不同茶树品种叶片细胞壁氟富集差异探究
2024-11-08刘昱杨培迪张培凯詹文礼李游姚苏航赵洋成杨刘振沈程文


摘要:茶树是一种氟超富集植物,不同茶树品种的叶片富集氟含量存在差异。为探究茶树叶片细胞壁氟富集规律,检测了15个茶树品种的叶片氟含量,以及叶片果胶、半纤维素、纤维素与其对应的氟含量,分析不同成熟度叶片细胞壁氟分布。结果显示,果胶是体现品种细胞壁氟富集成熟度差异的关键细胞壁组分。15个品种中,涟茶7号、槠叶齐、上梅洲种的成熟叶果胶含量与果胶氟含量都比嫩叶高,而铁观音是唯一成熟叶比嫩叶中果胶氟含量低的品种。细胞壁组分氟占比的相关性分析表明,茶树叶片成熟度越高,氟含量越高,且均呈细胞壁多个组分富集氟的趋势。通过综合评价及聚类分析筛选,茶树品种寒绿成熟叶组和嫩叶组均具有高细胞壁氟富集水平(D1-4=0.704 6,D5-8=0.928 6);福鼎大白茶、湘波绿、湘波绿2号、碧云、黔湄702具有中等细胞壁氟富集水平(D1-4=0.326 7~0.486 1,D5-8=0.484 4~0.699 3);云南乌黑大叶、槠叶齐具有低细胞壁氟富集水平(D1-4=0.146 5~0.268 8,D5-8=0.222 3~0.345 7)。本研究从品种和叶片成熟度角度探究茶树细胞壁氟富集规律及其差异性,为低氟茶树分子育种提供品种参考。
关键词:茶树品种;氟;叶片细胞壁;成熟度
中图分类号:S571.1 文献标识码:A 文章编号:1000-369X(2024)05-735-12
Investigation of Differential Fluorine Enrichment in Leaf Cell Walls of Different Tea Cultivars
LIU Yu, YANG Peidi, ZHANG Peikai, ZHAN Wenli, LI You, YAO Suhang, ZHAO Yang,
CHENG Yang, LIU Zhen*, SHEN Chengwen*
1. Key Lab of Education Ministry of Hunan Agricultural University for Tea Science, Changsha 410128, China ;
2. Tea Research Institute, Hunan Academy of Agricultural Sciences, Changsha 410125, China
Abstract: Tea plants are known to hyperaccumulate fluorine, with significant variation in fluorine accumulation among different cultivars. To explore the patterns of fluorine accumulation in tea leaf cell walls, the fluorine content in leaves of 15 tea cultivars was measured. Moreover, the contents of pectin, hemicellulose, cellulose components, and their respective fluorine contents were analyzed. The distribution of fluorine in the cell walls at different maturity stages of leaves were examined. The results indicate that pectin is a crucial component reflecting the differences in fluoride accumulation and maturity level of the cell walls among cultivars. Only the mature leaves of ‘Liancha 7’, ‘Zhuyeqi’ and ‘Shangmeizhouzhong’ increased pectin content and fluoride content in pectin compared to the young leaves. ‘Tieguanyin’ was the only cultivar with the fluoride content in pectin decreased in the mature leaves compared to the young leaves. Correlation analysis of the fluoride proportion in the cell wall components reveals that higher fluoride content in the leaves correlates with greater maturity, reflecting a trend of multi-component fluoride accumulation in the cell walls. Through a comprehensive evaluation and cluster analysis, ‘Hanlü’ exhibits consistently high levels of fluorine accumulation in cell walls at different maturity stages (D1-4=0.704 6, D5-8=0.928 6). ‘Fuding Dabaicha’, ‘Xiangbolü’, ‘Xiangbolü 2’, ‘Biyun’, and ‘Qianmei 702’ show moderate levels of fluorine enrichment in cell walls (D1-4=0.3267-0.4861, D5-8=0.484 4~0.699 3), while ‘Yunnan Wuheidaye’ and ‘Zhuyeqi’ exhibit low levels of fluorine accumulation (D1-4=0.146 5-0.268 8, D5-8=0.2223-0.345 7). This study explored the regularity and difference of fluorine accumulation in tea cell walls from the perspectives of cultivar and leaf maturity, and provided insights for molecular breeding of low-fluorine tea cultivars.
Keywords: tea cultivar, fluorine, leaf cell wall, maturity
黑茶是广受欢迎的茶类饮品,然而其粗老原料中氟含量过高,已成为了阻碍黑茶产业发展的重要问题。与其他植物相比,茶树具有氟超富集的特性,在自然条件下叶片能积累大量的氟(871~1 337 mg·kg-1)而不出现毒害症状[1]。研究表明,茶树叶片细胞壁氟含量可占茶树叶片总氟含量的55.45%~80.49%,是富集氟的主要部位[2]。植物细胞壁是植物细胞最外层包围结构,由多糖、糖蛋白、木质素等生物大分子构成,具有连接细胞并维持细胞形态,参与胞内外离子稳态调控,信号传递及胁迫响应等功能[3]。质外体途径运输的氟离子进入茶树细胞首先由细胞壁螯合固定,因此细胞壁是氟进入茶树叶片中固定富集的第一道屏障和“仓库”。果胶(Pectin)、半纤维素(Hemicellulose)和纤维素(Cellulose)三大类多糖构成细胞壁90%的物质总量,其中果胶是一类富含半乳糖醛酸的杂多糖,含有的多种游离官能团,如-COO-和-NH2,能与Ca、Al金属离子结合带正电荷,进而结合氟离子[4]。现有研究认为,果胶是茶树叶片细胞壁结合氟的主要组分。刘思怡等[5]研究表明,福云6号成熟叶细胞壁中,果胶结合的氟占细胞壁总氟比例最高(55.44%),其次为半纤维素(36.60%),最低为纤维素(7.96%)。
茶树富集氟的特性与茶树品种密切相关。朱晓静等[6]测定相同生长环境条件下嘉茗1号、梅占、迎霜、福安大白茶、福鼎大白茶、福云6号的一芽三叶氟含量,发现叶片氟含量最高的是梅占(192.50 mg·kg-1),最低的是福鼎大白茶(132.50 mg·kg-1)。Yang等[7]将福云6号和嘉茗1号茶树品种的一年生茶树幼苗在氟的质量浓度为4 mg·L-1营养液中培养4周后,按照一芽二叶采摘,测定叶片氟含量,发现福云六号氟含量为168.22 mg·kg-1,显著低于嘉茗1号氟含量(246.56 mg·kg-1)。陈瑞鸿等[8]测定了薮北、嘉茗1号、水古、龙井43等茶树品种的夏梢第6叶的氟含量,发现龙井43品种的氟含量最高(1 278.50 mg·kg-1),水古品种的氟含量最低(692.90 mg·kg-1)。蔡荟梅等[9]对平阳特早茶、嘉茗1号和凫早2号一年生幼苗进行50 mg·L-1氟浓度水培试验,测定其成熟叶细胞壁氟含量,发现平阳特早茶为588.99 mg·kg-1,显著高于凫早2号(298.50 mg·kg-1)。目前,由于不同研究选取的品种和叶片成熟度不同,很难进行统一对比。因此,将茶树新梢按不同成熟度分为嫩叶组和成熟叶组分别考察,对研究不同品种氟富集差异变化十分必要。
以往对茶树细胞壁氟富集的研究多集中在少数茶树品种氟处理后的亚细胞分布差异,因品种和叶片成熟度不同,很难对不同研究结果进行统一对比,不同茶树品种新梢叶片氟富集差异规律还有待明确。本研究在前期基础上选择15个茶树品种,以秋季新梢为材料,叶片分为嫩叶组(1~4叶)、成熟叶组(5~8叶)。采用AIS细胞壁分离法等探究叶片细胞壁氟富集差异,以期明确不同品种茶树在细胞壁组分上的氟富集规律,并为进一步研究高氟品种与低氟品种细胞壁氟富集差异机理提供参考。
1 材料与方法
1.1 试验材料
参试的15个茶树品种(树龄为20 a)定植于湖南省农业科学院茶叶研究所高桥试验基地,分别为云南乌黑大叶(YNWHDY)、槠叶齐9号(ZYQ9H)、槠叶齐(ZYQ)、涟茶7号(LC7H)、铁观音(TGY)、盘龙山福引(PLSFY)、湘波绿2号(XBL2H)、湘波绿(XBL)、福鼎大白茶(FDDB)、黔湄702(QM702)、黔湄303(QM303)、湘安17号(XA17H)、碧云(BY)、寒绿(HL)、上梅洲种(SMZ)。2023年5月,对15个茶树品种进行深修剪,修剪深度为8~10 cm,原则为剪掉树冠上层的“鸡爪枝”。修剪后每亩(1亩≈667 m2)施200 kg菜籽饼和100 kg复合肥。于10月31日选取叶片成熟度一致的15个品种茶树新梢为试验材料。去除新梢茎部,叶片分为嫩叶组(1~4叶)、成熟叶组(5~8叶)。100 ℃蒸汽杀青,80 ℃烘干,打磨后过100目筛,于﹣20 ℃冰箱备用。
1.2 试验方法
1.2.1 叶片细胞壁分离
采用AIS法进行细胞壁分离,参考Renard等[10]稍作修改。称取5.000 g足干茶粉,加入50 mL 90%的预冷乙醇溶液匀浆,静置20 min后在4 ℃,5 000 r·min-1条件下离心10 min,弃上清液保留沉淀,此步骤重复3次。沉淀依次用冰丙酮、冰甲醇-三氯甲烷混合液(V冰甲醇∶
V三氯甲烷=1∶1)和冰甲醇各匀浆静置20 min洗涤1次(每次试剂用量为10 mL·g-1)。每次洗涤后,悬浊液在4 ℃下以5 000 r·min-1离心10 min,弃上清液保留沉淀。沉淀冷冻干燥即为叶片粗细胞壁,于﹣20 ℃冰箱保存备用。
1.2.2 细胞壁组分提取
使用分步提取法对茶树叶片细胞壁组分进行分离提取,参考Figueroa等[11]的方法并稍作修改。称取3.000 g足干叶片粗细胞壁,加入100 mL Na2CO3-CDTA混合提取液(提取液中Na2CO3的浓度为50 mmol·L-1,CDTA的浓度为20 mmol·L-1),在25 ℃、170 r·min-1条件下振荡提取24 h。4 ℃、2 000 r·min-1条件下离心并收集上清液,沉淀物用30 mL提取液洗涤离心1次,合并上清液,上清液冷冻干燥得到总果胶组分。沉淀干燥后继续加入100 mL NaOH-
NaBH4混合提取液(提取液中NaOH的浓度为2 mol·L-1,NaBH4的浓度为40 mmol·L-1),同样的步骤提取半纤维素,通过稀盐酸将上清液pH调整为6.0,冷冻干燥得到半纤维素组分。剩下沉淀残渣冷冻干燥后即为纤维素组分。
1.2.3 氟含量测定
全氟含量测定采用碱熔灰化前处理-氟离子电极法,参考朱晓静等[12]的方法并稍作修改。称取茶粉、细胞壁组分提取样品各0.1500 g(精确到0.000 1 g),置于30 mL镍坩埚中轻敲坩埚使样品分布均匀,加入3 mL NaOH溶液(8.375 mol·L-1),放入120 ℃烘箱中不带盖保持2 h后取出。将坩埚放入马弗炉,升温至300 ℃保持30 min,再升温至600 ℃熔融灰化60 min。冷却至室温取出,缓慢加入稀盐酸溶液(V浓盐酸∶V去离子水=1∶2)7 mL,冷却后转移到50 mL容量瓶。容量瓶加入3 mL溴甲酚紫酸碱指示剂。用1 mol·L-1 NaOH溶液滴定刚好变为紫红色,用稀盐酸溶液滴定液体刚好由紫红转为黄色(pH=6.0),纯水定容至50 mL。取25 mL待测液于50 mL烧杯中,加入25 mL TISAB缓冲液,搅拌均匀,氟离子选择电极测定。果胶、半纤维素组分氟含量以细胞壁组分提取所用叶片粗细胞壁质量表示:不同组分全氟含量/细胞壁组分提取称样量(3.000 g)。
1.2.4 细胞壁组分含量测定
样品采用足干茶粉。果胶含量以半乳糖醛酸含量表示,以D-(+)半乳糖醛酸作为标准品,采用硫酸咔唑法[13]于525 nm波长比色测定。半纤维素含量以木葡聚糖含量表示,以D-木糖作为标准品,采用二硝基水杨酸法[14]于540 nm波长比色测定,选用半纤维素含量检测试剂盒[索莱宝(北京)有限公司]。纤维素含量以β-D-葡萄糖含量表示,以葡萄糖作为标准品,采用蒽酮法[15]于620 nm波长比色测定,选用纤维素含量检测试剂盒[索莱宝(北京)有限公司]。所有比色测定均使用BioTekSynergy4多模式酶标仪(BioTek,Burlington,American)。
1.3 数据统计与分析
采用Excel 2016软件进行数据处理,SPSS 26.0软件与GraphPad Prism 9.5软件进行数据分析与画图。选择Bartlett’s test检验(P<0.05)进行单因素方差分析(One-way ANOVA),Tukey多重比较分析。通过SIMCA 14.1软件进行层次聚类分析。
1.3.1 隶属函数值
U(Xi)=(Xi-Xmin)/(Xmax-Xmin)······(1)
U(Xi)为第i个主成分综合指标的隶属函数值,Xi为第i个主成分综合值,Xmax、Xmin分别表示第i个主成分综合值中最大值及最小值。
1.3.2 权重系数
Wi=Pi/∑Pi································(2)
Wi为第i个主成分综合指标权重系数,Pi为主成分分析中第i个主成分综合指标的贡献率。
1.3.3 综合评价值
D=∑[U(Xi)×Wi] (3)
D为各品种综合评价值,得分越高表示该品种叶片细胞壁氟富集水平越高。
2 结果与分析
2.1 不同茶树品种叶片细胞壁组分与细胞壁组分氟含量差异分析
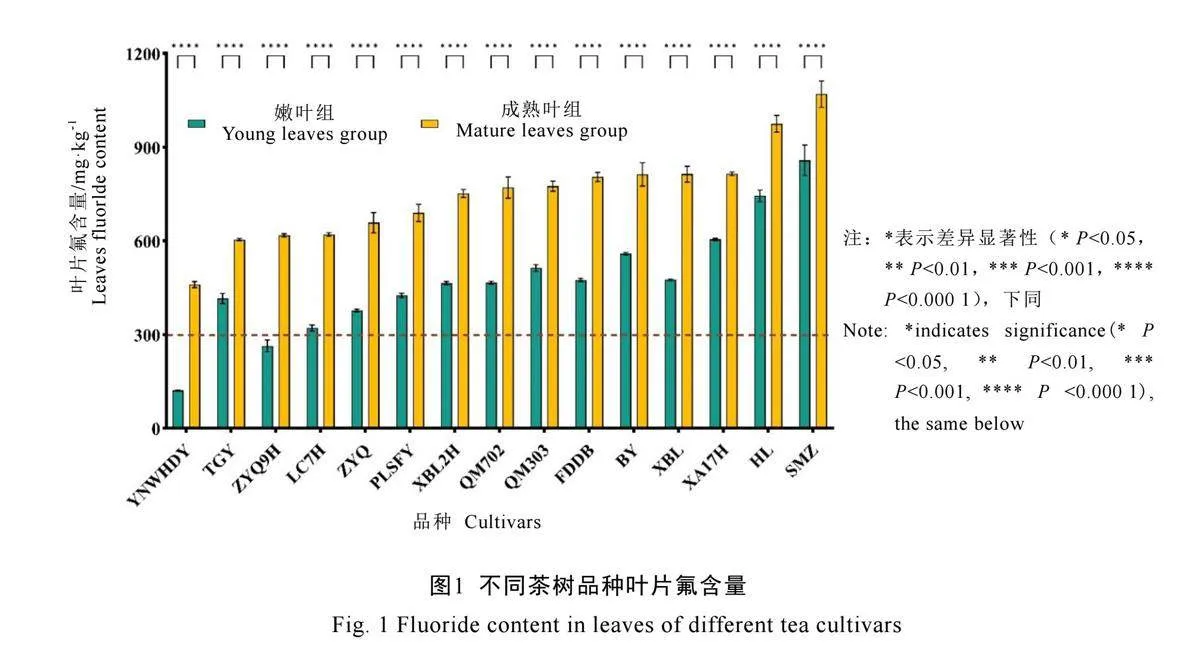
15个品种叶片氟含量检测结果见图1,结果显示各品种茶树两组叶片氟含量均差异极显著(P<0.000 1)。所选15个品种茶树嫩叶组氟含量范围为121.2~858.1 mg·kg-1,成熟叶组氟含量范围为460.4~1 070.2 mg·kg-1。两组氟含量最低品种均为云南乌黑大叶,最高均为上梅洲种。参考GB 19965—2005 砖茶氟含量的标准[16]规定最高含氟量300 mg·kg-1,仅云南乌黑大叶嫩叶组(121.2 mg·kg-1)、槠叶齐9号嫩叶组(264.2 mg·kg-1)符合标准。
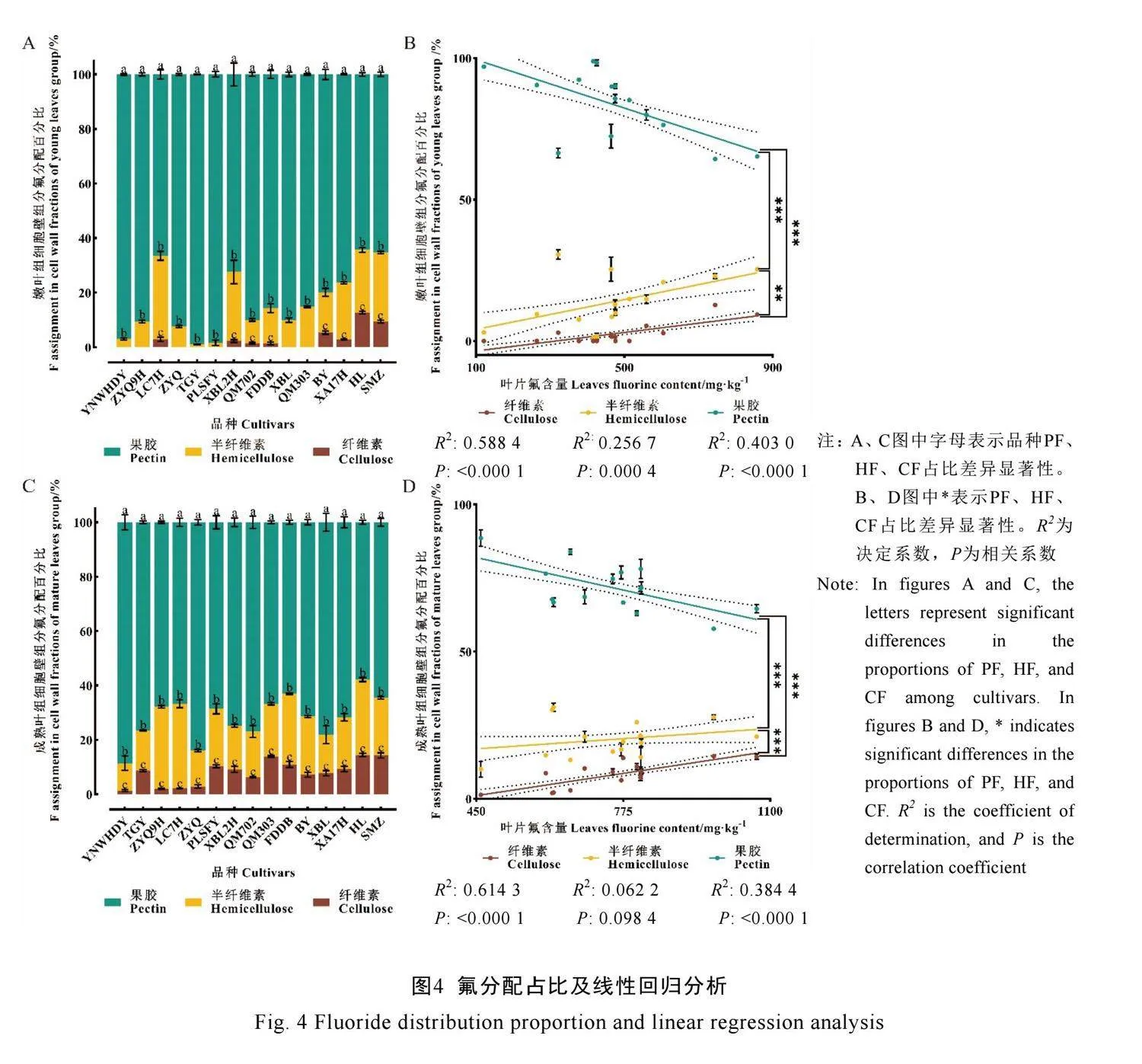
尽管各品种茶树成熟叶组叶片氟含量均高于嫩叶组且差异显著,但各品种细胞壁组分含量与细胞壁组分氟含量在嫩叶和成熟叶中是否存在差异需进一步探讨。对各指标进行单因素方差分析,图2A显示7个指标中,仅果胶含量两组差异不显著,表明各品种果胶含量在两组中可能存在差异变化。将不同品种的组分含量与组分氟含量进行差异关联分析。结果见图2B,槠叶齐9号果胶氟含量差异不显著,其余品种在两组间均差异显著;除铁观音外,成熟叶组比嫩叶组果胶氟含量高。云南乌黑大叶、盘龙山福引、湘波绿2号、福鼎大白茶、碧云果胶含量差异不显著;涟茶7号、槠叶齐、上梅洲种成熟叶组比嫩叶组果胶含量高,与其果胶氟含量变化相同;黔湄702、湘波绿、黔湄303、湘安17号、寒绿成熟叶组比嫩叶组果胶含量低,与果胶氟含量变化相反。图2C显示,云南乌黑大叶半纤维素氟含量两组差异不显著;其余品种在两组间均差异显著,成熟叶组比嫩叶组半纤维素氟含量高。云南乌黑大叶、槠叶齐9号、福鼎大白茶、上梅洲种半纤维素含量差异不显著;其余品种成熟叶组比嫩叶组半纤维素含量低,与半纤维素氟含量变化相反。图2D显示,云南乌黑大叶、槠叶齐9号、涟茶7号、铁观音两组间纤维素氟含量差异不显著,其余品种在两组间均差异显著,成熟叶组比嫩叶组纤维素氟含量高。碧云纤维素含量差异不显著;其余品种成熟叶组比嫩叶组纤维素含量高,与纤维素氟含量变化相同。
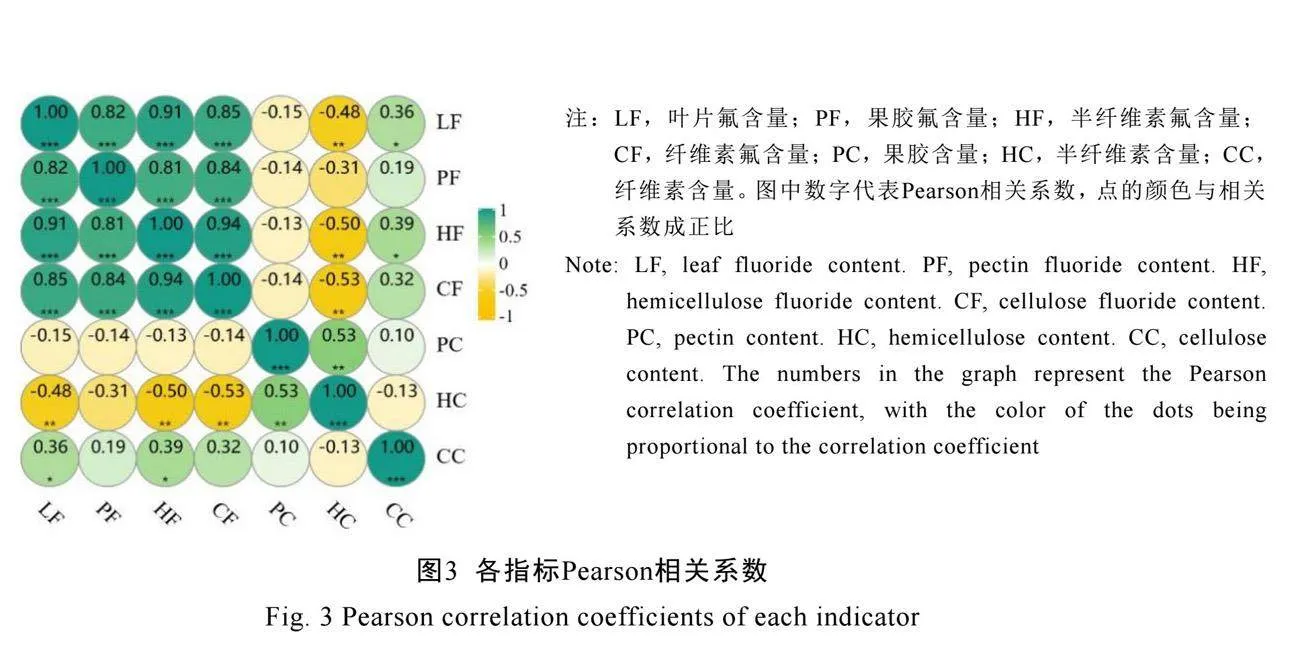
2.2 不同茶树品种叶片细胞壁氟富集相关性及线性回归分析
为探究不同茶树品种叶片是否存在细胞壁氟富集相关规律,本研究对叶片氟含量、果胶氟含量、半纤维素氟含量、纤维素氟含量、果胶含量、半纤维素含量、纤维素含量7个理化指标进行Pearson相关性分析。结果显示(图3),叶片氟含量、果胶氟含量、半纤维素氟含量、纤维素氟含量之间均显著正相关(P<0.001),而细胞壁组分中仅果胶含量与半纤维素含量显著正相关(P<0.01),且半纤维素含量与叶片氟含量、半纤维素氟含量、纤维素氟含量显著负相关(P<0.01)。
基于上述相关性分析,为明确各品种细胞壁组分氟分配情况及细胞壁组分氟含量与叶片氟含量的关系,对嫩叶组及成熟叶组叶片氟含量与果胶氟含量、半纤维素氟含量、纤维素氟含量占比进行线性回归分析。图4A显示,15个品种嫩叶组果胶氟含量在细胞壁组分占比均超过50%,铁观音果胶氟含量为98.93%,占比最高;寒绿果胶氟含量为64.34%,占比最低。值得注意的是,云南乌黑大叶、槠叶齐9号、槠叶齐、铁观音、盘龙山福引、黔湄303、湘波绿并未检测出纤维素氟含量。类似嫩叶组,图4C显示,成熟叶组云南乌黑大叶果胶氟含量为88.61%,占比最高;寒绿果胶氟含量为57.82%,占比最低。对比两组氟分配占比,除涟茶7号、湘波绿2号外,果胶氟含量占比均在成熟叶组中下降,纤维素氟含量占比除涟茶7号外均上升。
回归分析结果显示(图4B、图4D),嫩叶组果胶氟含量占比与叶片氟含量显著负相关(P<0.000 1,R2=0.403 0),半纤维素氟含量占比(P=0.000 4,R2=0.256 7)、纤维素氟含量占比(P<0.000 1,R2=0.588 4)与叶片氟含量显著正相关,果胶氟含量占比、半纤维素氟含量占比、纤维素氟含量占比均差异显著。成熟叶组果胶氟含量占比与叶片氟含量显著负相关(P<0.000 1,R2=0.3844),纤维素氟含量占比与叶片氟含量显著正相关(P<0.000 1,R2=0.614 3),半纤维素氟含量占比与叶片氟含量相关性不显著(P=0.098 4)。这表明在所选品种中无论是嫩叶组还是成熟叶组,叶片氟含量越高的品种,细胞壁中果胶组分氟含量占比反而越低,纤维素组分氟含量占比越高。半纤维素组分则较为复杂,在嫩叶组中叶片氟含量越高的品种其氟含量占比越高,而在成熟叶组中无明显趋势。
2.3 不同品种茶树叶片细胞壁氟富集综合评价及聚类分析
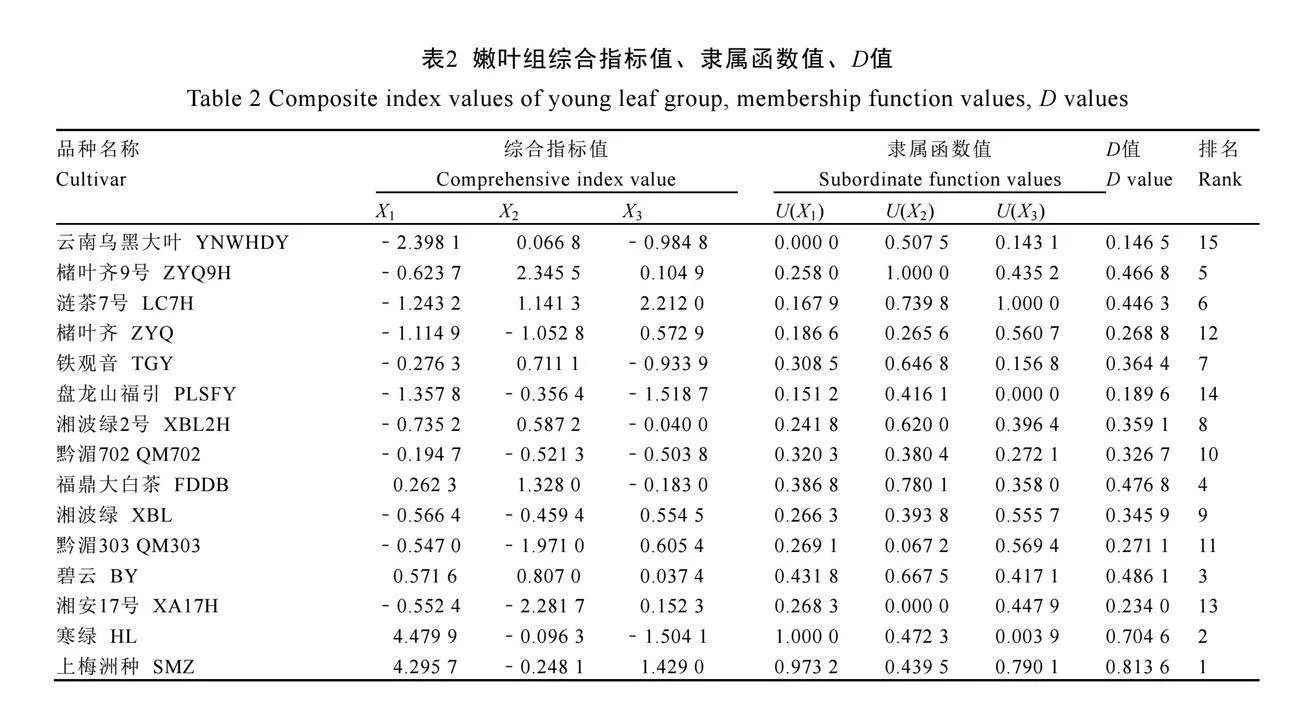
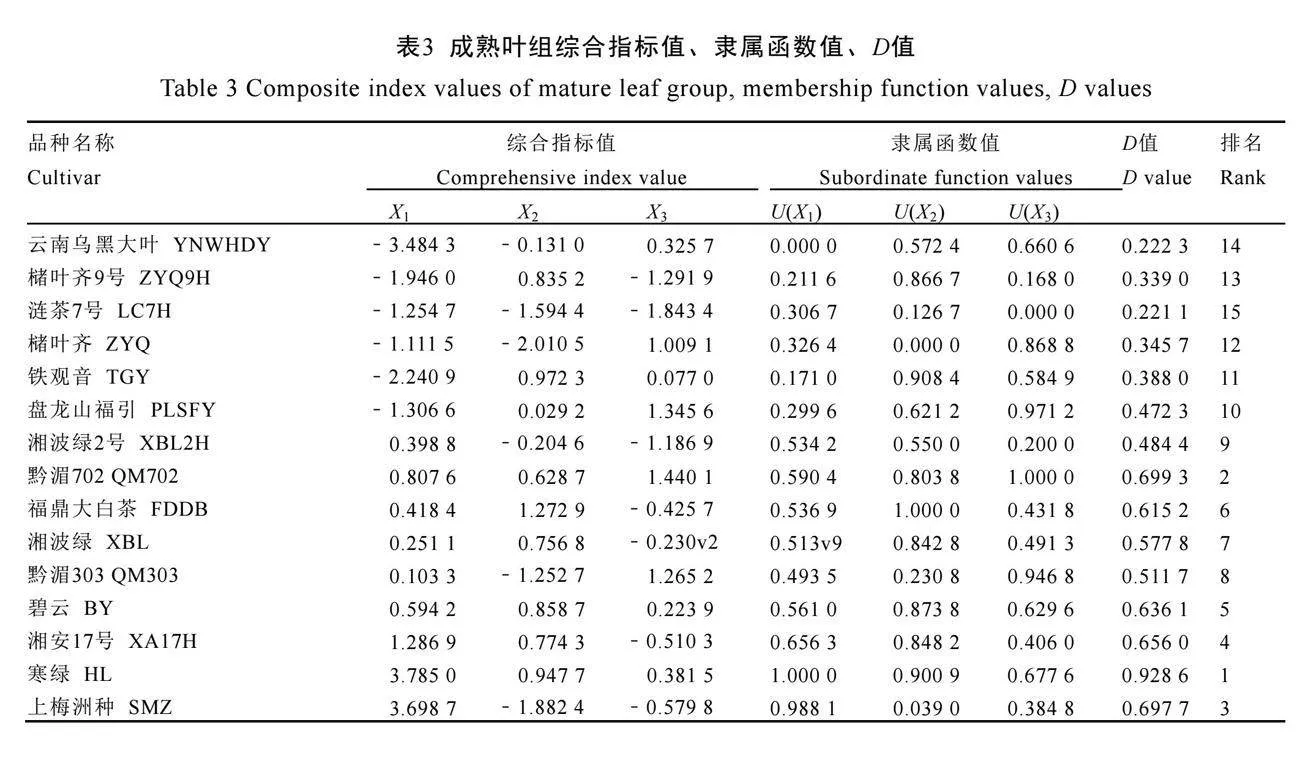
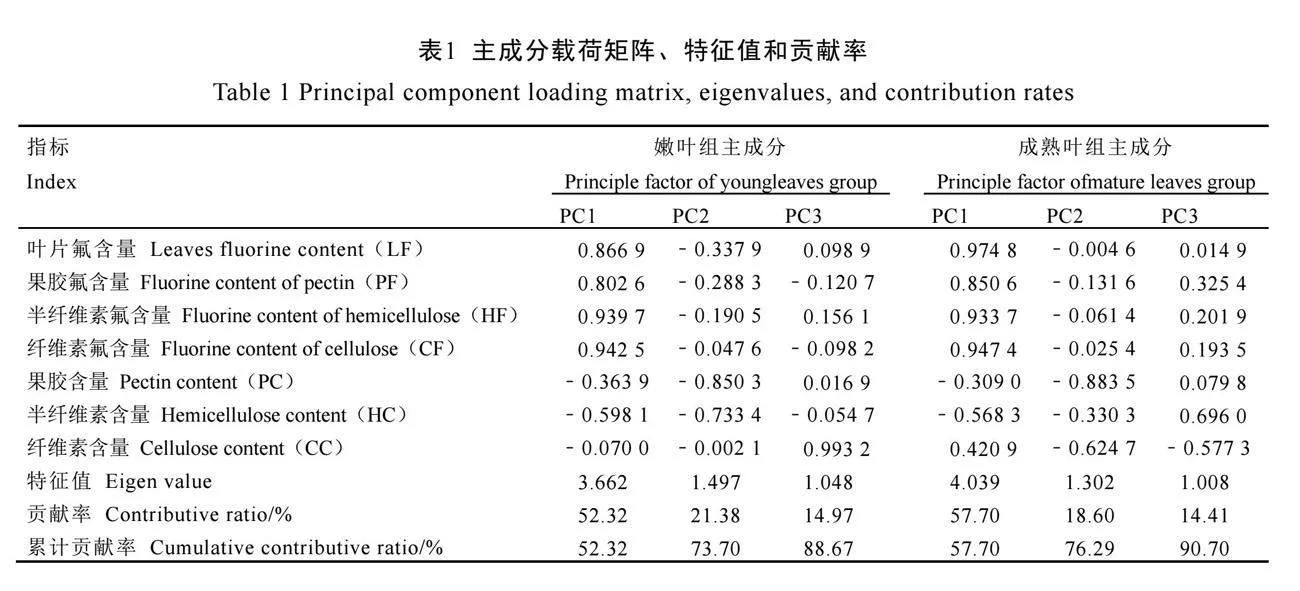
对各品种茶树两组7个指标分别进行主成分分析(表1),均选取3个特征值大于1的主成分。根据表1结果,参考汤云川等[17]公式计算两组3个主成分综合指标值及隶属函数值,根据公式(3)计算综合评价值(D值),见表2、表3。
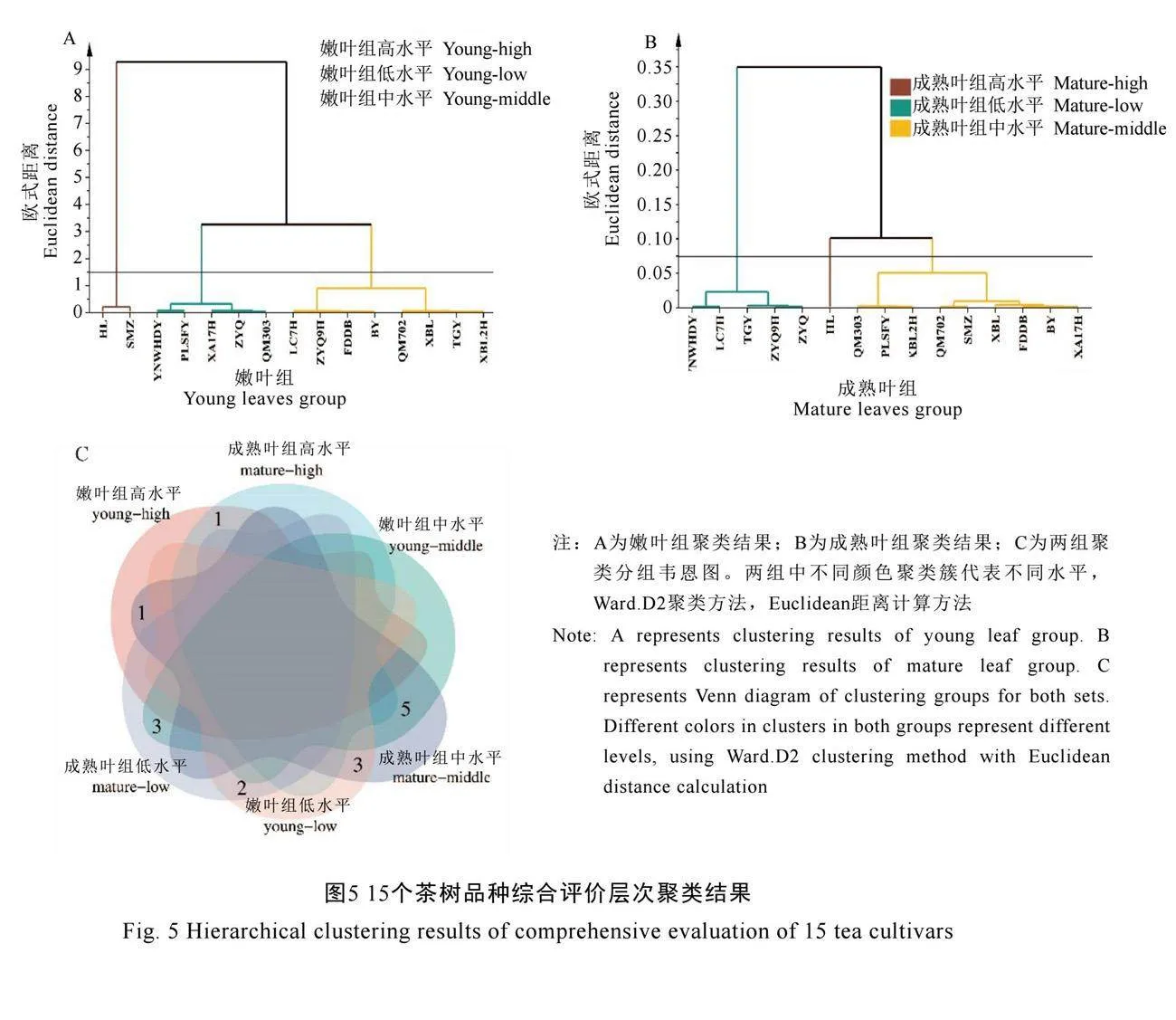
以D值大小为依据进行排名,D值越高表明该品种叶片细胞壁氟富集能力越强。对15个品种嫩叶组及成熟叶组的D值进行层次聚类
分析(图5)。结果显示,嫩叶组平均欧氏距离为1.5,成熟叶组平均欧氏距离为0.075时,15个品种两组中均划分为3类(图5A、图5B)。对聚类结果取交集筛选两组均保持相同细胞壁氟富集水平的品种(图5C),细胞壁氟富集高水平品种为寒绿(D1-4=0.704 6,D5-8=0.928 6);中等水平品种为福鼎大白茶、湘波绿、湘波绿2号、碧云、黔湄702(D1-4=0.326 7~0.486 1,D5-8=0.484 4~0.699 3);低水平品种为云南乌黑大叶、槠叶齐(D1-4=0.146 5~0.268 8,D5-8=0.222 3~
0.345 7)。综合上述分析,对比细胞壁氟富集高水平品种寒绿与低水平品种云南乌黑大叶、槠叶齐。寒绿两组中细胞壁组分含量更低,细胞壁组分氟含量更高;果胶组分氟含量占比更低,纤维素组分氟含量占比更高,倾向细胞壁多个组分富集氟趋势。
3 讨论
3.1 细胞壁组分含量与细胞壁组分氟含量差异关联
高成熟度叶片代表着细胞壁为适应更高
细胞膨压及伸长,作为细胞壁骨架的纤维素合成增加,同时减少“粘合剂”半纤维素使细胞壁松弛[18],本研究结果支持这一结论。成熟叶比嫩叶叶片氟含量更高,半纤维素含量下降而半纤维素氟含量增加且不存在品种差异。罗金蕾等[19]通过梯度氟胁迫也发现类似结果,半纤维素氟含量增加且半纤维素主要成分木葡聚糖含量降低。由此推测,外源氟胁迫和高成熟度叶片氟富集导致的叶片氟含量增加,均可能抑制细胞壁半纤维素合成。
纤维素含量与纤维素氟含量两组差异变化相同,成熟叶组比嫩叶组含量高。在以往研究中,Boex-Fontvieille等[20]通过改变光合作用速率测量纤维素合成速率,结果发现叶片光合作用水平降低但细胞壁仍优先合成纤维素。不同于果胶和半纤维素在胞内高尔基体中组装,纤维素微纤维通过质膜上纤维素合酶合成相对简单[21]。这可能是大部分品种成熟叶纤维素含量增加的原因。所有细胞壁组分中纤维素含量最高而对应氟的占比最低,这与杜亚如[22]研究结果相同,说明纤维素富集氟并不是其主要功能。
果胶是体现品种氟富集差异的重要细胞壁组分,占初生细胞壁组分的30%,而其负电荷官能团却占初生细胞壁的70%[23]。这可以用来解释为什么果胶含量低于半纤维素和纤维素含量,而其氟含量却更高。同时果胶是胞间层主要成分,而纤维素、半纤维素是初生细胞壁主要成分[24]。结合罗金蕾等[19]氟胁迫处理后叶片胞间层较初生细胞壁电子密度更高可知,胞间层可能是茶树叶片细胞壁主要氟积累结构。本研究发现,15个茶树品种中仅涟茶7号、槠叶齐、上梅洲种的成熟叶比嫩叶果胶含量高,且与它们的果胶氟含量变化规律一致。这与氟胁迫下细胞壁组分氟含量的情况相同。Luo等[25]研究发现,外源氟处理提高了果胶代谢合成途径关键酶半乳糖醛酸转移酶(GAUT)、半乳糖醛酸激酶(GALAK)、葡糖醛酸异构酶(GAE)酶活性和基因表达量,表明氟能促进果胶生物合成。湘波绿在氟胁迫下第5叶糖基水解酶、糖基转移酶基因特异性表达,高成熟度叶片光合作用生产的碳水化合物倾向于转化为D-葡萄糖提供更多能量而非合成果胶[26]。这可能是本研究中部分品种成熟叶比嫩叶果胶含量低的原因。铁观音是唯一成熟叶比嫩叶果胶氟含量低的品种,这可能与细胞壁果胶甲酯化修饰相关。高尔基体合成的果胶聚合物高度甲酯化,通过囊泡运输至质膜外,由果胶甲基酯酶(PME)去甲酯化“拆包装”后暴露大量游离羧基以结合离子[27]。Yang等[28]发现,施用钼后冬小麦拔节期叶片虽然果胶含量提高97.03%,但果胶甲基酯酶活性降低,表明钼通过抑制果胶去甲酯化减少可结合钼离子的可溶性果胶转化。这与铁观音的果胶组分差异变化类似,后续将通过检测果胶甲基酯酶活性以及转录组学等方法对品种差异进一步探究。
3.2 茶树细胞壁氟分配及富集规律
刘思怡等[5]研究认为,果胶是细胞壁主要氟富集组分。这与本研究结果一致。本研究显示,成熟叶组相比嫩叶组整体果胶氟含量占比更低,纤维素氟含量占比更高,这说明成熟度更高的叶片倾向细胞壁多组分富集氟。同时,在所选品种中无论嫩叶组还是成熟叶组,高氟品种相比低氟品种,细胞壁主要氟富集的果胶组分氟含量占比更低。这说明高氟品种细胞壁更倾向多组分富集氟。在茶树叶片细胞壁铝分布中也得到类似结果,李春雷等[29]发现高铝品种农抗早细胞壁果胶铝占比(60.1%)低于低铝品种平阳特早茶(78.7%),半纤维素、纤维素铝占比均更高。综上所述,茶树品种叶片氟含量越高,叶片成熟度越高,均表现细胞壁多组分富集氟趋势。
不同品种茶树叶片细胞壁氟富集能力不同。本研究以叶片细胞壁氟富集能力角度筛选出3类保持相同水平的茶树品种。相比细胞壁氟富集低水平的品种云南乌黑大叶和槠叶齐,细胞壁氟富集高水平的品种寒绿各细胞壁组分中氟含量更高,但各细胞壁组分含量更低,这说明寒绿并非通过增加细胞壁组分含量而富集更多氟。Ren等[30]发现,铜胁迫下蓖麻根细胞壁半纤维素铜含量占比最高且对铜胁迫反应更灵敏,而果胶铜结合能力很容易达到饱和。由此推测这可能与寒绿富集氟的机理相似。氟高富集的茶树品种寒绿,可能通过提高各细胞壁组分中氟含量占比从而提高细胞壁富集氟的水平。
本研究从不同茶树品种叶片细胞壁组分与各细胞壁组分中氟含量差异关联、细胞壁氟分配占比、细胞壁氟富集综合评价等方面,开展了茶树细胞壁氟富集规律研究。但相关代谢途径和基因表达尚不明确,未来可从糖代谢能量分配、细胞壁修饰重组等方面对品种细胞壁氟富集差异规律进行深入研究。
参考文献
[1] Shu W S, Zhang Z Q, Lan C Y, et al. Fluoride and aluminium concentrations of tea plants and tea products from Sichuan province, PR China [J]. Chemosphere, 2003, 52(9): 1475-82.
[2] 李春雷. 氟对茶树幼苗生理生化的影响及其作用机制研究[D]. 武汉: 华中农业大学, 2011.
Li C L. Study on the effect and mechanism of fluoride in the physiology and biochemistry of tea seedings [D]. Wuhan: Huazhong Agricultural University, 2011.
[3] Singh B, Avci U, Inwood S E E, et al. A specialized outer layer of the primary cell wall joins elongating cotton fibers into tissue-like bundles [J]. Plant Physiology, 2009, 150(2): 684-699.
[4] 高慧敏, 朱晓静, 李攀攀, 等. 茶多糖对氟离子的吸附特性研究[J]. 茶叶科学, 2016, 36(4): 396-404.
Gao H M, Zhu X J, Li P P, et al. Research on adsorption characteristics of tea polysaccharides to fluorine [J]. Journal of Tea Science, 2016, 36(4): 396-404.
[5] 刘思怡, 朱晓静, 房峰祥, 等. 茶树叶片氟亚细胞分布及其与细胞壁结合特性的研究[J]. 茶叶科学, 2018, 38(3): 305-312.
Liu S Y, Zhu X J, Fang F X, et al. Fluorine subcellular distribution and its combining characteristics with cell wall in tea leaves (Camellia sinensis) [J]. Journal of Tea Science, 2018, 38(3): 305-312.
[6] 朱晓静. 茶多糖氟测定方法及多糖与氟结合方式的初步研究[D]. 武汉: 华中农业大学, 2017.
Zhu X J. Study on the determination method of fluorine in tea polysaccharide and way of combination with the tea polysaccharide [D]. Wuhan: Huazhong Agricultural University, 2017.
[7] Yang X, Yu Z, Zhang B, et al. Effect of fluoride on the biosynthesis of catechins in tea [Camellia sinensis (L.) O. Kuntze] leaves [J]. Scientia Horticulturae, 2015, 184: 78-84.
[8] 陈瑞鸿, 梁月荣, 陆建良, 等. 茶树对氟富集作用的研究[J]. 茶叶, 2002(4): 187-190.
Chen R H, Liang Y R, Lu J L, et al. Studies on fluorine enrichment in tea plant (Camellia sinensis) [J]. Journal of Tea, 2002(4): 187-190.
[9] 蔡荟梅, 彭传燚, 李成林, 等. 三个品种茶树氟富集特性及其在亚细胞中的分布[J]. 中国农业科学, 2013, 46(8): 1668-1675.
Cai H M, Peng C Y, Li C L, et al. Fluoride accumulation and its subcellular distribution in three tea plants [J]. Scientia Agricultura Sinica, 2013, 46(8): 1668-1675.
[10] Renard C. Variability in cell wall preparations: quantification and comparison of common methods [J]. Carbohydrate Polymers, 2005, 60(4): 515-522.
[11] Figueroa C R, Opazo M C, Vera P, et al. Effect of postharvest treatment of calcium and auxin on cell wall composition and expression of cell wall-modifying genes in the Chilean strawberry (Fragaria chiloensis) fruit [J]. Food Chemistry, 2012, 132(4): 2014-2022.
[12] 朱晓静, 房峰祥, 张月华, 等. 茶叶及茶多糖中氟测定前处理方法的比较研究[J]. 茶叶科学, 2015, 35(2): 145-150.
Zhu X J, Fang F X, Zhang Y H, et al. Comparison on the pretreatment methods of tea and tea polysaccharides for determination of fluorine content [J]. Journal of Tea Science, 2015, 35(2): 145-150.
[13] 刘生, 郑添妍, 高晗, 等. 不同提取方法对黄秋葵果胶含量和结构特性的影响[J]. 现代食品科技, 2019, 35(3): 161-168.
Liu S, Zheng T Y, Gao H, et al. Effects of different extraction methods on the content and structural properties of pectin from okra [J]. Modern Food Science and Technology, 2019, 35(3): 161-168.
[14] 王政, 黄广民, 赵斌, 等. 新鲜椰衣纤维表面半纤维素含量的测定及去除[J]. 热带作物学报, 2018, 39(1): 156-161.
Wang Z, Huang G M, Zhao B, et al. Determination and removal of hemicellulose content on the surface of coir fiber [J]. Chinese Journal of Tropical Crops, 2018, 39(1): 156-161.
[15] 安玉民, 王菊葵, 黄烨, 等. 马铃薯秸秆中纤维素与半纤维素含量的测定[J]. 现代农业科技, 2016(17): 159-160.
An Y M, Wang J K, Huang Y, et al. Determination of cellulose and hemicellulose content in potato stalk [J]. Modern Agricultural Science and Technology, 2016(17): 159-160.
[16] 中华人民共和国卫生部. 砖茶含氟量: GB 19965—2005 [S]. 北京: 中国标准出版社, 2005.
Ministry of Health of the People's Republic of China. Fluoride content in brick tea: GB 19965—2005 [S]. Beijing: Standards Press of China, 2005.
[17] 汤云川, 张庆沛, 冯焱, 等. 大凉山地区不同马铃薯品种产量和营养品质的综合评价[J]. 中国蔬菜, 2024(6): 89-100.
Tang Y C, Zhang Q P, Feng Y, et al. Comprehensive evaluation of yield and nutritional quality of different potato varieties in daliangshan zone [J]. China Vegetables, 2024(6): 89-100.
[18] Joo J Y, Kim M S, Sung J. Transcriptional changes of cell wall organization genes and soluble carbohydrate alteration during leaf blade development of rice seedlings [J]. Plants, 2021, 10(5): 823. doi: 10.3390/plants10050823.
[19] 罗金蕾. 茶树叶细胞壁富集氟机制研究[D]. 武汉: 华中农业大学, 2020.
Luo J L. Study on mechanism of fluorine accumulation in cell wall of tea plant leaf [D]. Wuhan: Huazhong Agricultural University, 2020.
[20] Boex-Fontvieille E, Davanture M, Jossier M, et al. Photosynthetic activity influences cellulose biosynthesis and phosphorylation of proteins involved therein in Arabidopsis leaves [J]. Journal of Experimental Botany, 2014, 65(17): 4997-5010.
[21] Pedersen G B, Blaschek L, Frandsen K E H, et al. Cellulose synthesis in land plants [J]. Molecular Plant, 2023, 16(1): 206-231.
[22] 杜亚如. 茶树叶片细胞壁果胶和半纤维素参与氟累积的机制研究[D]. 武汉: 华中农业大学, 2019.
Du Y R. Studies on mechanism of cell wall pectin and hemicellulose involved in the fluorine accumulation in tea leaves [D]. Wuhan: Huazhong Agricultural University, 2019.
[23] Jarvis M C. Structure and properties of pectin gels in plant cell walls [J]. Plant, Cell & Environment, 2006, 7(3): 153-164.
[24] Cosgrove D J. Structure and growth of plant cell walls [J] . Nature Reviews Molecular Cell Biology, 2024, 65(17): 340-358.
[25] Luo J, Hu K, Qu F, et al. Metabolomics analysis reveals major differential metabolites and metabolic alterations in tea plant leaves (Camellia sinensis L.) under different fluorine conditions [J]. Journal of Plant Growth Regulation, 2021, 40(2): 798-810.
[26] Yang P, Liu Z, Zhao Y, et al. Comparative study of vegetative and reproductive growth of different tea varieties response to different fluoride concentrations stress [J]. Plant Physiology and Biochemistry, 2020, 154: 419-428.
[27] Sénéchal F, Wattier C, Rustérucci C, et al. Homogalacturonan-modifying enzymes: structure, expression, and roles in plants [J]. Journal of Experimental Botany, 2014, 65(18): 5125-60.
[28] Yang X, Gan Q, Sun X, et al. Effects of molybdenum on cell wall component of wheat leaf under different growth stages [J]. Journal of Soil Science and Plant Nutrition, 2021, 21(1): 587-595.
[29] 李春雷, 徐红梅, 刘杰, 等. 铝在茶树叶片亚细胞中的分布及其与细胞壁的结合研究[J]. 浙江农业学报, 2023, 35(3): 509-514.
Li C L, Xu H M, Liu J, et al. Aluminum subcellular distribution and its combining characreristics with cell wall in tea leaves [J]. Acta Agriculturae Zhejiangensis, 2023, 35(3): 509-514.
[30] Ren C, Qi Y, Huang G, et al. Contributions of root cell wall polysaccharides to Cu sequestration in castor (Ricinus communis L.) exposed to different Cu stresses [J]. Journal of Environmental Sciences, 2020, 88: 209-216.
