连作年限对喀斯特山地设施辣椒根际土壤养分、酶活性及真菌群落结构的影响
2024-11-07余高赵仕龙冯顺梅陈芬


摘要:辣椒连作所引发的植株发育不良、产量下降、根腐病频发等问题,严重影响辣椒产业可持续发展,造成重大的经济损失,但其具体影响机制尚不清楚。采用野外田间调查、室内化验分析相结合的方法,对贵州铜仁设施大棚辣椒连作1、3、5年的土壤pH值、有机质含量、养分含量、生态化学计量特征、酶活性、真菌群落结构及其相关性进行研究。结果表明,随着设施辣椒连作年限的延长,土壤pH值和有机质含量降低,土壤全磷、有效磷含量增加,碱解氮、速效钾含量则呈现先下降后上升的趋势,而土壤全氮、全钾含量变化则不明显。长期连作会影响土壤养分间的平衡供应,连作5年时土壤C/N、C/P、N/P较未定植过辣椒的土样(CK)分别降低11.35%、26.69%、17.44%。除连作1、3年的土壤脲酶活性与CK相比略有升高外,过氧化氢酶、蔗糖酶、酸性磷酸酶活性均随着设施辣椒连作年限的增加而不断降低。长期连作也会影响土壤真菌群落结构,连作3年土壤子囊菌门的相对丰度分别较连作1、5年高26.04%、8.31%;连作1年土壤被孢霉门的相对丰度分别较连作3、5年高12.98%、14.93%;连作1年土壤担子菌门的相对丰度较连作3年高2.11%。冗余分析结果表明,土壤有机质含量和蔗糖酶活性是影响设施辣椒连作土壤真菌群落结构的主要环境因子。相关性网络分析表明,土壤pH值、有机质含量、全磷含量、N/P、C/N、C/P、碱解氮含量、过氧化氢酶活性、酸性磷酸酶活性和蔗糖酶活性是影响连作土壤子囊菌门、担子菌门、捕虫霉亚门、被孢霉门、球囊菌门、壶菌门、毛霉门和油壶菌门等真菌群落结构的主要环境因子。综上所述,设辣椒连作障碍产生的原因可能是连作改变了土壤的理化性质,降低了土壤酶活性,影响了土壤真菌群落结构,进而破坏了土壤微生态平衡。
关键词:辣椒;连作;土壤养分;土壤酶活性;化学计量特征;真菌群落结构;喀斯特山地
中图分类号:S641.304 文献标志码:A
文章编号:1002-1302(2024)18-0270-07
收稿日期:2023-09-26
基金项目:贵州省梵净山地区生物多样性保护与利用重点实验室项目(编号:黔科合平台人才[2020]2003);铜仁市科学技术局计划(编号:铜市科研[2021]72号);铜仁市科技产学研合作平台项目(编号:铜市科研[2021]33号);铜仁学院研究生教育创新计划(编号:2023yjscxjh011)。
作者简介:余 高(1988—),男,湖南益阳人,硕士,副教授,主要从事土壤肥力与生态修复研究。E-mail:httywwwyu1014@sina.com。
通信作者:陈 芬,博士,教授,主要从事土壤肥力与改良研究。E-mail:chenfen2018@126.com。
辣椒是贵州省主要经济作物之一。据统计,2022年贵州省辣椒种植面积和产量分别达到36.2万hm2和751万t,均位居全国第一。贵州省是典型的喀斯特地形地貌,山地多、平坝少,辣椒种植基地较为集中,加上贵州省的黄壤土存在“黏、酸、瘦、瘠”等特点,近年来,随着贵州省设施辣椒种植面积的持续增长,复种指数以及农药、化肥用量的不断增加,而有机肥供应不足,造成土壤肥力不断下降,辣椒连作障碍日趋严重,由此导致土壤养分失调、病虫害问题加剧、产质量降低等一系列制约贵州省辣椒产业可持续发展的严重问题。
大量研究发现,长期连作会导致土壤养分失调、酶活性降低、理化性质恶化、自毒物质累积和土壤微生物菌群失调等,同时这些因素的综合作用会导致土壤质量下降、病虫害加剧、作物生长受阻[1-6]。设施栽培下由于其温室小气候以及化肥、农药的过量施用导致作物连作问题更为严重[5]。如岳思君等研究发现,硒砂瓜长期连作会降低土壤pH值以及有机质、速效钾含量,抑制土壤有机碳的矿化,阻碍土壤养分循环[7]。郑立伟等的研究表明,设施甜瓜连作会通过降低土壤酶活性而阻碍土壤养分的循环转化和供应[8]。Liu等的研究表明,大豆连作增加了链格孢属、Boeremia、Lectera longa和尖镰孢菌的相对丰度[9]。张敏等的研究表明,光果甘草连作能显著降低有益菌节杆菌属、假单胞菌属和Naganishia的相对丰度,而增加病原真菌镰刀菌属和亡革菌属的相对丰度[10]。目前,有部分研究证实,辣椒连作障碍主要是由辣椒连作导致的土壤养分失衡以及微生物群落发生改变等共同引起的[11-12],但二者之间的内在机制和关键影响因素有待进一步研究。针对具有典型喀斯特地形地貌的贵州黄壤区,关于设施辣椒连作后根际土壤环境因子(养分特性和酶活性)与微生物群落之间内在关系的研究鲜有报道。因此,本研究比较分析不同连作年限下设施辣椒根际土壤真菌群落结构、土壤pH值、有机质含量、养分含量及其生态化学计量特征、酶活性及内在关联,以期为贵州黄壤区设施辣椒连作障碍研究和土壤环境的生态修复提供理论参考和实践依据。
1 材料与方法
1.1 试验区概况
试验区位于贵州省江口县坝盘镇挂扣村坝区大棚蔬菜种植基地(108°53′47″E,27°43′27″N),该地区处于云贵高原向黔东、湘西低丘陵过渡地带,受梵净山的影响,小气候特点突出,年均气温16.2 ℃,年平均日照时间1 257.3 h,无霜期288 d,年均降水量1 369.6 mm,年均相对湿度81%,海拔324 m,属于亚热带季风湿润气候区。辣椒种植为1年2茬,每茬基肥施用量为复合肥(N、P2O5、K2O含量均为15%)900 kg/hm2、商品有机肥1 200 kg/hm2,辣椒生育期肥水管理保持一致。
1.2 样品采集
于2022年8月收获期采集研究区连作1、3、5年的辣椒根际土壤样品,以种植区未定植过辣椒的土样为对照(CK)。采用五点取样法采集辣椒根际0~20 cm表层混合土壤1 kg,重复3次,将土样充分混合均匀并按四分法去除多余土壤后,保留0.5 kg土样,低温保存并迅速带回实验室。一部分土样保存于-80 ℃冰箱中,用于土壤真菌群落结构测定[1];一部分土样室内自然风干、去除杂质、粉碎并过筛后密封保存,用于检测土壤理化指标。
1.3 测定指标及方法
根际土壤pH值以及有机质、养分含量均参照鲍士旦的方法[13]进行测定;土壤酶活性参考关松荫的方法[14]进行测定,其中,过氧化氢酶活性采用高猛酸钾滴定法,以20 min后1 g土壤消耗0.1 mol/L KMnO4溶液的微升数表示;脲酶活性测定采用靛酚比色法,以1 g土壤在37 ℃下培养24 h后酶解尿素释放NH-3-N的毫克数来表示;蔗糖酶活性测定采用3,5-二硝基水杨酸比色法,以1 g土壤在37 ℃下培养24 h后产生葡萄糖的毫克数来表示;酸性磷酸酶活性采用磷酸苯二钠比色法,以1 g土壤在37 ℃下24 h后释放酚的毫克数来表示。土样的DNA提取和真菌的高通量测序委托第三方检测公司上海派森诺生物科技股份有限公司完成。
1.4 数据分析
采用Microsoft Excel 2010、Origin和SPSS 24.0 软件对数据进行处理、作图和分析,采用R语言进行冗余分析和关联网络分析。
2 结果与分析
2.1 不同连作年限对设施辣椒土壤pH值和有机质含量的影响
由图1可以看出,设施辣椒连作会降低土壤pH值,且随着连作年限的延长,土壤pH值呈现逐渐降低趋势。其中,连作1、3、5年土壤pH值较CK分别降低3.21%、4.28%、8.56%,且差异均达到显著或极显著水平。连作1年土壤有机质含量最高,达到21.87 g/kg,较CK极显著提高11.20%,较连作3、5年土壤高6.58%、7.86%,但与2个连作年限差异不显著。
2.2 不同连作年限对设施辣椒土壤全量养分含量的影响
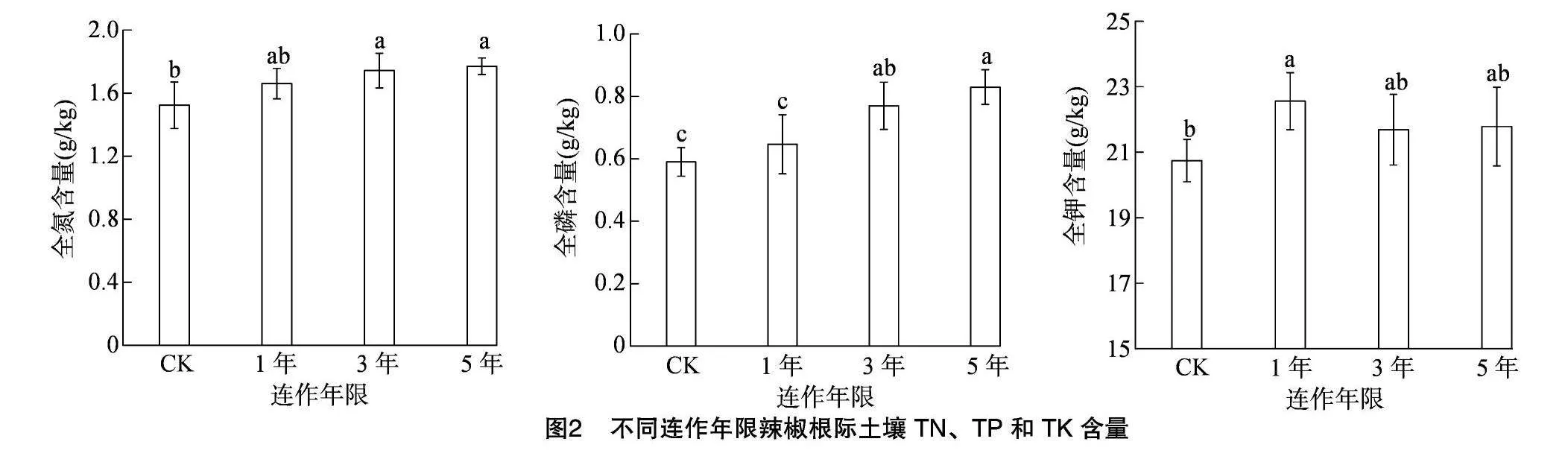
由图2可以看出,与CK相比,连作可在一定程度上提高设施辣椒土壤全氮、全磷、全钾含量。随着连作年限的延长,设施辣椒土壤全氮、全磷含量呈现逐渐增加的趋势,全钾含量呈先降低后升高的趋势。其中,连作3、5年土壤全氮和全磷含量较CK分别显著提高14.47%、16.45%和30.51%、40.68%,连作1年土壤全氮、全磷含量与CK间差异不显著;连作1年土壤全钾含量较CK显著提高8.74%,但与连作3、5年土壤差异不显著,且连作3、5年土壤全钾含量与CK间差异不显著。
2.3 不同连作年限设施辣椒土壤全量养分生态化学计量特征
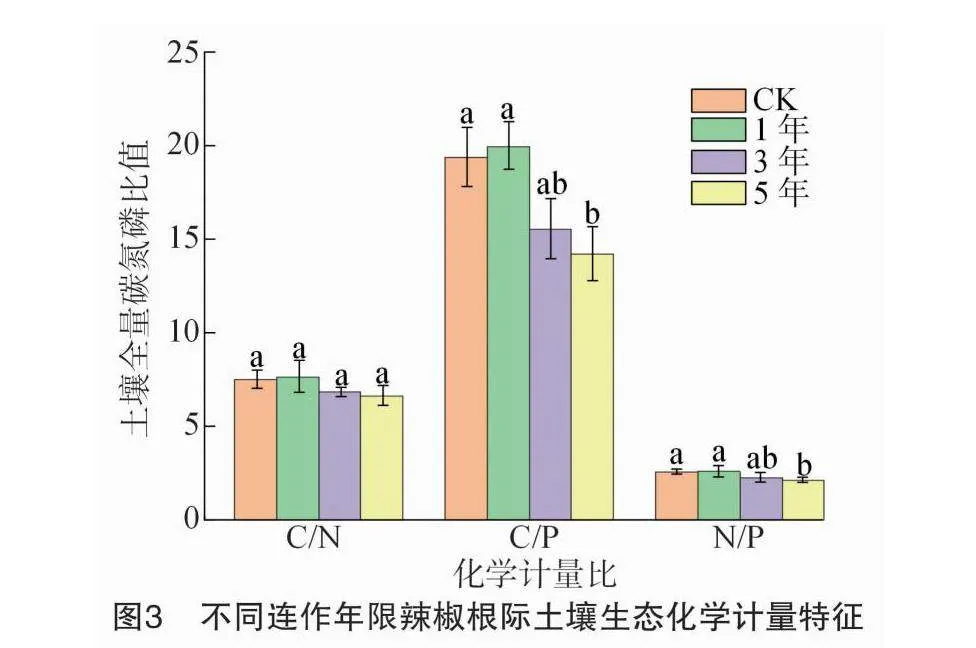
由图3可知,不同连作年限设施辣椒土壤C/N、C/P和N/P变化范围分别为6.64~7.64、14.17~29.62和2.13~2.58。随着连作年限的增加,设施辣椒土壤C/N、C/P和N/P总体呈逐渐下降的趋势。与CK相比,连作1、3年土壤C/N、C/P和N/P均无显著变化,连作至5年时土壤C/N降低11.35%,但差异不显著,而C/P、N/P分别降低26.69%、17.44%,且差异均达显著水平。随着设施辣椒连作年限的延长,土壤全量养分生态化学计量比降幅表现为C/P>C/N>N/P。
2.4 不同连作年限对设施辣椒土壤速效养分含量的影响
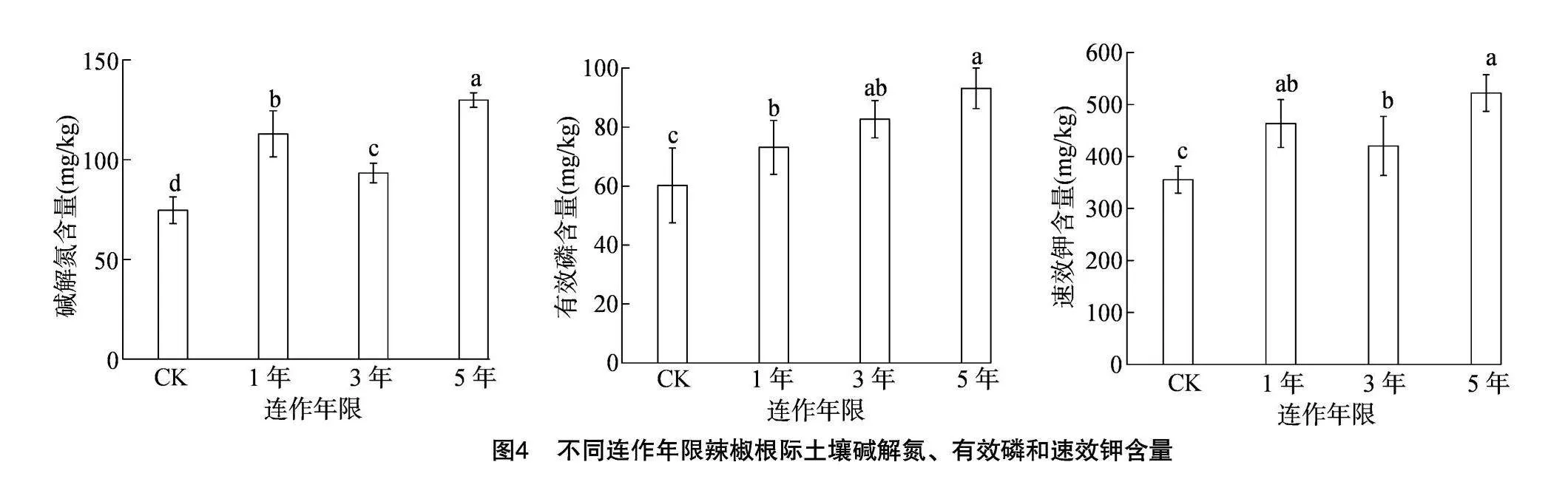
由图4可知,与CK相比,设施辣椒连作均能明显提高土壤碱解氮、有效磷、速效钾含量。随着连作年限的延长,设施辣椒根际土壤碱解氮、速效钾含量呈先降低后升高的变化趋势,而有效磷含量呈逐渐增加的趋势。其中,连作5年土壤碱解氮含量较CK、连作1年、连作3年分别极显著或显著提高74.03%、14.98%、39.16%;连作5年土壤有效磷含量最高,达到93.16 mg/kg,较CK、连作1年土壤分别极显著或显著提高54.77%、27.42%,与连作3年土壤差异不显著,但仍提高12.68%;连作5年土壤速效钾含量达到521.97 mg/kg,较CK、连作3年土壤分别极显著或显著提高46.92%、24.23%,与连作1年土壤差异不显著,但仍提高12.66%。
2.5 不同连作年限对设施辣椒土壤酶活性的影响
从图5可以看出,与CK相比,设施辣椒连作降低了根际土壤过氧化氢酶、蔗糖酶、酸性磷酸酶活性,提高了连作1、3年的土壤脲酶活性。随着连作年限的延长,根际土壤过氧化氢酶、脲酶、蔗糖酶、酸性磷酸酶活性均呈不断降低趋势。其中,连作5年土壤过氧化氢酶、脲酶、蔗糖酶、酸性磷酸酶活性分别较CK、连作1年、连作3年显著降低12.11%~17.33%、28.81%~36.54%、38.71%~68.07%、61.25%~80.91%。不同连作年限的土壤蔗糖酶、酸性磷酸酶活性均与CK之间的差异达到极显著水平,连作1年的过氧化氢酶活性和连作3年的脲酶活性均与CK之间差异均不显著。
2.6 土壤真菌群落结构
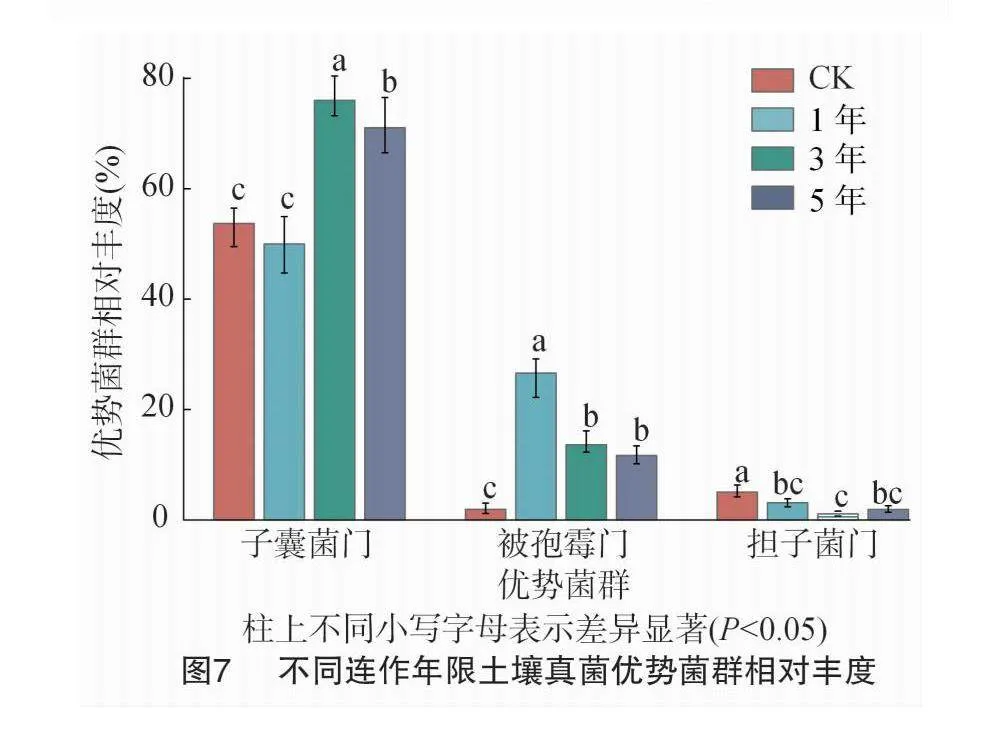
所有采集样本的土壤真菌主要来自子囊菌门、被孢霉门、担子菌门、毛霉门和球囊菌门等(图6)。其中,子囊菌门、被孢霉门和担子菌门占所有土壤微生物总数的78.15%,是所测土壤的优势菌群。不同连作年限下,设施辣椒连作3年土壤子囊菌门的相对丰度分别较连作1、5年显著高26.04%、8.31%(图7)。连作1年土壤被孢霉门的相对丰度分别较连作3、5年显著高12.98%、14.93%。连作1年土壤担子菌门的相对丰度分别较连作3、5年提高2.11%、1.23%,但差异不显著。与CK相比,除连作1年土壤子囊菌门的相对丰度变化不明显外,设施辣椒连作均会显著提高土壤子囊菌门和被孢霉门的相对丰度,降低担子菌门的相对丰度。
2.7 设施辣椒土壤真菌群落与环境因子的关系
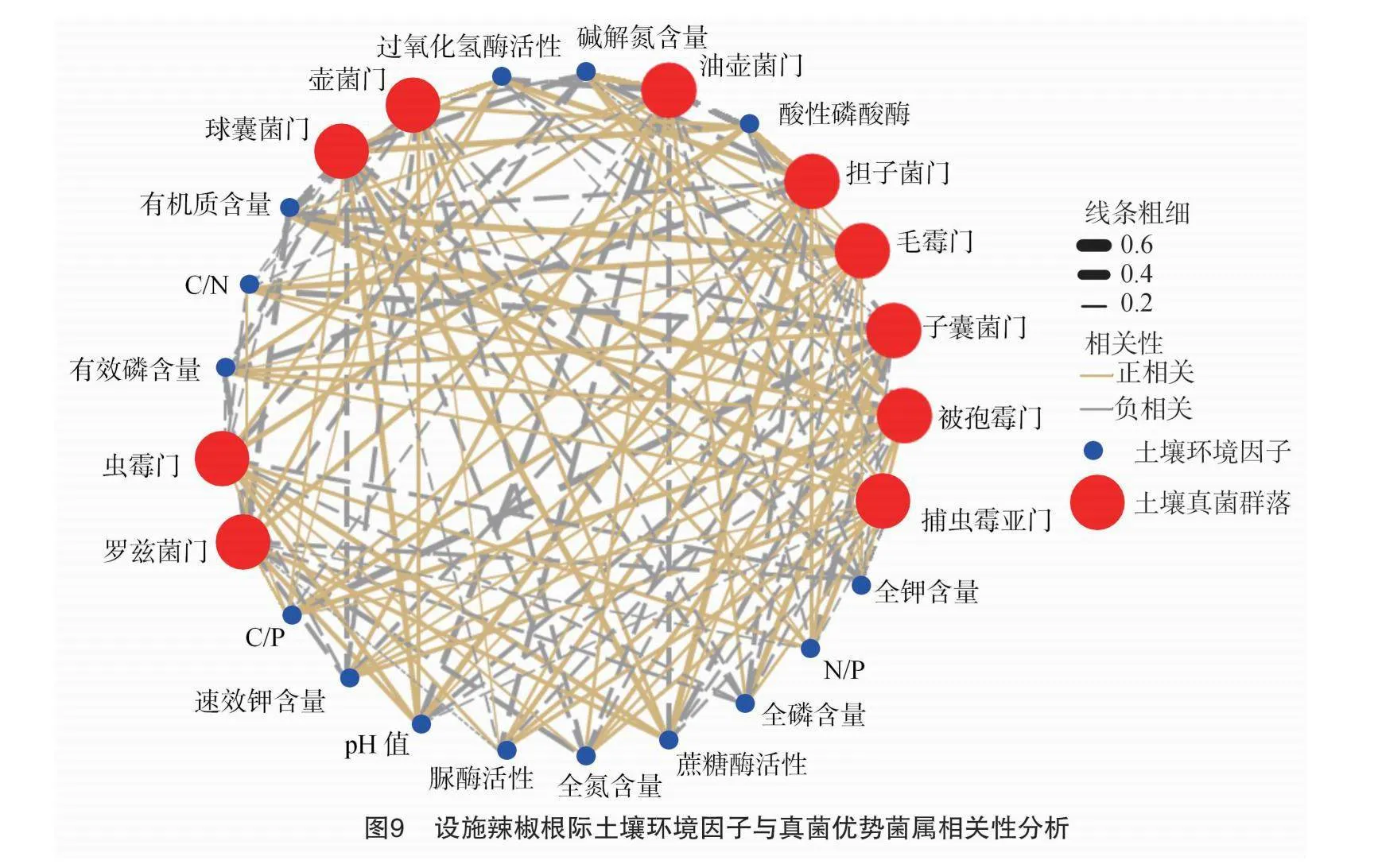
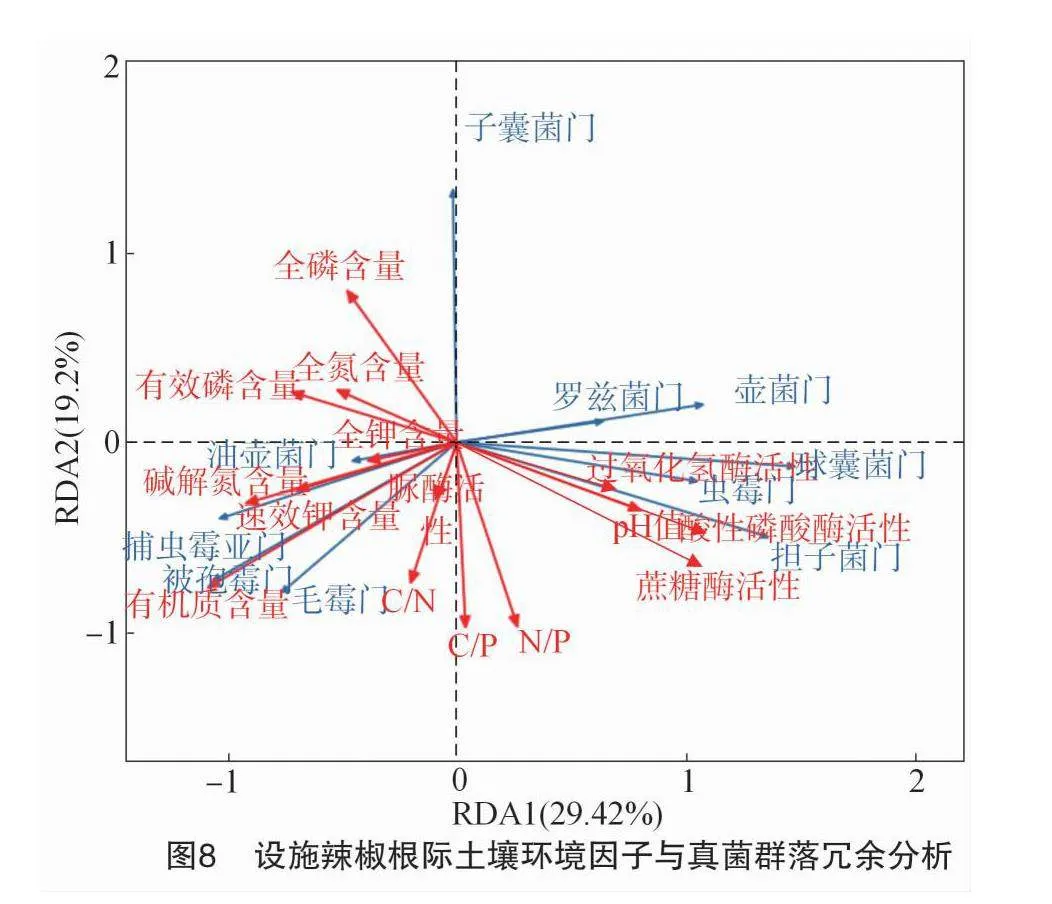
由图8可知,第1排序轴和第2排序轴分别可解释土壤真菌群落变化的29.42%和19.2%,其中,第1排序轴主要与土壤pH值、有机质含量、碱解氮含量、蔗糖酶活性及酸性磷酸酶活性相关性较大;第2排序轴主要与全磷含量、C/P和N/P的相关性较大。采用envfit函数对设施辣椒根际土壤pH值、有机质含量、养分含量、生态化学计量特征、酶活性等环境因子进行显著性检验,结果显示,有机质含量(r2=0.753,P=0.002)、蔗糖酶活性(r2=0.483,P=0.041)对设施辣椒土壤真菌群落结构的影响分别达极显著(P<0.01)和显著水平(P<0.05)。表明影响连作条件下设施辣椒根际土壤真菌群落结构的主要因子为土壤有机质含量和蔗糖酶活性。
采用R语言将相对丰度位列前十的土壤真菌群落与设施辣椒根际土壤pH值、有机质含量、养分含量、生态化学计量特征、酶活性等环境因子进行相关性网络分析,结果如图9所示,可以看出,毛霉门和捕虫霉亚门的相对丰度与有机质含量,担子菌门的相对丰度与土壤pH值、蔗糖酶活性、酸性磷酸酶活性,油壶菌门的相对丰度与碱解氮含量,子囊菌门的相对丰度与全磷含量,球囊菌门的相对丰度与酸性磷酸酶活性、蔗糖酶活性呈显著正相关(P<0.05);子囊菌门的相对丰度与N/P、C/N,球囊菌门的相对丰度与有机质含量,油壶菌门的相对丰度与土壤pH值、过氧化氢酶活性、酸性磷酸酶活性,壶菌门的相对丰度与有机质含量呈显著负相关(P<0.05);被孢霉门的相对丰度与有机质含量呈极显著正相关(P<0.01);子囊菌门与C/P呈极显著负相关(P<0.01)。进一步对相关性网络分析数据进行过滤和去重后发现,土壤pH值是影响担子菌门和油壶菌门群落结构的主要环境因子;有机质含量是影响毛霉门、捕虫霉亚门、球囊菌门、被孢霉门和壶菌门群落结构的主要因子;全磷含量、过氧化氢酶活性、N/P、C/N和C/P是影响子囊菌门群落结构的主要因子;碱解氮含量是影响油壶菌门群落结构的主要因子;酸性磷酸酶活性和蔗糖酶活性是影响球囊菌门和担子菌门群落结构的主要因子。
3 讨论与结论
本研究发现,设施辣椒连作会降低土壤pH值而引起土壤酸化,这与韩海蓉等的uFJA3qMpzd0851N0rAz34LMUzyd11COyDlnAHxdf2z4=研究结果[15-16]一致。究其原因,一方面是本研究中的辣椒种植为1年2季,种植户为追求高产而盲目施用大量化肥,但有机肥施用不足,加上大棚温度高,土壤水分蒸发量大,导致土壤中的盐分不断在地表累积而造成土壤酸化,pH值降低[1-2];另一方面可能是设施辣椒每年投入的肥料种类及用量基本一致,单一的辣椒品种对土壤养分的选择性吸收相对固定,使得土壤胶体上吸附的H+和Al3+在土壤中常年积累,浓度不断升高[17]。作为植物养分吸收和储存的载体,土壤养分变化对植物的生理生化、生长及产量和品质都有一定的影响[18]。大量研究表明,连作过程中,作物在生育期间对养分的选择性吸收会造成土壤中某些养分亏缺,而其他养分则大量累积,导致土壤养分失衡,进而影响作物生长[19-21]。杨阳等研究了连作对党参根际土壤理化性质的影响,结果发现,党参连作会增加土壤全氮、全磷、速效钾含量,而速效磷含量变化则无明显规律[22]。Li等研究了不同连作年限下温室黄瓜土壤的理化指标差异,结果发现,连作年限延长会导致土壤碱解氮含量呈现先升后降的变化趋势,而土壤有机质、有效磷、速效钾含量则呈现先升后降再升的变化趋势[23]。本研究结果发现,连作条件下,设施辣椒土壤有机质含量降低,全磷、有效磷含量增加,速效钾含量呈先降后升趋势,而土壤全氮、全钾含量变化趋势不明显。这与前人的研究结果[22-23]存在一定差异,这可能与土壤类型、研究区作物种类、肥料施用种类和施用量存在差异等因素有关。土壤养分化学计量特征是判断土壤有机构成和土壤质量高低的重要指标之一,耕地土壤中的C、N、P除受到气候及植kIGNKQqFLwN+qx+qlGO/O9P3z/ORKgZtHSO8l0ao0OA=被等自然因素影响外,受土地利用方式、耕作及施肥等人为活动的影响更为强烈[24]。本研究中,随着设施辣椒土壤连作年限的延长,土壤C/N、C/P和N/P总体呈逐渐下降趋势,连作至5年时土壤C/N、C/P、N/P较CK分别降低11.35%、26.69%、17.44%,表明设施辣椒连作加速了土壤微生物对土壤中有机质的分解,同时外源氮、磷、钾供应量的增加,更不利于土壤有机质的积累,从而使土壤养分平衡被破坏,导致土壤中氮、磷、钾等养分对土壤肥力的限制性越来越小。这与王琪等在烤烟连作中的研究结果[25]基本一致。
土壤酶是一种生物催化剂,可促进土壤中有机物分解矿化,直接或间接参与土壤养分循环过程[26],能有效反映土壤微生物代谢水平,其活性可以作为衡量土壤环境质量水平及肥力的重要生物指标[27]。洪彪等通过研究连作对广藿香扦插苗土壤酶活性的影响,发现广藿香连作会引起土壤微生态系统失衡,并降低土壤过氧化氢酶、脲酶、蔗糖酶、磷酸酶活性[28]。侯乾等的研究也表明,随着马铃薯连作年限的延长,土壤过氧化氢酶、脲酶、蔗糖酶、酸性磷酸酶活性均降低[29]。刘姣姣在研究花椒连作障碍的发生机制中发现,土壤磷酸酶、过氧化氢酶、蔗糖酶活性随着花椒连作年限的增加均呈先降后升的趋势,而土壤脲酶活性则无明显变化规律[30]。本研究发现,除连作1、3年的土壤脲酶活性与CK相比略有升高外,过氧化氢酶、蔗糖酶、脲酶、酸性磷酸酶活性均会随着设施辣椒连作年限的增加而不断降低,这与以上研究结果存在一定的差异,可能与研究区域的土壤类型、施肥种类以及气候差异等因素有关[31]。
微生物群落是维持土壤生态系统平衡的重要组成部分[32]。本研究表明,辣椒连作均会增加土壤被孢霉门的相对丰度,降低担子菌门的相对丰度,这主要与土壤环境因子的变化有关。冗余分析结果表明,土壤有机质含量和蔗糖酶活性是影响设施辣椒连作根际土壤真菌群落结构的主控因子,这与Pang等的研究结果[33]相似。连作可导致土壤有机质含量下降,而土壤有机质作为农田肥力高低的核心指标之一[34],其下降会导致土壤变得黏稠,透气性被严重破坏,进而影响土壤微生物群落组成,此外,土壤有机质作为微生物的能量来源和代谢底物,其含量的降低亦会减少土壤真菌的代谢底物,抑制微生物的生命活动,改变土壤微生物群落结构[35]。土壤酶活性能够有效反映土壤微生物的代谢状态与活力,对维持土壤肥力、有机物质循环以及农作物生长发育具有重要影响。本研究中,土壤有机质含量与毛霉门、捕虫霉亚门和被孢霉门的相对丰度呈显著或极显著正相关,与壶菌门和球囊菌门的相对丰度呈显著负相关,说明土壤有机质含量的降低会促进壶菌门、球囊菌门微生物的生长繁殖,抑制毛霉门、捕虫霉亚门和被孢霉门微生物的生长繁殖。土壤酸性磷酸酶活性和蔗糖酶活性与担子菌门和球囊菌门的相对丰度呈极显著正相关,说明土壤酸性磷酸酶活性和蔗糖酶活性的降低会抑制担子菌门和球囊菌门微生物的生长繁殖。此外,本研究还发现,土壤pH值降低会抑制担子菌门微生物的生长繁殖;碱解氮含量降低会抑制油壶菌门微生物的生长繁殖;全磷含量的升高会促进子囊菌门微生物的生长繁殖;N/P和C/N的降低会促进子囊菌门的生长繁殖;土壤pH值、过氧化氢酶活性和酸性磷酸酶活性的降低会促进油壶菌门微生物的生长繁殖;C/P的降低会促进子囊菌门微生物的生长繁殖。
综上,设施辣椒土壤真菌群落结构会受到土壤pH值、有机质含量、养分含量、生态化学计量特征、酶活性等环境因子的直接或间接影响,而导致设施辣椒连作障碍的主要原因则可能是设施辣椒连作改变了土壤理化性质,降低了土壤酶活性,影响了土壤真菌群落结构,进而破坏了土壤微生态平衡。
参考文献:
[1]李崇玮,柏新富,陈国忠,等. 不同恢复年限老参地土壤养分以及酚酸类代谢物含量差异[J]. 植物生态学报,2021,45(11):1263-1274.
[2]李昕竹,贡 璐,唐军虎,等. 塔里木盆地北缘绿洲不同连作年限棉田土壤有机碳组分特征及其与理化因子的相关性[J]. 环境科学,2022,43(10):4639-4647.
[3]Tayyab M,Yang Z Q,Zhang C F,et al. Sugarcane monoculture drives microbial community composition,activity and abundance of agricultural-related microorganisms[J]. Environmental Science and Pollution Research International,2021,28(35):48080-48096.
[4]黄菊英,崔 东,刘玉珊,等. 丹参根际微生物和连作障碍的研究进展[J]. 江苏农业科学,2023,51(3):8-15.
[5]程国亭,王延峰,姜文婷,等. 设施番茄土壤障碍综合防控研究进展[J]. 中国蔬菜,2023(2):16-24.
[6]曾子凡,马 艳,罗 佳,等. 长期施肥对设施蔬菜土壤理化及生物学性状的影响研究进展[J]. 江苏农业科学,2023,51(6):9-15.
[7]岳思君,冯翠娥,杨彦研,等. 不同连作年限硒砂瓜土壤细菌群落结构特征[J]. 干旱地区农业研究,2020,38(1):230-236.
[8]郑立伟,赵阳阳,王一冰,等. 不同连作年限甜瓜种植土壤性质和微生物多样性[J]. 微生物学通报,2022,49(1):101-114.
[9]Liu Z X,Liu J J,Yu Z H,et al. Long-term continuous cropping of soybean is comparable to crop rotation in mediating microbial abundance,diversity and community composition[J]. Soil and Tillage Research,2020,197:104503.
[10]张 敏,马 淼.甘草根际土壤微生物群落对长期连作的响应[J]. 生态学报,2022,42(22):9017-9025.
[11]李 莹,刘兰英,姜宇杰,等. 辣椒连作对土壤细菌群落的影响[J]. 微生物学报,2023,63(1):297-318.
[12]徐 晗,鄢紫薇,覃卫林,等. 生物质炭调控辣椒连作障碍研究进展[J]. 江苏农业科学,2023,51(6):23-31.
[13]鲍士旦. 土壤农化分析[M]. 3版.北京:中国农业出版社,2000.
[14]关松荫. 土壤酶及其研究法[M]. 北京:农业出版社,1986.
[15]韩海蓉,李 屹,陈来生,等. 设施辣椒连作对土壤理化性状、酶活性及微生物区系的影响[J]. 中国土壤与肥料,2021(3):237-242.
[16]Sun K N,Fu L Y,Song Y,et al. Effects of continuous cucumber cropping on crop quality and soil fungal community[J]. Environmental Monitoring and Assessment,2021,193(7):436.
[17]杜杏蓉,李运国,邓小鹏,等. 连作对不同类型植烟土壤化学性状、酶活性及细菌群落的影响[J]. 中国烟草科学,2021,42(5):30-35.
[18]陈晓婷,王裕华,林立文,等. 连作百香果对土壤理化性质和微生物特性的影响及病原真菌的分离与鉴定[J]. 热带作物学报,2021,42(2):495-502.
[19]孙 倩,吴宏亮,陈 阜,等. 不同轮作模式下作物根际土壤养分及真菌群落组成特征[J]. 环境科学,2020,41(10):4682-4689.
[20]Pervaiz Z H,Iqbal J,Zhang Q M,et al. Continuous cropping alters multiple biotic and abiotic indicators of soil health[J]. Soil Systems,2020,4(4):59.
[21]Li W H,Liu Q Z,Chen P. Effect of long-term continuous cropping of strawberry on soil bacterial community structure and diversity[J]. Journal of Integrative Agriculture,2018,17(11):2570-2582.
[22]杨 阳,李海亮,马凯丽,等. 连作对党参根际土壤理化性质、微生物活性及群落特征的影响[J]. 环境科学,2023,44(11):6387-6398.
[23]Li Y,Chi J L,Ao J,et al. Effects of different continuous cropping years on bacterial community and diversity of cucumber rhizosphere soil in sold2b6dc7ecaaa4d2a8e595efa1ebc77511cd0ff6924e5058883d21c0fe6733d54ar-greenhouse[J]. Current Microbiology,2021,78(6):2380-2390.
[24]Li Y,Wu J S,Liu S L,et al. Is the C∶N∶P stoichiometry in soil and soil microbial biomass related to the landscape and land use in southern subtropical China?[J]. Global Biogeochemical Cycles,2012,26(4):GB4002.
[25]王 棋,徐传涛,王昌全,等. 烤烟连作对土壤生态化学计量特征的影响[J]. 农业资源与环境学报,2020,37(5):702-708.
[26]朱长伟,孟威威,石 柯,等. 不同轮耕模式下小麦各生育时期土壤养分及酶活性变化特征[J]. 中国农业科学,2022,55(21):4237-4251.
[27]余 高,陈 芬,王 海.有机肥替代化肥对土壤养分、酶活性及太子参产质量的影响[J]. 北方园艺,2023(14):79-86.
[28]洪 彪,李 明,黄结雯,等. 连作对广藿香扦插苗土壤微生物及酶活性的影响[J]. 江西农业大学学报,2020,42(3):438-447.
[29]侯 乾,王万兴,李广存,等. 马铃薯连作障碍研究进展[J]. 作物杂志,2019(6):1-7.
[30]刘姣姣,何 静,陈 伟,等. 花椒连作对土壤化学性质及酶活性的影响[J]. 分子植物育种,2019,17(22):7545-7550.
[31]Zhang L Y,Xiang Y Z,Jing Y M,et al. Biochar amendment effects on the activities of soil carbon,nitrogen,and phosphorus hydrolytic enzymes:a meta-analysis[J]. Environmental Science and Pollution Research International,2019,26(22):22990-23001.
[32]XuZ W,Zhang T Y,Wang S Z,et al. Soil pH and C/N ratio determines spatial variations in soil microbial communities and enzymatic activities of the agricultural ecosystems in Northeast China: Jilin Province case[J]. Applied Soil Ecology,2020,155:103629.
[33]PangZ Q,Tayyab M,Kong C B,et al. Continuous sugarcane planting negatively impacts soil microbial community structure,soil fertility,and sugarcane agronomic parameters[J]. Microorganisms,2021,9(10):2008.
[34]常芳娟,张贵云,张丽萍,等. 生物熏蒸配施微生物菌剂对西瓜连作土壤真菌群落结构的影响[J]. 中国生态农业学报(中英文),2022,30(2):248-257.
[35]Jin L F,Guo D Y,Ning D Y,et al. Covering the trees of Kinokuni tangerine with plastic film during fruit ripening improves sweetness and alters the metabolism of cell wall components[J]. Acta Physiologiae Plantarum,2018,40(10):182.
