障单位面积内有一定数量的结果植株,不一定所有单株均结实,也可以实现连年丰产。唐勇[42]报道了隔年交替结果模式可以提高荔枝产量、果实品质,实现省力化栽培,山桐子的隔年结果模式是否可行有待进一步验证。
综上,山桐子的果实发育与花芽分化间存在着竞争关系,有果枝的花芽分化受到果实发育的抑制,而无果枝易于花芽分化;果实膨大期叶片可溶性糖与可溶性蛋白含量的比值越高,越能保证花芽分化期之后芽内可溶性糖和可溶性蛋白含量的持续增加,保证花芽分化,促进翌年开花结实,防止小年的发生。
