火焰处理对巫山植烟土壤微生物多样性及代谢特征的影响
2024-11-07刘建军曹安全杨再军彭玖华王利祥陈和青张丰收薛刚








摘要:通过火焰消毒机械在旋耕的过程中利用燃烧燃料的方式把深度15 cm土壤旋起来与400 ℃的火焰直接接触且维持2~3 s,以探明火焰处理对巫山植烟土壤微生物多样性及代谢特征的影响。采用随机区组设计,共设置3个处理:CK为对照,不对土壤作任何处理;T1处理为火焰处理1次;T2处理为火焰处理连续2次;重复6次。结果表明:(1)各样本间细菌和真菌特有ASV(amplicon sequence variant)个数均具有明显差异;(2)与对照相比,火焰处理对土壤细菌和真菌的丰度和多样性影响均不显著,但随着火焰处理次数的增加样本会出现分离;(3)T1处理增加了变形菌门细菌、鞘氨醇单胞菌属细菌和担子菌门、壶菌门、球囊菌门、白环蘑属、粗糙孔菌属及斜盖伞属真菌的相对丰度,降低了酸杆菌门细菌和子囊菌门、被孢霉门真菌及被孢霉属、腐质霉属、小脆柄菇属真菌的相对丰度;(4)T2处理增加了拟杆菌门细菌、黄杆菌属细菌和子囊菌门、担子菌门、壶菌门及白环蘑属、Echria属、奈瑟氏菌属真菌的相对丰度,降低了芽单胞菌门细菌、被孢霉门真菌和被孢霉属、镰刀菌属、小脆柄菇属真菌的相对丰度;(5)不同处理土壤细菌在目水平到属水平上,分别有5、8、14组差异物种出现显著差异(P<0.05),不同处理土壤真菌在纲水平到属水平上,分别有1、1、3、3组差异物种出现显著差异(P<0.05);(6)T2处理土壤伯克氏菌属、黏菌属、全噬菌属细菌相对丰度较高或显著较高,T1处理和T2处理土壤真菌重要性前2的附球菌属、裂壳菌属的相对丰度均较高,其中T2处理均显著较高(P<0.05);(7)T1 vs. CK、T2 vs. CK、T2 vs. T1比对组的差异代谢物分别有77、52、33种;(8)T1 vs. CK 的比对组中有4种显著差异代谢物及7条显著差异代谢通路,T2 vs. CK、T2 vs. T1和T2 vs. T1 vs. CK 的比对组中均有6种显著差异代谢物,显著差异代谢通路分别有19、15、17条。综合而言,火焰处理能显著影响土壤微生物的群落结构组成、差异物种和物种重要性,同时随着火焰处理次数的增加差异代谢物数量减少,而显著差异代谢物数量及代谢通路条数增加。
关键词:火焰处理;微生物群落结构;差异物种;差异代谢物;代谢通路
中图分类号:S572.04;S572.061 文献标志码:A
文章编号:1002-1302(2024)18-0232-19
收稿日期:2023-11-13
基金项目:中国烟草总公司重庆市公司科技计划(编号:B2021NY1309)。
作者简介:刘建军(1986—),男,重庆巫山人,硕士,农艺师,主要从事烟草栽培工作。E-mail:ljjun86@163.com。
通信作者:薛 刚,博士,讲师,主要从事烟草栽培、烘烤及分级工作。E-mail:xuegangyan2009@sina.corn。
巫山作为全国烤烟八大香型之一醇甜香的代表产地、全国核心产区,巫山烤烟入选《中国大百科全书》第2版网络版中国原产地物产目录。而由于巫山地形复杂,土地资源有限,烤烟常年连作加之施肥、种植制度、耕作模式等不合理,导致烟田土壤板结、酸化,土传病害严重,特别是青枯病和黑胫病,进而导致烟叶生长发育受到限制,尤其是在烤烟后期上部叶采烤时发生严重,叶片枯萎,为减低烟农损失,采取上部叶抢采的紧急措施,但由于叶片成熟不够,烤后烟叶易烤青、挂灰,影响烟叶品质,同时,产量降低,烟农收入减少,限制了巫山地区烟草产业的可持续发展。
优质烟叶生产与生态环境和土壤相关,同时还受土壤微生物和土壤代谢物的影响。土bd218bdb3f51f775948dccc2ccd249c836c6da1eb3c55ab8822511c8a8390246壤的重要组成包括微生物[1-5],它是土壤中最活跃的份子[6],土壤微生物群落的组成和相对丰度对土壤中有机残体及有机质的分解转化起着积极的作用。土壤中同时存在有益和有害微生物,它们相互影响,当土壤有益微生物相对丰度降低处于阈值以下时,作物生长受到影响。因此,前人对土壤消毒进行了大量研究。目前广泛采用的土壤消毒技术有生物方法[7-12]、化学方法[13-21]和物理方法[22-29]。其中物理消毒技术对土壤生态破坏最小,同时减少农药的施用,有利于降低作物农残,对环境和食品安全有显著优势。其中火焰燃烧和我国焚烧秸秆效果相似,都是通过物理燃烧释放大量的热,将田间土壤中的有害微生物、草籽、害虫和挥发性有害物质消除,达到土壤生态重构的目的,以适应烟草种植。
烟草连作致使土传病害发生,一方面是因为连作导致土壤病原菌大量富集,使其相对丰度显著升高,另一方面是因为连作导致土壤代谢物异常,使其化感自毒作用发挥主要作用。有机酸、直链醇、酚酸、萜类和生物碱等代谢物多数属于自毒物质[30],其中萜类和酚酸类物质是导致土壤化感自毒作用的关键代谢物[31]。作物长期连作后土壤代谢物苯甲酸、羟基苯甲酸和肉桂酸等酚酸类物质与土壤微生物互作而不断富集,严重影响土壤有机质的分解矿化[32-35],导致化感自毒作用加剧。
为缓解连作对烤烟产量和品质的影响,前人通过生物方法和化学方法研究较多,但对于物理方法的火焰处理鲜有报道。火焰处理在我国的芹菜[36]、水稻[37]、黄瓜[38]、草莓[39]、甘薯[40]等作物已有应用。火焰处理是通过高温使生物蛋白质发生变性,进而致使生物细胞失活,最终导致生物死亡。因此,火焰处理可为烟草绿色优质生产提供技术支撑。本试验通过火焰处理研究其对巫山植烟土壤微生物多样性和代谢特征的影响,为减轻巫山土传病害和烤烟优质绿色生产提供技术依据。
1 材料与方法
1.1 试验地概况
试验于2023 年4月16日在重庆市巫山县三溪乡月池村开展。试验地位于110.14°E、31.15°N,海拔977 m,地貌地形为山地平地,土壤为黄壤土,耕层深度为30 cm,烟草连作年限16年。试验田集中连片、肥力中等,土壤pH值为6.6,有机质含量为19.40 g/kg,全氮含量为1.38 g/kg,碱解氮含量为85.80 mg/kg,有效磷含量为 16.00 mg/kg,速效钾含量为208.00 mg/kg。
1.2 试验材料
火焰消毒机械(实用新型专利:一种土壤高温消毒装置,专利号:ZL202222878061.0)。
1.3 试验设计
采用随机区组设计,共设置3个处理:CK,作为对照,不对土壤作任何处理;T1处理,火焰处理1次;T2处理,火焰处理连续2次。重复6次,每重复3行,每行面积为20.70 m2。
火焰消毒机械在旋耕的过程中利用燃烧燃料的方式把深度15 cm土壤旋起来与400 ℃的火焰直接接触且维持2~3 s。
1.4 土壤样品的采集与处理
火焰消毒机械处理后30 min时取土样。每个处理取6个点,用取土器取15 cm土层土壤样品,过 0.85 mm孔径的筛,取 5 g土壤装入10 mL离心管,并立即放入盛有20 kg干冰的干冰桶中,并于当天邮寄到上海欧易生物科技有限公司和上海鹿明生物科技有限公司,分别测定土壤微生物和土壤代谢物,公司收到土壤样品时,干冰有大量剩余,土壤中微生物不受后续检测影响。
1.5 测定项目及方法
1.5.1 土壤微生物群落高通量测序
采用16S进行细菌测序,测序区域为16S V3~V4区,343F-798R为测序引物[41];采用ITS进行真菌测序,测序区域为ITS1区,ITS1F-ITS2R作为测序引物[42]。
1.5.2 非靶向代谢组学测定土壤代谢物
采用液相色谱-质谱联用(LC-MS)对土壤生物进行非靶向代谢组学研究。
1.6 数据处理
利用Origin 8.505448fee051e56138dfbefd1fe15162c、QIIME2 2020.11、PyNAST 0.1、R 3.5.1、PICRUSt 2 2.3.0b0、KronaTools 2.7.1、LEfSe 1.0.0等软件进行土壤微生物的聚类分析、多样性分析、NMDS分析、差异分析和随机森林分析并制图。
2 结果与分析
2.1 火焰处理对植烟土壤微生物的影响
2.1.1 火焰处理对土壤微生物测序数据和ASV聚类分析的影响
统计各样本的序列数、序列长度等参数并对数据进行统计分析。对于细菌,高通量测序得到78 108~81 654对原始序列,44 845~62 213条高质量序列,41 617~56 170 条有效序列,平均产生52 737.5条有序序列,各样本ASV数量分布在 449~1 607之间。对于真菌,高通量测序得到78 363~81 960对原始测序列,42 531~77 432条高质量序列,42 524~77 222 条有效序列,平均产生70 237.5条有序序列,各样本 ASV 数量分布在65~337之间。根据ASV聚类分析结果绘制成花瓣图(flower plot)(图1),细菌和真菌群落花瓣图的中心Core为所有样本共有ASV个数。
由图1-a可以看出,各样本间细菌特有ASV个数具有明显差异,18个样本共有ASV数为2个,其中T1_1特有ASV个数最多,为1 605个,T2_6特有ASV个数最少,为447个。由图1-b可以看出,各样本间真菌特有ASV个数具有明显差异,18个样本共有ASV数为4个,其中T1_1特有ASV个数最多,为333个,T1_3特有ASV个数最少,为61个。由图1-c可以看出,与对照相比,火焰处理1次的土壤细菌和真菌的ASV数量均增加,而火焰处理连续2次的土壤细菌和真菌的ASV数量均减少。
2.1.2 火焰处理对土壤微生物丰度和多样性的影响
经过高通量测序,根据100%的相似度阈值对细菌和真菌群落进行聚类判定与物种注释,共计得到9 196个细菌ASV和1 565个真菌ASV。其中,细菌隶属于36门86纲200目313科601属1 216 种;真菌隶属于15门37纲78目148科252属268种。
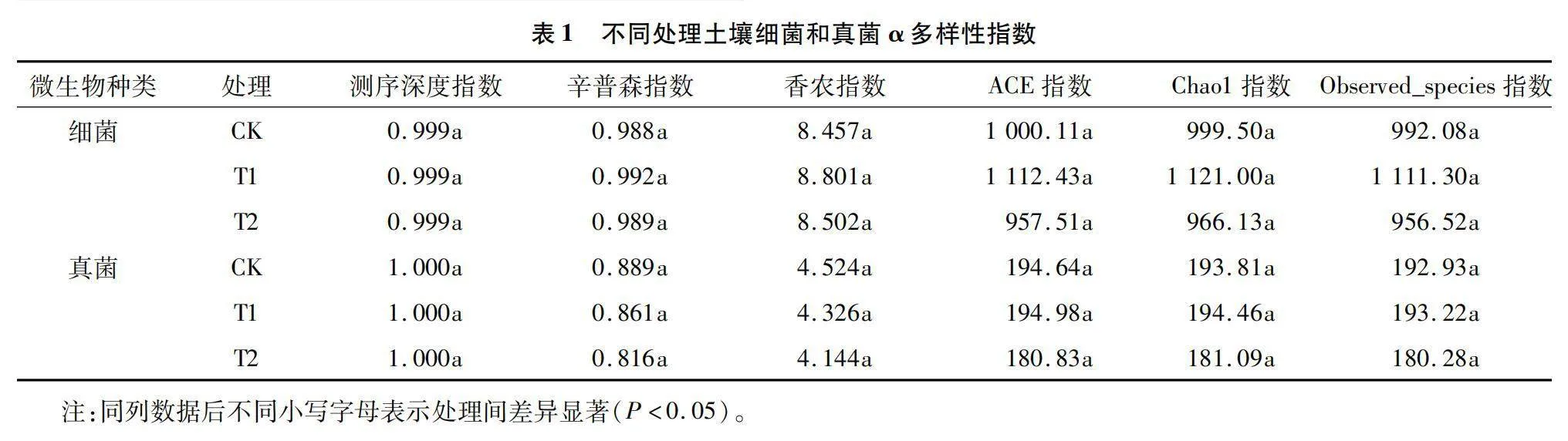
由表1可以看出,细菌和真菌不同处理的测序深度指数均高于0.99,说明所有处理的测序数据能反映样本中细菌和真菌群落结构的多样性。与对照相比,不同火焰处理的辛普森指数、香农指数、ACE指数、Chao1指数和Observed_species指数等均无显著差异,说明所有处理土壤细菌和真菌丰度、多样性无显著变化,火焰处理对土壤细菌和真菌的丰度和多样性影响不显著。
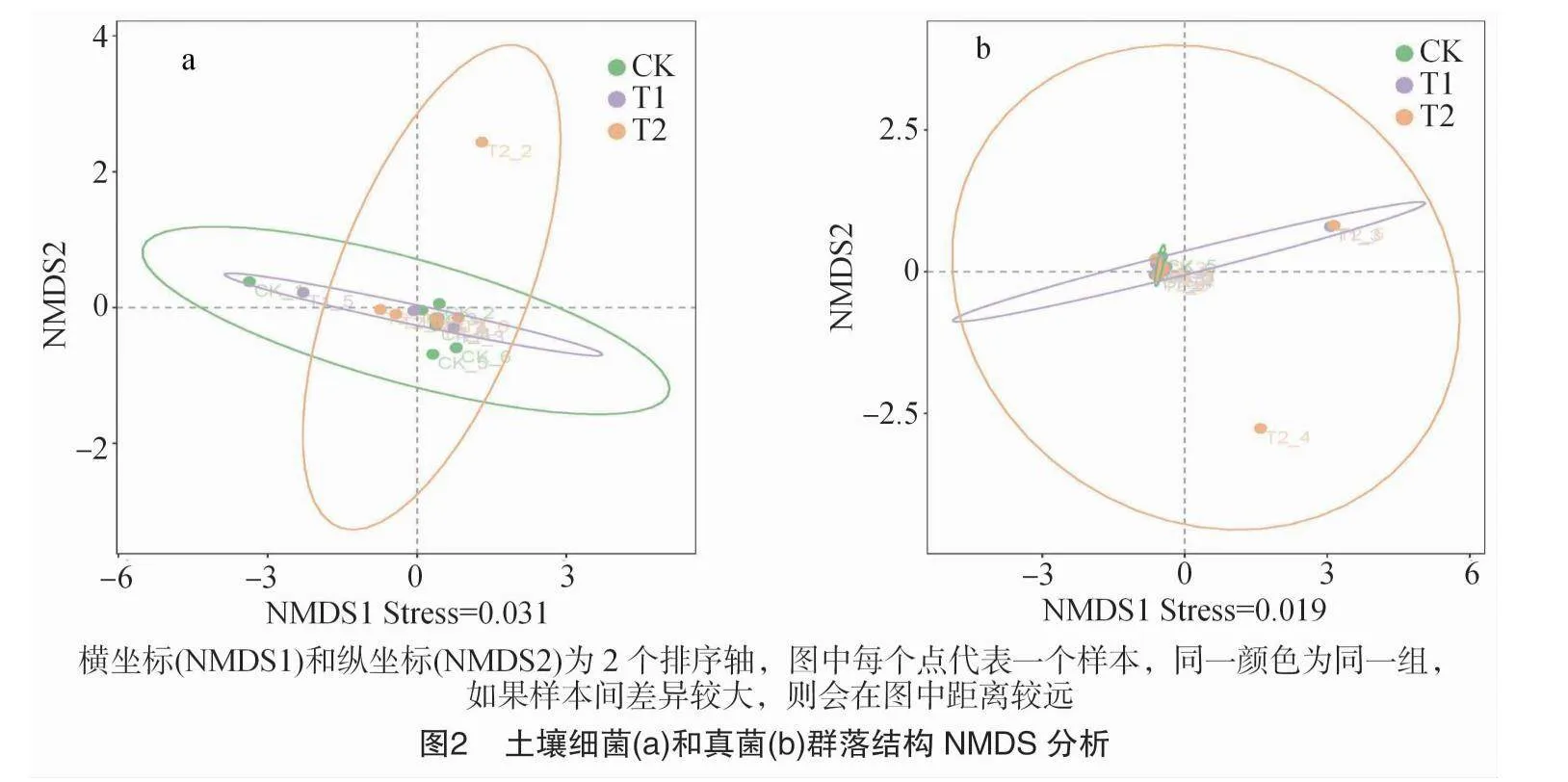
2.1.3 火焰处理下土壤微生物群落结构的NMDS分析
由图2-a和图2-b可以看出,土壤细菌和真菌的Stress(应力值)分别为0.031、0.019,说明样品对火焰处理下土壤细菌和真菌具有实际解释意义,且具有很好的代表性。与对照相比,对于细菌,火焰处理1次的T1处理所有样本会富集到一起,随着火焰处理次数的增加样本会出现分离的情况;对于真菌,随着火焰处理次数的增加,样本出现分离的情况更加明显。
2.1.4 火焰处理对土壤微生物门水平群落组成的影响
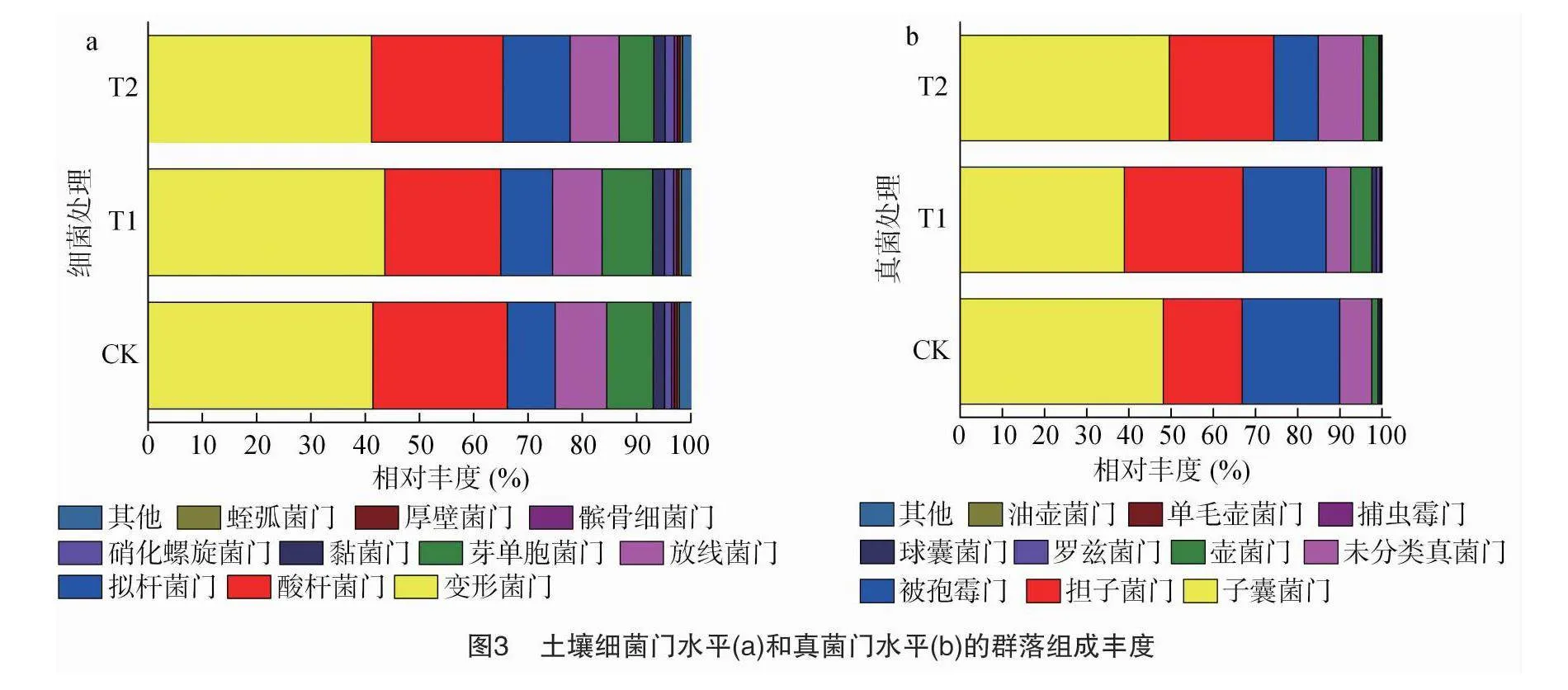
由图3-a可以看出,不同处理土壤细菌门水平群落组成相对丰度大于1.00%的有变形菌门、酸杆菌门、拟杆菌门、放线菌门、芽单胞菌门、黏菌门、硝化螺旋菌门7种,其中变形菌门、酸杆菌门是CK和T1处理土壤中优势细菌门(相对丰度>10.00%),两者相对丰度之和分别占到了总测序丰度的 66.18%和64.96%;T2处理土壤中变形菌门、酸杆菌门和拟杆菌门为优势细菌门,三者相对丰度之和占到了总测序丰度的 77.74%。与对照相比,T1处理土壤中酸杆菌门细菌相对丰度降低,变形菌门细菌相对丰度增加,T2处理土壤中芽单胞菌门细菌相对丰度降低,拟杆菌门细菌相对丰度增加。
由图3-b可以看出,CK、T2处理土壤真菌门水平群落组成相对丰度大于1.00%的有子囊菌门、担子菌门、被孢霉门、未分类真菌门、壶菌门5种,T1处理有6种,增加了球囊菌门;其中子囊菌门、担子菌门、被孢霉门为CK和T1处理的优势真菌门(相对丰度>10.00%),三者相对丰度之和分别占到了总测序丰度的 89.97%和86.74%,而子囊菌门、担子菌门、被孢霉门、未分类真菌门为T2处理土壤中优势真菌门,四者相对丰度之和占到了总测序丰度的 95.49%。与对照相比,T1处理子囊菌门、被孢霉门和未分类真菌门真菌相对丰度降低,担子菌门、壶菌门和球囊菌门真菌相对丰度增加;T2处理土壤中被孢霉门的相对丰度降低,子囊菌门、担子菌门、未分类真菌门和壶菌门真菌相对丰度增加。
2.1.5 火焰处理对土壤微生物属水平群落组成的影响
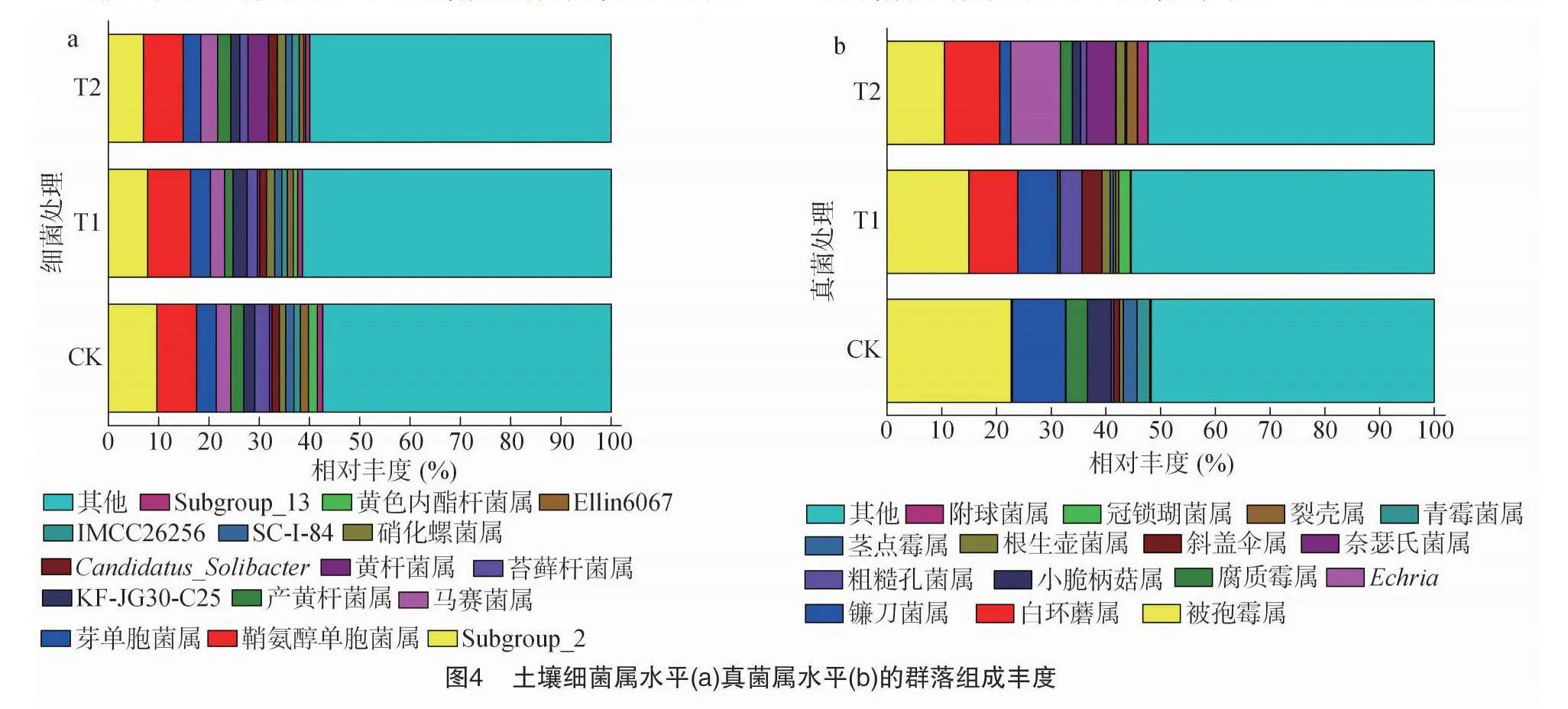
由图4-a可以看出,不同处理土壤优势细菌属(相对丰度>5.00%)为Subgroup_2和鞘氨醇单胞菌属。与对照相比,不同火焰处理Subgroup_2的相对丰度均降低,T1处理土壤中鞘氨醇单胞菌属细菌相对丰度升高,T2处理土壤中黄杆菌属细菌相对丰度升高。
由图4-b可以看出,与对照相比,T1处理土壤中白环蘑属、粗糙孔菌属和斜盖伞属真菌相对丰度明显增加,被孢霉属、镰刀菌属、腐质霉属、小脆柄菇属、茎点霉属和青霉菌属真菌相对丰度明显降低;T2处理土壤中白环蘑属、Echria和奈瑟氏菌属的相对丰度明显增加,被孢霉属、镰刀菌属、小脆柄菇属和青霉菌属真菌的相对丰度明显降低。
镰刀菌属真菌是烟草根腐病致病菌,亦是自然界中常见的与作物的生长密切相关的病原菌,在生产上烟草根腐病与黑胫病常常混合发生。由图4-b可以看出,镰刀菌属真菌相对丰度随火焰处理次数的增加而快速降低,说明火焰处理可以降低土壤根腐病致病菌相对丰度,尤其是T2处理更为明显。
2.1.6 火焰处理对土壤微生物差异物种的影响
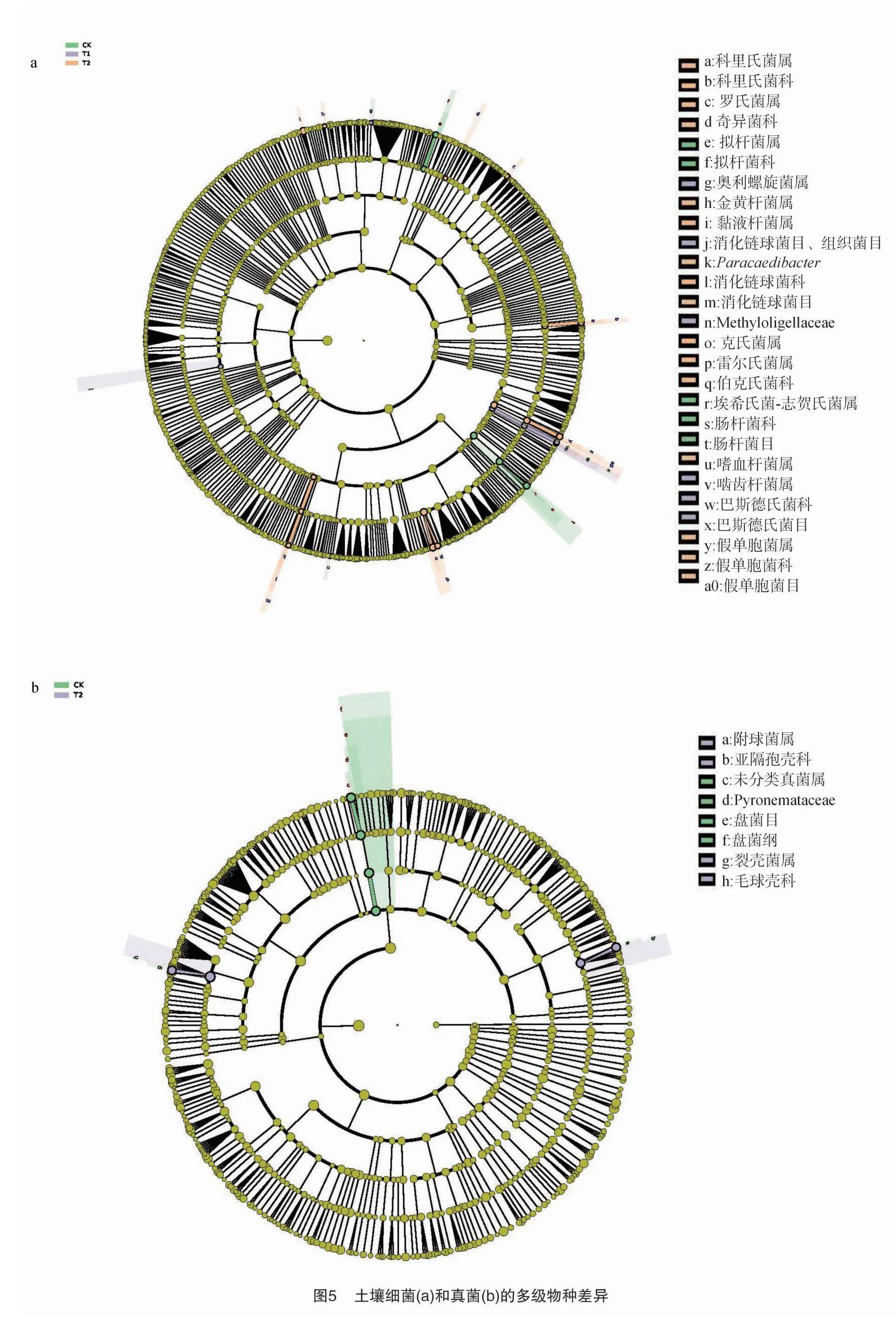
微生物多元变量统计分析采用Kruskal Wallis算法进行,并进行Lefse多级物种差异判别分析(图 5),比较从门到属水平微生物种类的显著性。由图5-a可以看出,不同处理土壤细菌从门水平到纲水平差异物种均不显著,在目水平到属水平上,分别有5、8、14组差异物种出现显著差异(P<0.05),其中CK的差异物种显著较高的有1目2科2属,T1处理的差异物种显著较高的有2目1科3属,T2处理的差异物种显著较高的有2目5科9属。由图5-b可以看出,不同处理土壤真菌在门水平差异物种均不显著,在纲水平到属水平上,分别有1、1、3、3组差异物种出现显著差异(P<0.05),其中CK的差异物种显著较高的有1纲1目1科1属,T1处理的差异物种与其他2个处理差异均不显著,T2处理的差异物种显著较高的有2科2属。
2.1.7 火焰处理对土壤微生物物种重要性的影响
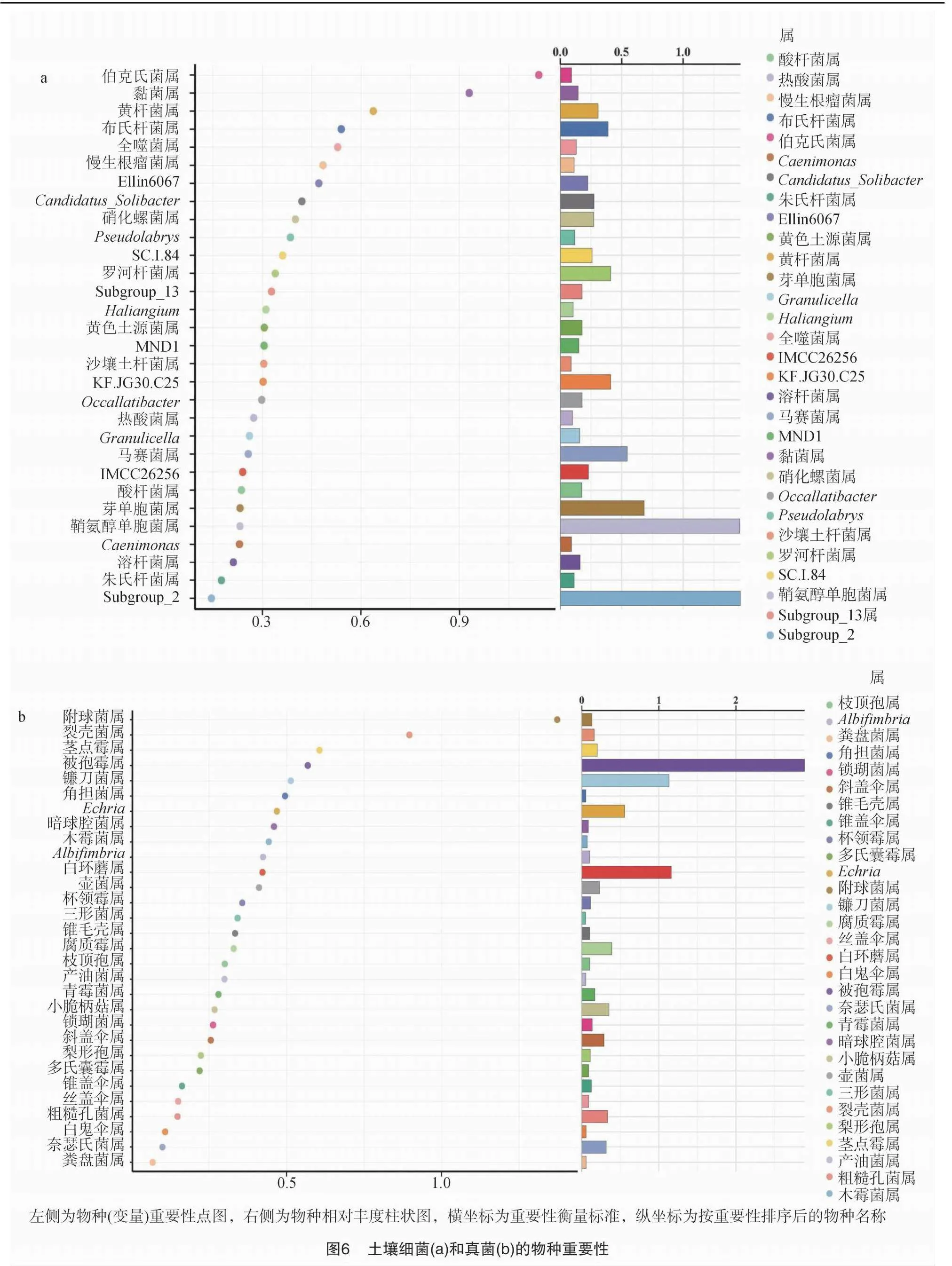
采用随机森林机器学习算法找出组间差异物种,取相对丰度前30的属绘制物种重要性点图(图6)。由图6-a可以看出,土壤细菌中前5的重要物种依次是伯克氏菌属、黏菌属、黄杆菌属、布氏杆菌属和全噬菌属。由图6-b可以看出,土壤真菌中前5的重要物种依次是附球菌属、裂壳菌属、茎点霉属、被孢霉属和镰刀菌属。
由图7-a可以看出,与对照相比,火焰处理连续2次的T2处理土壤细菌第1重要的伯克氏菌属相对丰度显著较高(P<0.05),第2重要的黏菌属较高但差异不显著,第3重要的黄杆菌属细菌差异不大,第4重要的布氏杆菌属细菌相对丰度差异不显著,第5重要的全噬菌属细菌相对丰度差异不显著且高于其他2处理。由图7-b可以看出,与对照相比,火焰处理(T1和T2处理)土壤真菌重要性前2的附球菌属、裂壳菌属真菌相对丰度均较高,其中T2处理均显著较高(P<0.05);随火焰处理次数的增加,土壤真菌重要性第3至第5的茎点霉属、被孢霉属和镰刀菌属真菌相对丰度均逐渐降低,其中T2处理茎点霉属、被孢霉属真菌相对丰度均显著低于对照(P<0.05)且T1与T2处理间差异不显著,T2处理镰刀菌属真菌相对丰度显著低于对照和T1处理(P<0.05),而T1与CK间差异不显著。
2.2 火焰处理对植烟土壤代谢物的影响
2.2.1 火焰处理对土壤差异代谢物数量的影响
不同处理筛选出的代谢物种类共2 943种,其中正离子模式下有1 808种,负离子模式下有1 135种。
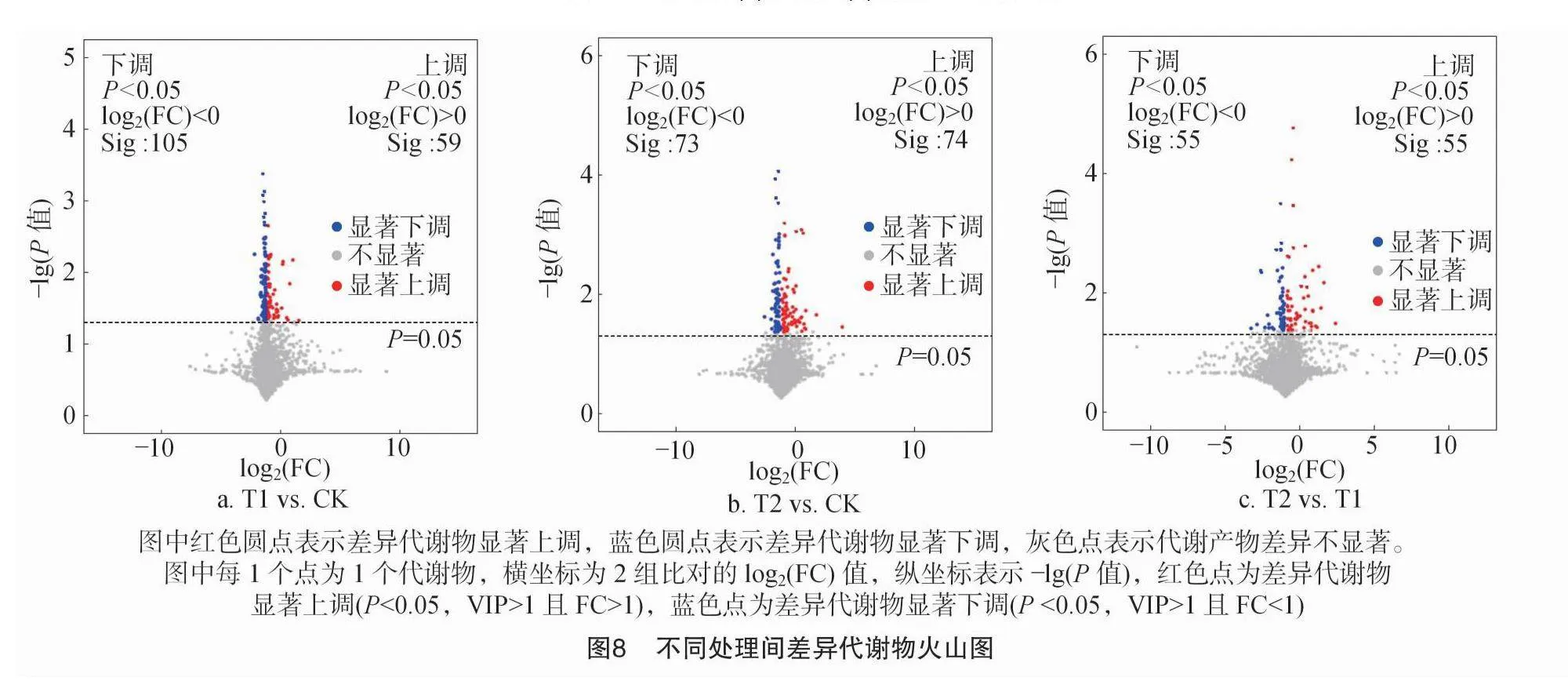
由图8可知,T1 vs. CK比对组的差异代谢物有77种,其中上调即T1处理代谢物含量高于CK的有25种,下调即T1处理代谢物含量低于CK的有52种;T2 vs. CK比对组的差异代谢物有45种,其中上调即T2处理代谢物含量高于CK的有20种,下调即T2处理代谢物含量低于CK的有25种;T2 vs. T1比对组的差异代谢物有33种,其中上调即T2处理代谢物含量高于T1处理的有11种,下调即T2处理代谢物含量低于T1处理的有22种;不同处理间差异代谢物上调和下调的数量趋势一致,均表现为T1 vs. CK>T2 vs. CK>T2 vs. T1。
2.2.2 火焰处理对土壤差异代谢物种类的影响
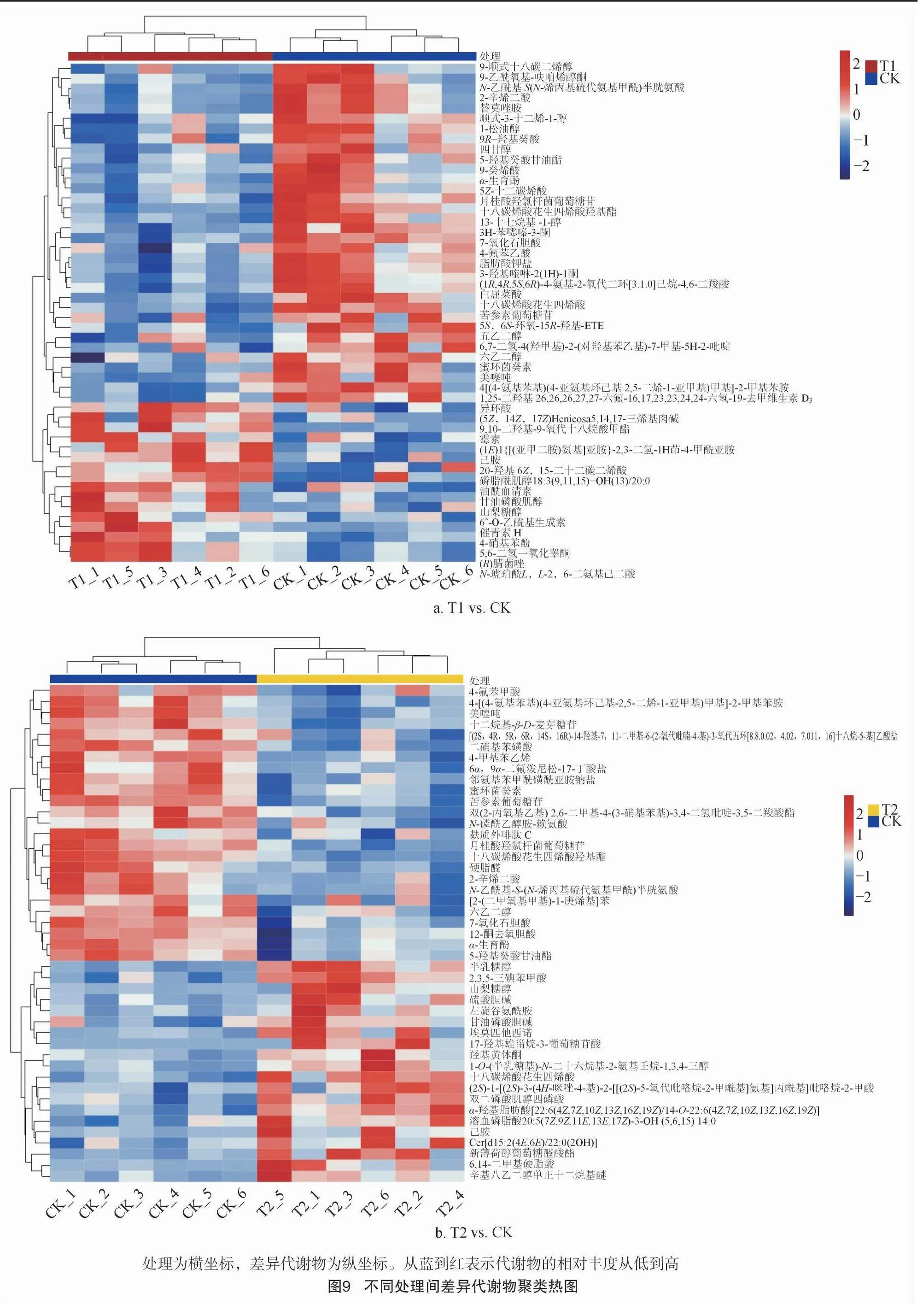
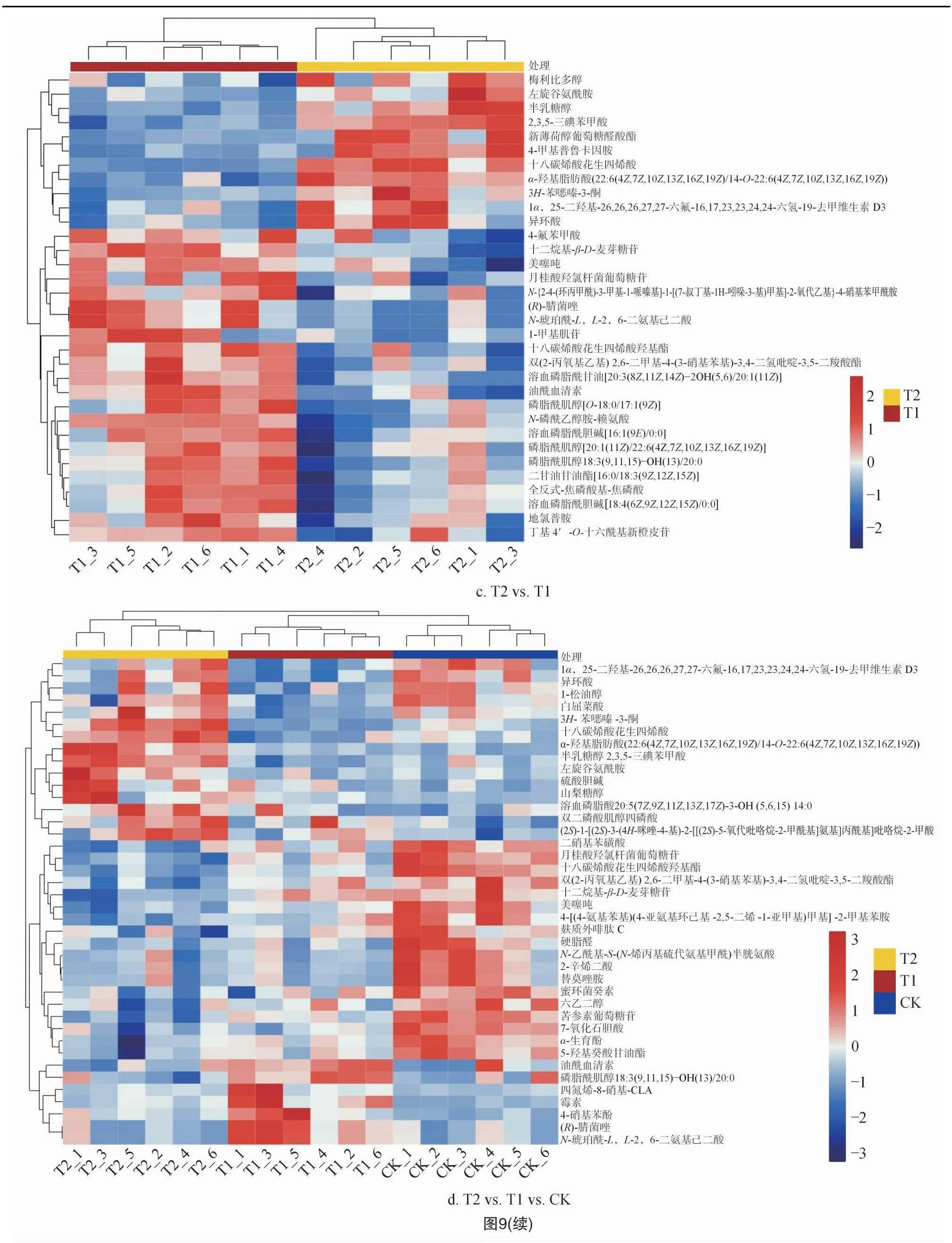
层次聚类结果表明,处理之间差异代谢物均聚为2类,一类为上调,一类为下调。由图9-a及表2可知,在T1 vs. CK 中25种上调的差异代谢物有11种脂质和类脂分子、4种苯环化合物、4种有机氧化物、3种有机酸及其衍生物、1种有机氮化合物、1种苯丙素类和聚酮类化合物和1种未分类化合物,52种下调的差异代谢物有26种脂质和类脂分子、4种苯环化合物、5种有机氧化物、4种有机酸及其衍生物、7种有机杂环化合物、4种苯丙素类和聚酮类化合物和2种未分类化合物。由图9-b及表2可知,在T2 vs. CK 中20种上调的差异代谢物有9种脂质和类脂分子、1种苯环化合物、5种机氧化物、3种有机酸及其衍生物、1种有机氮化合物和1种未分类化合物,25种下调的差异代谢物有12种脂质和类脂分子、5种苯环化合物、1种有机氧化物、2种有机酸及其衍生物、2种有机杂环化合物、2种苯丙素类和聚酮类化合物和1种未分类化合物。由图9-c及表2可知,在T2 vs. T1 中11种上调的差异代谢物有6种脂质和类脂分子、2种苯环化合物、1种有机氧化物、1种有机酸及其衍生物、1种有机杂环化合物,22种下调的差异代谢物有11种脂质和类脂分子,4种苯环化合物,2种有机杂环化合物,1种有机酸及其衍生物,1种核苷、核苷酸及类似物和3种未分类化合物。由图9-d及表2可知,在T2 vs. T1 vs. CK 中15种上调的差异代谢物有5种脂质和类脂分子、3种有机氧化物、3种有机酸及其衍生物、2种有机杂环化合物、1种苯环化合物和1种未分类化合物,25种下调的差异代谢物有11种脂质和类脂分子、4种苯环化合物、3种有机酸及其衍生物、3种有机杂环化合物、1种有机氧化物、1种苯丙素类和聚酮类化合物、2种未分类化合物。
2.2.3 火焰处理对土壤差异代谢物代谢通路及富集的影响
基于KEGG数据库对不同处理差异代谢物以P≤0.05为阈值进行代谢通路富集分析。代谢通路中P表示该代谢通路富集的显著性,且P越小,该代谢通路差异越显著。
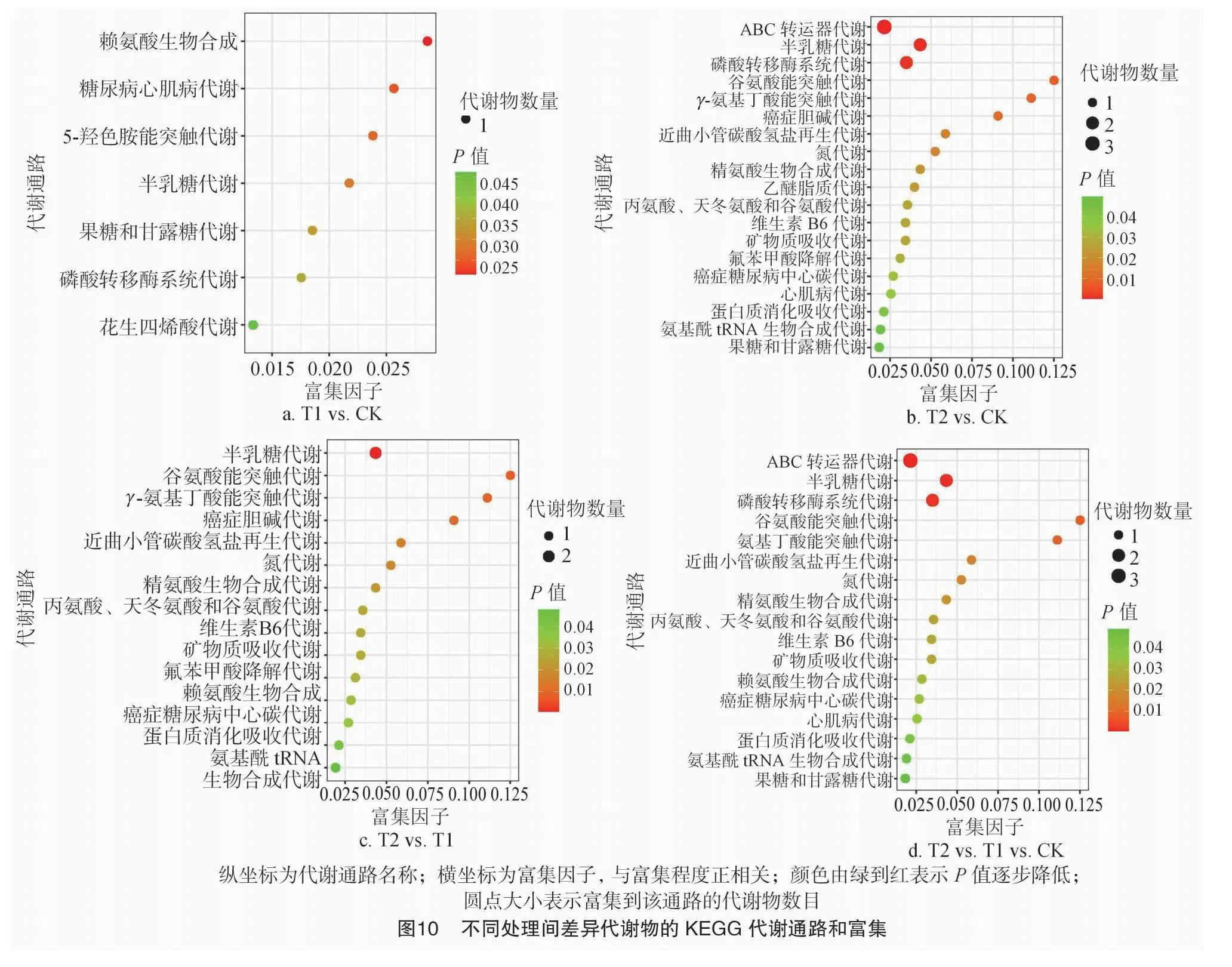
由图10-a和表3可知,T1 vs. CK 的比对组中,显著差异代谢物共有4种且均为上调即T1处理差异代谢物含量高于CK,分别为N-琥珀酰-L,L-2,6-二氨基庚二酸酯、山梨醇、4-硝基苯酚和5,6-二氢一氧化睾酮,这4种显著差异代谢物分别参与1、5、1、2条共9条代谢通路,其中有7条代谢通路具有显著差异,分别为赖氨酸生物合成代谢、糖尿病心肌病代谢、5-羟色胺能突触代谢、半乳糖代谢、果糖和甘露糖代谢、磷酸转移酶系统代谢和花生四烯酸代谢。
由图10-b和表3可知,T2 vs. CK 的比对组中,显著差异代谢物共有6种且5种上调1种下调,5种上调的代谢物分别为甘油磷酸胆碱、硫酸胆碱、半乳糖醇、山梨醇和左旋谷氨酰胺,1种下调的代谢物为4-氟苯甲酸,这6种显著差异代谢物共参与25条代谢通路,其中有19条代谢通路具有显著差异,甘油磷酸胆碱参与了癌症胆碱代谢和醚脂代谢2条代谢通路,硫酸胆碱参与了ABC转运器代谢1条代谢通路,半乳糖醇参与了半乳糖代谢和磷酸转移酶系统代谢2条代谢通路,山梨醇参与了ABC转运器代谢、半乳糖代谢、磷酸转移酶系统代谢、糖尿病心肌病代谢和果糖和甘露糖代谢5条代谢通路,4-氟苯甲酸参与了氟苯甲酸降解代谢1条代谢通路,左旋谷氨酰胺参与了ABC转运器代谢、谷氨酸能突触代谢、γ-氨基丁酸能突触代谢、近曲小管碳酸氢盐再生代谢、氮代谢、精氨酸生物合成代谢、丙氨酸、天冬氨酸和谷氨酸代谢、维生素B6代谢、矿物质吸收代谢、癌症糖尿病中心碳代谢、蛋白质消化吸收代谢和氨基酰-tRNA 生物合成代谢12天代谢通路。
由图10-c和表3可知,T2 vs. T1 的比对组中,显著差异代谢物共有6种且3种上调3种下调,3种上调的代谢物分别为半乳糖醇、梅利比多醇和左旋谷氨酰胺,3种下调的代谢物分别为溶血磷脂酰胆碱[18∶4(6Z,9Z,12Z,15Z)/0∶0]、N-琥珀酰-L,L-2,6-二氨基庚二酸酯和4-氟苯甲酸,这6种显著差异代谢物共参与23条代谢通路,其中有15条代谢通路具有显著差异,半乳糖醇和梅利比多醇均参与了半乳糖代谢且均为1条代谢通路,左旋谷氨酰胺参与了谷氨酸能突触代谢、γ-氨基丁酸能突触代谢、近曲小管碳酸氢盐再生代谢、氮代谢、精氨酸生物合成、丙氨酸、天冬氨酸和谷氨酸代谢、维生素B6代谢、矿物质吸收代谢、癌症糖尿病中心碳代谢、蛋白质消化吸收代谢和氨基酰-tRNA 生物合成代谢11条代谢通路,溶血磷脂酰胆碱[18∶4(6Z,9Z,12Z,15Z)/0∶0]参与了癌症胆碱代谢1条代谢通路,N-琥珀酰-L,L-2,6-二氨基庚二酸酯参与了赖氨酸生物合成代谢1条代谢通路,4-氟苯甲酸参与了氟苯甲酸降解代谢1条代谢通路。
由图10-d和表3可知,T2 vs. T1 vs. CK 的比对组中,显著差异代谢物共有6种,分别为硫酸胆碱、半乳糖醇、N-琥珀酰-L,L-2,6-二氨基庚二酸酯、山梨醇、4-硝基苯酚和左旋谷氨酰胺,这6种显著差异代谢物共参与23条代谢通路,其中有17条代谢通路具有显著差异,硫酸胆碱参与了ABC转运器代谢1条代谢通路,半乳糖醇参与了半乳糖代谢和磷酸转移酶系统代谢2条代谢通路,N-琥珀酰-L,L-2,6-二氨基庚二酸酯参与了赖氨酸生物合成代谢1条代谢通路,山梨醇参与了ABC转运器代谢、半乳糖代谢、磷酸转移酶系统代谢、糖尿病心肌病代谢和果糖和甘露糖代谢5条代谢通路,左旋谷氨酰胺参与了ABC转运器代谢、谷氨酸能突触代谢、γ-氨基丁酸能突触代谢、近曲小管碳酸氢盐再生代谢、氮代谢、精氨酸生物合成代谢、丙氨酸、天冬氨酸和谷氨酸代谢,维生素B6代谢,矿物质吸收代谢,癌症糖尿病中心碳代谢,蛋白质消化吸收代谢和氨基酰-tRNA生物合成代谢12条代谢通路,而4-硝基苯酚代谢通路差异不显著。
3 结论与讨论
3.1 讨论
土壤微生物多样性越高,土壤生态系统越稳定,土壤越健康[43]。娄俊鑫等研究表明,火焰处理对土壤真菌α多样性指数无影响不显著,但对真菌丰度和真菌群落结构有显著影响[28]。本试验研究表明,各样本间细菌和真菌特有ASV个数均具有明显差异,而火焰处理对土壤细菌和真菌的丰度和α多样性影响均不显著,这说明虽然火焰与土壤直接接触,但并没有破坏土壤微生物多样性,不会打破土壤生态平衡。火焰处理下土壤微生物群落结构的NMDS分析表明,随着火焰处理次数的增加,样本会出现分离的情况,这说明火焰处理对土壤细菌和真菌群落组成有显著影响。这可能是因为火焰处理使微生物群落内部种群发生了变化,打破原有平衡,种群丰度改变,从而造成群落差异。
与对照相比,火焰处理1次的T1处理变形菌门细菌的相对丰度增加,酸杆菌门细菌的相对丰度降低;火焰处理连续2次的T2处理拟杆菌门细菌的相对丰度增加,芽单胞菌门细菌的相对丰度降低。变形菌门细菌主要参与复合降解[44];酸杆菌门细菌对植物残体降解有重要作用[45],属寡营养类群,在低营养环境中的丰度反而更高[46],且具有嗜酸性,其丰度与土壤pH值呈显著负相关[47];拟杆菌门细菌属于富营养类群且对碳源非常敏感,可以利用土壤中的速效碳源快速生长[48];芽单胞菌门细菌对土壤质量和环境稳定性具有重要影响。这说明火焰处理促进有机物质分解,增加土壤中碳源,增强土壤肥力,富营养型拟杆菌门细菌丰度升高,而寡营养型酸杆菌门细菌相对丰度降低,同时变形菌门细菌相对丰度升高,土壤复合降解能力增强,由于火焰与土壤直接接触,芽单胞菌门细菌相对丰度降低,对土壤生态稳定性有一定影响。
与对照相比,T1处理子囊菌门、被孢霉菌门真菌的相对丰度降低,担子菌门、壶菌门和球囊菌门真菌的相对丰度增加;T2处理被孢霉门真菌的相对丰度降低,子囊菌门、担子菌门、壶菌门真菌的相对丰度增加。子囊菌门真菌能加快有机物代谢,促进土壤碳循环[49],随着火焰处理次数的增加,土壤中碳源增加,促进土壤子囊菌门真菌繁殖,加速土壤碳循环;被孢霉菌门真菌中较多的属为植物病害的主要属[50],火焰处理降低了被孢霉菌门真菌相对丰度降低且随火焰处理的增加其丰度显著降低;担子菌门真菌能与植物共生形成菌根,有利于作物根系发育,壶菌门真菌能释放酵素分解土壤中的腐烂的有机物,本试验中火焰处理增加了担子菌门真菌和壶菌门真菌的相对丰度,有利于增加土壤养分和后期烟草根系发育;球囊菌门真菌通过与作物根系共生形成丛枝菌根,有助于作物根系吸收无机盐,尤其是磷,火焰处理1次的T1处理增加了土壤球囊菌门真菌相对丰度,有利于后期烟草根系吸收土壤养分。
与对照相比,火焰处理1次的T1处理鞘氨醇单胞菌属细菌的相对丰度升高,火焰处理2次的T2处理黄杆菌属细菌的相对丰度升高。鞘氨醇单胞菌属细菌可降解芳香化合物[51],有利于降解除草剂和农药等有毒物质[52];黄杆菌属有机营养型好氧菌,具有降解复合有机物和生物聚合物的能力[53]。说明火焰处理能增加鞘氨醇单胞菌属细菌和黄杆菌属细菌的相对丰度,增强土壤对有毒物质的降解能力。
与对照相比,T1处理白环蘑属、粗糙孔菌属和斜盖伞属真菌的相对丰度增加,被孢霉属、腐质霉属和小脆柄菇属真菌的相对丰度降低;T2处理的白环蘑属、Echria和奈瑟氏菌属真菌的相对丰度增加,被孢霉属、镰刀菌属和小脆柄菇属真菌的相对丰度降低。白环蘑属真菌可与植物形成外生菌根[54],本试验中火焰处理增加了白环蘑属真菌的相对丰度,利于后期烟株根系对土壤养分的吸收;粗糙孔菌属真菌是有益菌,具有免疫调节生物学活性,本试验中火焰处理1次增加了土壤粗糙孔菌属真菌相对丰度,能增加植物抗病性;斜盖伞属真菌能与植株建立共生关系,有助于植株更好地吸收和利用营养,特别是钾元素,促进其生长发育,同时斜盖伞属真菌的子实体对某些癌症具有抑制作用,本试验火焰处理1次的T1处理能促进斜盖伞属真菌相对丰度增加,说明短时间的高温处理有利于斜盖伞属真菌生长繁殖;被孢霉属真菌可促进土壤碳循环,通过释放有机酸来溶解难利用的磷,并且可以加强植物对病原体的抵抗能力,促进植物生长[55],J6VvfdyPJfjEKSmPjvy05Q==本试验中被孢霉属真菌相对丰度均降低,且随火焰处理次数的增加,其相对丰度依次降低,这与娄俊鑫等研究的中档火力火焰处理能显著提高被孢霉属真菌相对丰度[28]的结论不一致,而与高档火力火焰处理能显著降低被孢霉属真菌相对丰度的结论一致,可能是因为本试验的火焰处理温度和娄俊鑫等研究的高档火力火焰温度相近所致。小脆柄菇属真菌是一种与植物密切共生的微生物,增强植物免疫能力和抗病性,腐质霉属真菌能分解有机物质,产生生物碱、腐殖酸、脂肪酸等有用化合物,对土壤肥力的提高和植物的生长具有重要作用,火焰处理后有机残体炭化减少,土壤中含糖物质降低,不利于被孢霉属真菌、小脆柄菇属真菌和腐质霉属真菌繁殖,他们相对丰度均降低;镰刀菌属真菌是土壤主要病原菌之一,可侵染多种作物,导致作物严重减产[56],提高土壤有机质含量能降低镰刀菌属丰度[57],本试验中火焰处理2次土壤中由于有机残体炭化致使有机质含量提高,土壤镰刀菌属真菌相对丰度降低,从而降低烟草土传病害的发生。
与对照相比,火焰处理连续2次的T2处理细菌差异物种在科水平和属水平上显著较多,而真菌差异物种在纲水平至属水平各有一种物种显著差异。火焰处理连续2次的T2处理土壤细菌第1重要的伯克氏菌属相对丰度显著较高(P<0.05),第2重要的黏菌属较高但差异不显著,第3重要的黄杆菌属细菌差异不大,第4重要的布氏杆菌属细菌相对丰度差异不显著(P>0.05),第5重要的全噬菌属的相对丰度差异不显著且相对丰度高于其他2个处理。火焰处理的T1和T2处理土壤真菌重要性前2的附球菌属和裂壳菌属的相对丰度均较高,其中T2处理均显著较高(P<0.05);随火焰处理次数的增加,T1和T2处理土壤真菌重要性第3至第5的茎点霉属、被孢霉属和镰刀菌属的相对丰度均逐渐降低,其中T2处理茎点霉属和被孢霉属的相对丰度均显著低于对照(P<0.05),镰刀菌属的相对丰度显著低于对照和T1处理(P<0.05)。伯克氏菌属细菌具有生物固氮和解磷作用,有利于作物吸收氮和磷,同时可产生植物激素如吲哚乙酸和分泌抗菌物质[58];黏菌属细菌是革兰氏阴性菌,具有降解土壤污染物的作用;黄杆菌属细菌有促进植物生长的作用[59];全噬菌属细菌是酸杆菌门细菌,具有嗜酸性,在生态系统中发挥重要作用;附球菌属真菌能促进真菌次级代谢产物的产生;布氏杆菌属细菌、茎点霉属真菌和镰刀菌属真菌均为致病菌。说明火焰处理连续2次能提高伯克氏菌属细菌、黏菌属细菌、全噬菌属细菌和附球菌属真菌、裂壳菌属真菌的相对丰度,降低布氏杆菌属细菌和茎点霉属、被孢霉属、镰刀菌属真菌的相对丰度,有利于降低土壤致病菌富集,提高土壤抗病性,促进后期作物生长发育。
土壤微生物通过矿化、硝化、氨化作用影响土壤养分释放和吸收,同时不同土壤微生物的功能不同,从而影响土壤微生物代谢特征。
与对照相比,火焰处理1次的T1处理有77种差异代谢物,其中上调即T1处理代谢物含量较高的有25种,下调即T1处理代谢物含量较低的有52种;火焰处理连续2次的T2处理有45种差异代谢物,其中上调的有20种,下调的有25种;T2 vs. T1比对组的差异代谢物有33种,其中上调的有11种,下调的有22种;不同比对组中土壤代谢物均以脂质和类脂分子最多。说明随着火焰处理次数的增加,土壤差异代谢物种类数量降低,且下调的种类多于上调的种类,这可能是因为火焰处理温度较高,不利于土壤代谢物的产生。
与对照相比,T1处理显著差异代谢物共有4种且均为上调,分别为N-琥珀酰-L,L-2,6-二氨基庚二酸酯、山梨醇、4-硝基苯酚和5,6-二氢一氧化睾酮,这4种显著差异代谢物有7条显著差异的代谢通路;T2处理显著差异代谢物共有6种且5种上调1种下调,分别为甘油磷酸胆碱、硫酸胆碱、半乳糖醇、山梨醇、左旋谷氨酰胺和4-氟苯甲酸,这6种显著差异代谢物有19条显著差异的代谢通路。T2 vs. T1 的比对组中,显著差异代谢物共有6种,分别为半乳糖醇、梅利比多醇、左旋谷氨酰胺、溶血磷脂酰胆碱[18∶4(6Z,9Z,12Z,15Z)/0∶0]、N-琥珀酰-L,L-2,6-二氨基庚二酸酯和4-氟苯甲酸,这6种显著差异代谢物有15条显著差异的代谢通路。T2 vs. T1 vs. CK 的比对组中,有6种差异代谢物显著,分别为硫酸胆碱、半乳糖醇、N-琥珀酰-L,L-2,6-二氨基庚二酸酯、山梨醇、4-硝基苯酚和左旋谷氨酰胺,这6种显著差异代谢物有17条显著差异的代谢通路。随着火焰处理次数的增加,土壤显著差异代谢物种类和代谢通路均增多,且除山梨醇外,其他代谢物种类均不同,说明火焰处理在改变土壤微生物群落组成的同时也改变了土壤代谢物,且除4-氟苯甲酸下调外,其他9种代谢物均上调。
3.2 结论
火焰处理1次的T1处理增加了变形菌门细菌、鞘氨醇单胞菌属细菌和担子菌门、壶菌门、球囊菌门、白环蘑属、粗糙孔菌属及斜盖伞属真菌的相对丰度,降低了酸杆菌门细菌和子囊菌门、被孢霉菌门真菌及被孢霉属、腐质霉属、小脆柄菇属真菌的相对丰度;火焰处理连续2次的T2处理增加了拟杆菌门细菌、黄杆菌属细菌和子囊菌门、担子菌门、壶菌门及白环蘑属、Echria、奈瑟氏菌属真菌的相对丰度,降低了芽单胞菌门细菌、被孢霉菌门真菌和被孢霉属、镰刀菌属、小脆柄菇属真菌的相对丰度。
火焰处理连续2次的T2处理土壤伯克氏菌属、黏菌属、全噬菌属细菌相对丰度较高或显著较高,火焰处理的T1和T2处理土壤附球菌属、裂壳菌属真菌相对丰度均较高,其中T2处理均显著较高,随火焰处理次数的增加,土壤茎点霉属、被孢霉属和镰刀菌属真菌相对丰度均逐渐降低,其中T2处理茎点霉属、被孢霉属真菌相对丰度均显著低于对照,镰刀菌属真菌相对丰度显著低于对照和T1处理。火焰处理2次的T2处理细菌差异物种在科水平和属水平上显著较多,而真菌差异物种在纲水平至属水平各有一种物种显著差异。不同比对组中土壤代谢物均以脂质和类脂分子最多。火焰处理1次的T1处理土壤差异代谢物有77种,火焰处理连续2次的T2处理土壤差异代谢物有45种;T2 vs. T1比对组的差异代谢物有33种。T1处理有4种显著差异代谢物和7条显著差异的代谢通路;T2处理有6种显著差异代谢物和19条显著差异的代谢通路。T2 vs. T1 的比对组中,有6种显著差异代谢物和15条显著差异的代谢通路。T2 vs. T1 vs. CK 的比对组中,有6种显著差异代谢物和17条显著差异的代谢通路。
火焰处理是火焰消毒、生物防治的物理方法,通过火焰与土壤直接接触,促进有机物质分解,改变土壤微环境,在土壤微生物多样性保持相对稳定的前提下改变土壤微生物群落结构,差异代谢物增多,代谢通路增加,进而降低土传病害致病菌尤其是镰刀菌属真菌相对丰度,增加有益菌相对丰度,增强土壤降解能力,促进后期作物生长发育。
参考文献:
[1]刘艳霞,李 想,邹 焱,等. 贵州省典型植烟生态区域根际土壤微生物群落多样性[J]. 生态学报,2018,38(9):3145-3154.
[2]郭 甜,何丙辉,蒋先军,等. 紫色土区植物篱对坡面土壤微生物特性的影响[J]. 水土保持学报,2011,25(5):94-98,185.
[3]唐莉娜,张秋芳,陈顺辉. 不同有机肥与化肥配施对植烟土壤微生物群落PLFAs和烤烟品质的影响[J]. 中国烟草学报,2010,16(1):36-40.
[4]Moscatelli M C,Lagomarsino A,Marinari S,et al. Soil microbial indices as bioindicators of environmental changes in a poplar plantation[J]. Ecological Indicators,2005,5(3):171-179.
[5]Joergensen R G,Emmerling C. Methods for evaluating human impact on soil microorganisms based on their activity,biomass,and diversity in agricultural soils[J]. Journal of Plant Nutrition and Soil Science,2006,169(3):295-309.
[6]邱并生. 伊贝根际微生物[J]. 微生物学通报,2010,37(8):1252.
[7]张建鹏. 化肥减量配施微生物菌肥及土壤调理剂对重茬马铃薯生长发育和土壤质量的影响[J]. 江苏农业科学,2023,51(7):205-212.
[8]郭玉鸽,申洪涛,李丽华,等. 微生物菌肥对不同酸碱度土壤及烤烟生长和致香物质的影响[J]. 江苏农业科学,2023,51(3):54-62.
[9]崔欣格,王 瑞,赵 昊,等. 微生物菌肥对不同连作土壤及烟株生长的影响[J]. 江苏农业科学,2023,51(13):240-245.
[10]岳宏忠,张东琴,侯 栋,等. 微生物菌肥部分替代化肥对设施黄瓜产量和土壤细菌群落结构的影响[J]. 西北农林科技大学学报(自然科学版),2022,50(7):118-126,137.
[11]潘光伟,孟 南,刘华山,等. 外源菌肥对二氯喹啉酸危害下烟草根际土壤微生物量及呼吸速率的影响[J]. 河南农业科学,2014,43(11):45-48.
[12]黎妍妍,李锡宏,王 林,等. 万寿菊秸秆熏蒸对烟株根际土壤原核微生物群落的影响[J]. 烟草科技,2021,54(4):15-22.
[13]陈雅琼,刘 海,田临卿,等. 土壤消毒结合微生态修复对烤烟生长及其主要病害的影响[J]. 贵州农业科学,2023,51(2):35-41.
[14]宋雨萌,陈昊晗,王 敏,等. 土壤熏蒸剂对大白菜根肿病的防效和土壤性质的影响[J]. 中国蔬菜,2022(10):81-85.
[15]燕平梅,高贵喜,曹坳程,等. 土壤熏蒸剂对土壤硝化、反硝化作用的影响[J]. 中国生态农业学报,2014,22(4):401-407.
[16]袁会珠,齐淑华. 溴甲烷土壤熏蒸防治春季大棚黄瓜土传病害[J]. 植物保护,1997,23(5):42-43.
[17]康萍芝,田生虎,吴晓燕,等. 棉隆土壤熏蒸对设施黄瓜枯萎病的田间防效评价[J]. 宁夏农林科技,2020,61(9):16-19.
[18]张大琪,颜冬冬,方文生,等. 生物熏蒸:环境友好型土壤熏蒸技术[J]. 农药学学报,2020,22(1):11-18.
[19]贾喜霞,师桂英,吕海龙,等. 土壤熏蒸-微生物有机肥联用缓解设施茄子连作障碍的土壤生物化学效应研究[J]. 甘肃农业大学学报,2019,54(3):84-92.
[20]刘 星,张书乐,刘国锋,等. 土壤熏蒸-微生物有机肥联用对连作马铃薯生长和土壤生化性质的影响[J]. 草业学报,2015,24(3):122-133.
[21]宋兆欣,王秋霞,郭美霞,等. 二甲基二硫作为土壤熏蒸剂的效果评价[J]. 农药,2008,47(6):454-456,464.
[22]宋 悦,王生北,张毅杰,等. 土壤蒸汽消毒技术研究与应用进展[J]. 中国农机化学报,2022,43(6):199-206.
[23]陈志杰,张 锋,张淑莲,等. 太阳能消毒对温室土壤环境效应及防治黄瓜根结线虫病效果[J]. 生态学报,2009,29(12):6664-6671.
[24]胡学博,曹坳程. 太阳能消毒防KeWiEo4lKS32cvlm1AJgDA==治植物土传病害[J]. 世界农业,2001(5):44-47.
[25]何 伟,罗文芳,于镇华,等. 高温闷棚对设施蔬菜根结线虫的防治效果及土壤微生物群落结构的影响[J]. 新疆农业科学,2022,59(1):179-189.
[26]李佳川,杨瑞平,张 显. 灌水高温闷棚处理对温室连作土壤修复效果的影响[J]. 北方园艺,2016(15):172-175.
[27]耿德仁. 土壤连作障碍电处理机对日光温室土壤进行消毒技术的分析[J]. 农业技术与装备,2012(4):48-49.
[28]娄俊鑫,刘 泓,沈少君,等. 不同农艺措施对烟田土壤真菌群落结构和功能的影响[J]. 中国烟草科学,2020,41(1):38-43,55.
[29]王振跃,施 艳,刘存祥. 微波消毒对土壤中病原微生物的防治效果[J]. 河南农业科学,2010(12):92-94.
[30]张兆波,毛志泉,朱树华. 6 种酚酸类物质对平邑甜茶幼苗根系线粒体及抗氧化酶活性的影响[J]. 中国农业科学,2011,44(15):3177-3184.
[31]侯 慧,董 坤,杨智仙,等. 连作障碍发生机理研究进展[J]. 土壤,2016,48(6):1068-1076.
[32]刘晓丽,赵 娜,贾钰莹,等. 酚酸类物质在植物土壤环境中的作用[J]. 园艺与种苗,2020,40(8):56-58,63.
[33]Rao G D,Liu X X,Zha W W,et al. Metabolomics reveals variation and correlation among different tissues of olive (Olea europaea L.)[J]. Biology Open,2017,6(9):1317-1323.
[34]Arapitsas P,Corte A D,Gika H,et al. Studying the effect of storage conditions on the metabolite content of red wine using HILIC LC-MS based metabolomics[J]. Food Chemistry,2016,197:1331-1340.
[35]Vander Weide J,Medina-Meza I G,Frioni T,et al. Enhancement of fruit technological maturity and alteration of the flavonoid metabolomic profile in merlot (Vitis vinifera L.) by early mechanical leaf removal[J]. Journal of Agricultural and Food Chemistry,2018,66(37):9839-9849.
[36]许光辉,赵奇龙,高 宇. 火焰高温消毒技术防治农田土壤病虫害研究与试验[J]. 农业工程,2014,4(增刊1):52-54.
[37]林 辉,郑国璋,郭 岚. 高温处理土壤对活化磷钾要素的效果[J]. 福建农业科技,1980(2):26-27.
[38]张广旭,吕亭辉,周 娣,等. 黄瓜连作土壤高温处理对根结线虫和枯萎病的影响[J]. 植物营养与肥料学报,2019,25(6):917-924.
[39]王素素,杜国栋,吕德国. 土壤高温处理对连作草莓根系呼吸代谢及植株发育的影响[J]. 果树学报,2011,28(2):234-239.
[40]岳 瑾,杨建国,杨伍群,等. 自走式精旋火焰土壤消毒机防治甘薯茎线虫、根腐病、草害技术研究[J]. 安徽农学通报,2018,24(11):60,98.
[41]Nossa C W,Oberdorf W E,Yang L Y,et al. Design of 16S rRNA gene primers for 454 pyrosequencing of the human foregut microbiome[J]. World Journal of Gastroenterology,2010,16(33):4135-4144.
[42]Bolyen E,Rideout J R,Dillon M R,et al. Reproducible,interactive,scalable and extensible microbiome data science using QIIME 2[J]. Nature Biotechnology,2019,37:852-857.
[43]Wardle D A,Bardgett R D,Klironomos J N,et al. Ecological linkages between aboveground and belowground biota[J]. Science,2004,304(5677):1629-1633.
[44]Cottrell M T,Kirchman D L. Natural assemblages of marine proteobacteria and members of the Cytophaga-Flavobacter cluster consuming low-and high-molecular-weight dissolved organic matter[J]. Applied and Environmental Microbiology,2000,66(4):1692-1697.
[45]王光华,刘俊杰,于镇华,等. 土壤酸杆菌门细菌生态学研究进展[J]. 生物技术通报,2016,32(2):14-20.
[46]Fierer N,Bradford M A,Jackson R B. Toward an ecological classification of soil bacteria[J]. Ecology,2007,88(6):1354-1364.
[47]丁新景,黄雅丽,敬如岩,等. 基于高通量测序的黄河三角洲4种人工林土壤细菌结构及多样性研究[J]. 生态学报,2018,38(16):5857-5864.
[48]Zeng J,Liu X J,Song L,et al. Nitrogen fertilization directly affects soil bacterial diversity and indirectly affects bacterial community composition[J]. Soil Biology and Biochemistry,2016,92:41-49.
[49]张胜男,闫德仁,黄海广,等. 短期封育对科尔沁沙地榆树疏林土壤微生物群落结构的影响[J]. 生态学杂志,2020,39(9):2860-2867.
[50]刘 泽. 中国被孢霉属及近缘属的分类与分子系统发育研究[D]. 北京:北京林业大学,2020:3-6.
[51]苟 敏,曲媛媛,杨 桦,等. 鞘氨醇单胞菌:降解芳香化合物的新型微生物资源[J]. 应用与环境生物学报,2008,14(2):276-282.
[52]胡 杰,何晓红,李大平,等. 鞘氨醇单胞菌研究进展[J]. 应用与环境生物学报,2007,13(3):431-437.
[53]Williams T J,Wilkins D,Long E,et al. The role of planktonic Flavobacteria in processing algal organic matter in coastal East Antarctica revealed using metagenomics and metaproteomics[J]. Environmental Microbiology,2013,15(5):1302-1317.
[54]刘 晴,徐 阳,张 妍,等. 基于高通量测序分析土层与间伐强度对辽东山区红松人工林真菌群落的影响[J]. 西南农业学报,2023,36(6):1234-1241.
[55]Osorio N W,Habte M. Soil phosphate desorption induced by a phosphate-solubilizing fungus[J]. Communications in Soil Science and Plant Analysis,2014,45(4):451-460.
[56]陈 懿,吴 春,李彩斌,等. 炭基肥对植烟黄壤细菌、真菌群落结构和多样性的影响[J]. 微生物学报,2020,60(4):653-666.
[57]Xu D B,Raza W,Yu G H,et al. Phytotoxicity analysis of extracts from compost and their ability to inhibit soil-borne pathogenic fungi and reduce root-knot nematodes[J]. World Journal of Microbiology and Biotechnology,2012,28(3):1193-1201.
[58]张珂飞,钟永嘉,孙丽莉,等. 植物有益伯克霍尔德氏菌的研究进展及其在农业中的应用[J]. 微生物学报,2021,61(8):2205-2218.
[59]Bhattacharyya P N,Jha D K. Plant growth-promoting rhizobacteria (PGPR):emergence in agriculture[J]. World Journal of Microbiology and Biotechnology,2012,28:1327-1350.
