不同干燥方式对鸡屎藤中酚类代谢物及抗氧化活性的影响
2024-10-15黄泽杨春亮








摘要:【目的】分析不同干燥方式处理鸡屎藤后酚类代谢物的变化和抗氧化活性差异,为鸡屎藤的开发利用提供理论依据。【方法】采用冷冻干燥和热烘干燥2种方式处理鸡屎藤,比较不同干燥方式处理鸡屎藤后酚类物质的抗氧化能力,并应用非靶向代谢组学方法对鸡屎藤酚类化合物进行定性分析,采用偏最小二乘法判别分析(PLS-DA)筛选酚类差异代谢物后,对其进行层次聚类分析和KEGG代谢通路富集分析,同时分析不同干燥方式处理后鸡屎藤差异代谢物与抗氧化活性之间的相关性。【结果】冷冻干燥和热烘干燥2种方式对鸡屎藤总酚和总黄酮含量均无显著影响(P>0.05);冷冻干燥鸡屎藤的1,1-二苯基-2-三硝基苯肼(DPPH)自由基清除能力、2,2-联氮-双(3-乙基苯并噻唑啉-6-磺酸)二铵盐(ABTS)阳离子自由基清除能力和铜离子还原能力均高于热烘干燥。采用超高效液相色谱串联傅里叶变换质谱仪对2种干燥方式处理的鸡屎藤均鉴定出99种酚类化合物,其中黄酮类54种,香豆素及其衍生物15种,肉桂酸及其衍生物13种,酚类9种,异黄酮类6种,黄酮醇类2种。主成分分析和层次聚类分析结果表明不同干燥方式处理的鸡屎藤酚类代谢物具有显著差异(P<0.05,下同),且热烘干燥方式有15种差异代谢物表达量显著高于冷冻干燥方式。应用PLS-DA共筛出25种极显著差异代谢物(P<0.01),其中10种代谢物上调,15种代谢物下调,变化显著的主要有黄酮类、酚类和香豆素及其衍生物。KEGG代谢通路富集分析发现,不同干燥方式下鸡屎藤差异代谢物富集代谢通路有7条,富集程度最高的前5条通路分别为类黄酮生物合成、苯丙烷生物合成、多种次生代谢产物的生物合成1、酪氨酸代谢及二苯乙烯类、二芳基庚烷类和姜辣素生物合成。25种酚类差异代谢物与DPPH自由基清除能力和铜离子还原能力均表现出显著相关性,其中有10种酚类差异代谢物与这2个指标呈显著正相关,15种酚类差异代谢物呈显著负相关。【结论】不同干燥方式处理鸡屎藤未改变酚类化合物种类,但对酚类化合物的含量及抗氧化活性均有明显影响。鸡屎藤发挥功能活性不是单一代谢物作用,而是多种代谢物协同作用的结果。
关键词:鸡屎藤;酚类;干燥方式:代谢组学;差异代谢物;抗氧化活性
中图分类号:S567.909.2文献标志码:A文章编号:2095-1191(2024)07-1992-14
Effects of different drying methods on phenolic metabolites and antioxidant activity in Paederiascandens
HUANG Ze,YANG Chun-liang
(School of Smart Tourism,Hainan College of Software Technology,Qionghai,Hainan 571400,China)
Abstract:【Objective】The study aimed to analyze the changes in phenolic metabolites and differences in antioxidant activity of Paederia scandens after different drying methods,providing theoretical basis for the development and utiliza-tion of P.scandens.【Method】Two drying methods,freeze drying and heat drying,were used to treat P.scandens,andthe antioxidant capacity of phenolic substances in P.scandens after different drying methods was compared.Non-targeted metabolomics was applied for the qualitative analysis of phenolic compounds in P.scandens.After screening for differen-tial phenolic metabolites using partial least squares discriminant analysis(PLS-DA),hierarchical clustering analysis and KEGG metabolic pathway enrichment analysis were conducted.Additionally,the correlation between the differential me-tabolites and antioxidant activity of P.scandens after treatment with different drying methods was analyzed.【Result】Freeze drying and heat drying had no significant effect on the total phenol and total flavonoid contents of P.scandens(P>0.05).The 1,1-diphenyl-2-trinitrophenylhydrazine(DPPH)radical scavenging capacity,2,2-azino-bis(3-ethyl-benzothiazole-6-sulfonic acid)diammonium salt(ABTS)cation radical scavenging capacity and copper ion reduction ca-pacity of freeze-dried P.scandens were all higher than those of the heat-dried samples.Using ultra-high-performance li-quid chromatography coupled with Fourier transform mass spectrometry,99 phenolic compounds were identified in P.scandens treated with two drying methods,including 54 flavonoids,15 coumarins and their derivatives,13 cinnamic acids and their derivatives,9 phenolics,6 isoflavonoids,and 2 flavonols.The results of principal component analysis and hierarchical cluster analysis showed that the phenolic metabolites of P.scandens treated with different drying methods were significantly different(P<0.05,the same below).The expression levels of 15 differential metabolites in the heat drying method were significantly higher than those in the freeze drying method.A total of 25 extremely significant differen-tial metabolites were screened out using PLS-DA(P<0.01),among which 10 metabolites were up-regulated and 15 me-tabolites were down-regulated.Significant changes were mainly observed in flavonoids,phenols,and coumarins and their derivatives.KEGG metabolic pathway enrichment analysis revealed that there were 7 metabolic pathways enriched with differential metabolites of P.scandens under different drying methods.The top 5 most enriched pathways were flavonoid biosynthesis,phenylpropane biosynthesis,biosynthesis of multiple secondary metabolites tyrosine metabolism,and biosynthesis of stilbenes,diarylheptanes and gingerol.The 25 phenolic differential metabolites showed significant correla-tions with both DPPH radical scavenging capacity and copper ion reduction capacity,with 10 phenolic differential me-tabolites exhibiting significant positive correlations with the two indicators and 15 phenolic differential metabolites sho-wing significant negative correlations.【Conclusion】Different drying methods applied to P.scandens do not change the types of phenolic compounds,but they have obvious effect on the content of phenolic compounds and antioxidant activity.The functional activity of P.scandens is not the result of a single metabolite,but rather the synergistic effect of multiple metabolites.
Key words:Paederia scandens;phenolics;drying methods;metabolomics;differential metabolites;antioxidant ac-tivity
Foundation items:High-level Talent Project of Hainan Natural Science Foundation(321RC1078)
0引言
【研究意义】鸡屎藤(Paederia scandens)又名鸡矢藤,隶属于茜草科,是一种多年生藤本植物,因茎叶揉碎后有鸡屎臭味而得名,主要分布于海南、广东、云南、安徽、福建等地区(黄国凯,2017;高天元等,2020)。鸡屎藤含有丰富的酚类化合物(王星星等,2021;唐超玲等,2023),多酚是一类广泛存在于植物体内有多元酚结构的次生代谢物,具有抗氧化、抗炎、抗癌、降血脂等特性(Duarte et al.,2021;Wang et al.,2022a)。新鲜的鸡屎藤水分含量高,储藏期短,且易遭受机械损伤和微生物侵染,导致腐烂和变质,严重影响其药用和食用价值;为延长鸡屎藤储藏期,提高其利用率,可采用不同的干燥方式对鸡屎藤进行处理。目前对植物叶片常用的干燥方式主要有冷冻干燥(Antal etal.,2014;刘乐等,2022)、热烘干燥(沈风娇等,2017;徐秀苹等,2017)和自然干燥(刘若菡等,2022;徐寸发等,2022)。高温条件下,酚类化合物内的酚羟基不稳定,热加工可能会影响酚类化合物的功能性和生物利用度(Oliver et al.,2016)。因此,研究不同干燥方式对鸡屎藤酚类化合物的影响,对鸡屎藤的开发利用具有重要意义。【前人研究进展】目前,关于鸡屎藤代谢物研究主要集中在成分的分离鉴定。胡海清等(2013)先后分离鉴定出鸡屎藤环烯醚萜苷类中的6β-O-β-D-葡萄糖基鸡屎藤酸和鸡屎藤蒽醌,同时分离鉴定出鸡屎藤苷、鸡屎藤酸、鸡屎藤酸甲酯和去乙酰车叶草酸甲酯。Wu等(2013)通过高效液相色谱—电喷雾电离串联质谱仪鉴定出鸡屎藤中的去乙酰车叶草酸、京尼平苷酸、车叶草酸、车叶草苷等成分。高天元等(2020)从4种鸡屎藤中共鉴定出23种化合物,包括12种环烯醚萜苷类成分、5种奎宁酸类衍生物、5种黄酮苷类化合物及1种简单化合物金线莲苷。Liu等(2022a)从鸡屎藤中鉴定出36种酚类化合物,包括5种苯甲酸、5种羟基肉桂酸、15种黄酮醇、4种黄酮、1种异黄酮、3种黄烷酮、1种花青素、1种原花青素和1种芪萜。Zhang等(2022)采用超高效液相色谱—四极杆飞行时间质谱仪(UPLC-Q-TOF/MS)对7个产地的鸡屎藤代谢物进行研究,结果发现不同产地的化合物种类相同,但含量有显著差异,陕西、大别山和湖北的样本与其他4个地区的差异最大,江苏、云南和广西之间的差异较小。Liu等(2024)采用超高效液相色谱—三重四级杆/线性离子阱质谱联用(UPLC-ESI-QTrap-MS/MS)技术,在纯化的鸡屎藤提取物中鉴定出15种黄酮类化合物,其中山柰酚-3-O-洋槐糖苷、二氢杨梅素、木犀草素-7-O-葡萄糖苷、槲皮苷、花青素-3-O-红霉苷、黄芪苷和(-)-表没食子儿茶素等化合物具有很强的抗氧化活性。干燥能有效抑制微生物生长和延缓生化反应,方便运输和贮存,提高利用率,不同干燥方式对植物的酚类化合物有一定影响。秦丹丹等(2018)研究发现经不同方式干燥后无花果的各酚类化合物含量均有显著差异。吴雅璐(2019)研究发现红外干燥和真空干燥后花椒叶中多酚、黄酮和单宁含量相对较高。邢颖等(2020)研究发现几种干燥方式中,冷冻干燥制备的生姜叶黄酮、多酚等活性成分及抗氧化能力保留效果最佳。Akther等(2023)对不同干燥处理的木槿花进行研究,结果发现冷冻干燥处理的总酚含量最高,且抗氧化活性最强。【本研究切入点】目前,针对鸡屎藤酚类化合物研究多集中在成分测定,而关于干燥方式对鸡屎藤酚类化合物的组成研究未涉及。非靶向代谢组学技术可对生物样品中的小分子代谢产物进行定性和定量分析,能较全面地反映样品的总代谢物特征(Rambaran and Nordstrm,2021;Liu et al.,2022b),目前尚未见采用该技术研究鸡屎藤酚类化合物。【拟解决的关键问题】采用冷冻干燥法和热烘干燥法处理鸡屎藤,应用非靶向代谢组学方法对鸡屎藤酚类化合物进行定性分析,采用偏最小二乘法判别分析(PLS-DA)筛选差异性酚类化合物后,对其进行层次聚类分析和KEGG代谢通路富集分析,同时分析不同干燥方式处理后鸡屎藤差异代谢物与抗氧化活性之间的相关性,探究不同干燥方式对鸡屎藤酚类化合物及抗氧化活性的影响,为鸡屎藤的开发利用提供理论依据。
1材料与方法
1.1试验材料
鸡屎藤为产自海南省琼海市的狭序鸡屎藤,于2023年4月购自琼海市菜市场。主要试剂:色谱纯甲醇(德国CNW公司);色谱纯乙腈(美国Fisher公司);没食子酸标准品、芦丁标准品、维生素C(Vc)标准品(上海源叶生物科技有限公司)。主要仪器设备:DMM-40型胶体磨(上海秦硕化工机械设备有限公司);CenLee16X台式高速离心机(湖南湘立科学仪器有限公司);PLUS-E2-20TJ实验室级超纯水机(南京易普易达科技发展有限公司);UHPLC-Q Exactive HF-X超高效液相色谱串联傅里叶变换质谱仪(赛默飞世尔科技公司);101A型数显电热鼓风干燥箱(上海浦东荣丰科学仪器有限公司);LGJ-10NS/CG真空冷冻干燥机(北京亚星仪科科技发展有限公司);Centrifuge 5430R型高速冷冻离心机(德国Eppendorf公司);FlexA-200酶联免疫分析仪(杭州优米仪器有限公司)。
1.2试验方法
1.2.1鸡屎藤样品制备热烘干燥法制备鸡屎藤:鸡屎藤叶片洗净沥干,50℃干燥24h后,粉碎过100目筛,得到热烘干燥鸡屎藤样品(H组)。
冷冻干燥法制备鸡屎藤:鸡屎藤叶片洗净沥干,存放于-80℃冰箱中12h,冷冻干燥48 h后,粉碎过100目筛,得到冷冻干燥鸡屎藤样品(D组)。
1.2.2可溶性总酚提取液制备参考张红建等(2023)的方法,并进行适当修改,制备可溶性总酚提取液。准确称取0.2 g样品于15 mL离心管中,加入10 mL 70%甲醇浸泡,将样品置于超声清洗器中,360 W、50℃下超声30 min后,过0.22μm孔径滤膜即得。
超高效液相色谱串联傅里叶变换质谱仪测试样液制备:吸取200μL可溶性总酚提取液于1.5 mL离心管中,加入400μL溶液[乙腈∶甲醇=1∶ 含0.02 mg/mL的内标(L-2-氯苯丙氨酸)],涡旋混匀30 s后,在5℃、40 kHz的条件下超声提取30min,于-20℃下静置30 min,然后在4℃下13000×g离心15 min,将上清液转移到带内插管的进样瓶中,上机分析。质控样由等量D组和H组总酚提取液混合得到。
1.2.3总酚含量测定参照张红建等(2023)的方法进行测定。移取可溶性总酚提取液125μL或标准物质与同等体积的福林酚试剂混合反应3min,分别加入1.25 mL 20%Na2CO3,在室温且避光条件下反应30 min,过0.22μm孔径滤膜,取滤液200μL在750 nm处测吸光度。以70%甲醇溶液作为空白对照,标准物质为没食子酸(50~500μg/mL)。每个样品进行3次测定并取平均值。
1.2.4总黄酮含量测定参照张红建等(2023)的方法进行测定。移取可溶性总酚提取液500μL于2 mL离心管中,先后加入30μL 5%NaNO2溶液和30μL 10%AlCl3溶液,每次添加后摇匀并置于室温中一定时间;加入200μL 1g6x8OFqRTHdlyqQA8iukO13WQ31bLH/hGu1Uvzxub+E= mol/L NaOH溶液,并用70%甲醇溶液将混合液总体积定容至1 mL;混匀后在室温下反应30 min,用酶标仪测定其在510 nm处吸光度。以70%甲醇溶液作为空白对照,20~100μg/mL芦丁作为标准黄酮化合物,绘制标准曲线。
1.2.5抗氧化活性测定参照王露(2018)的方法并稍作修改,测定1,1-二苯基-2-三硝基苯肼(DPPH)自由基清除能力和2,2-联氮-双(3-乙基苯并噻唑啉-6-磺酸)二铵盐(ABTS)阳离子自由基清除能力。DPPH自由基清除能力:将1.2.2中的可溶性总酚提取液稀释10倍后,取60µL于96孔酶标板中,依次加入40µL DPPH溶液和150µL甲醇溶液;摇动混合并在室温条件下暗处反应30min,于517 nm处测其吸光度;以Vc为标样,在0~50µg/mL范围内绘制标准曲线,结果以每克鸡屎藤干制样品中Vc毫克数计。ABTS阳离子自由基清除能力:将1.2.2中的可溶性总酚提取液稀释10倍后,取30µL于96孔酶标板中,再加入150µLABTS稀释液,室温条件下暗处反应10 min,于734 nm处测其吸光度。以Vc为标样,在0~50µg/mL范围内绘制标准曲线,结果以每克鸡屎藤干制样品中Vc毫克数计。
参照王晓宇(2008)的方法并稍作修改,测定铜离子还原能力。将1.2.2中的可溶性总酚提取液稀释10倍后,取25µL于96孔酶标板中,依次加入50µL CuCl2、50µL新亚铜试剂溶液和50µL 1 mol/L醋酸铵缓冲液(pH 7),最后加入30µL超纯水,温室条件下反应30min,于450 nm处测其吸光度。以没食子酸为标样,在0~50µg/mL范围内绘制标准曲线,结果以每克鸡屎藤干制样品中没食子酸毫克数计。1.2.6酚类化合物种类及相对含量测定色谱条件:色谱柱为ACQUITY UPLC HSS T3(100 mm×2.1 mm,1.8µm);流动相A为95%水+5%乙腈(含0.1%甲酸),流动相B为47.5%乙腈+47.5%异丙醇+5%水(含0.1%甲酸),进样量3μL,柱温40℃。质谱条件:样品质谱信号采集采用正负离子扫描模式;扫描范围70~1050 m/z,鞘气流速50 arb,辅助气流速13 arb,毛细管温度325℃;正模式喷雾电压、负模式喷雾电压和S-Lens电压分别为3500 V、-3500 V和50 V;一级质谱分辨率和二级质谱分辨率分别为60000 Full MS和7500 MS2。
1.3统计分析
在美吉生物云平台(https://www.majorbio.com/web/www/index)上进行非靶向代谢组学数据的分析处理。原始数据通过代谢组学处理软件Progenesis QI与Human Metabolome Database(HMDB)(https://www.hmdb.ca)、Metlin(https://metlin.scripps.edu)、MassBank(http://www.massbank.jp/)等数据库及自建数据库进行比对,对酚类化合物进行定性分析。采用PLS-DA筛选酚类差异代谢物,并将筛选出的物质上传至KEGG网站,进行代谢通路富集分析、层次聚类分析及相关分析。使用SPSS 23.0进行差异显著性分析。
2结果与分析
2.1不同干燥方式对鸡屎藤总酚、总黄酮含量和抗氧化活性的影响
由表1可知,不同干燥方式对鸡屎藤总酚含量无显著影响(P>0.05,下同),2种干燥方式条件下鸡屎藤总酚含量基本一致。不同干燥方式对鸡屎藤总黄酮含量也无显著影响,热烘干燥处理的鸡屎藤总黄酮含量较冷冻干燥提高2.78%,可能是因为植物中的黄酮大多与糖结合,以黄酮苷形式存在,少部分以游离状态存在,适当的加热能使黄酮分离,导致总黄酮含量增加。冷冻干燥鸡屎藤的DPPH自由基清除能力、ABTS阳离子自由基清除能力和铜离子还原能力均高于热烘干燥,其中DPPH自由基清除能力和铜离子还原能力具有显著差异(P<0.05,下同),而ABTS阳离子自由基清除能力差异不显著,表明2种干燥方式处理的鸡屎藤对ABTS阳离子自由基清除能力相当。相较于冷冻干燥,热烘干燥鸡屎藤的抗氧化活性有所下降,表明不同干燥方式会影响鸡屎藤抗氧化能力指标。
2.2冷冻干燥和热烘干燥处理鸡屎藤的酚类化合物鉴定结果
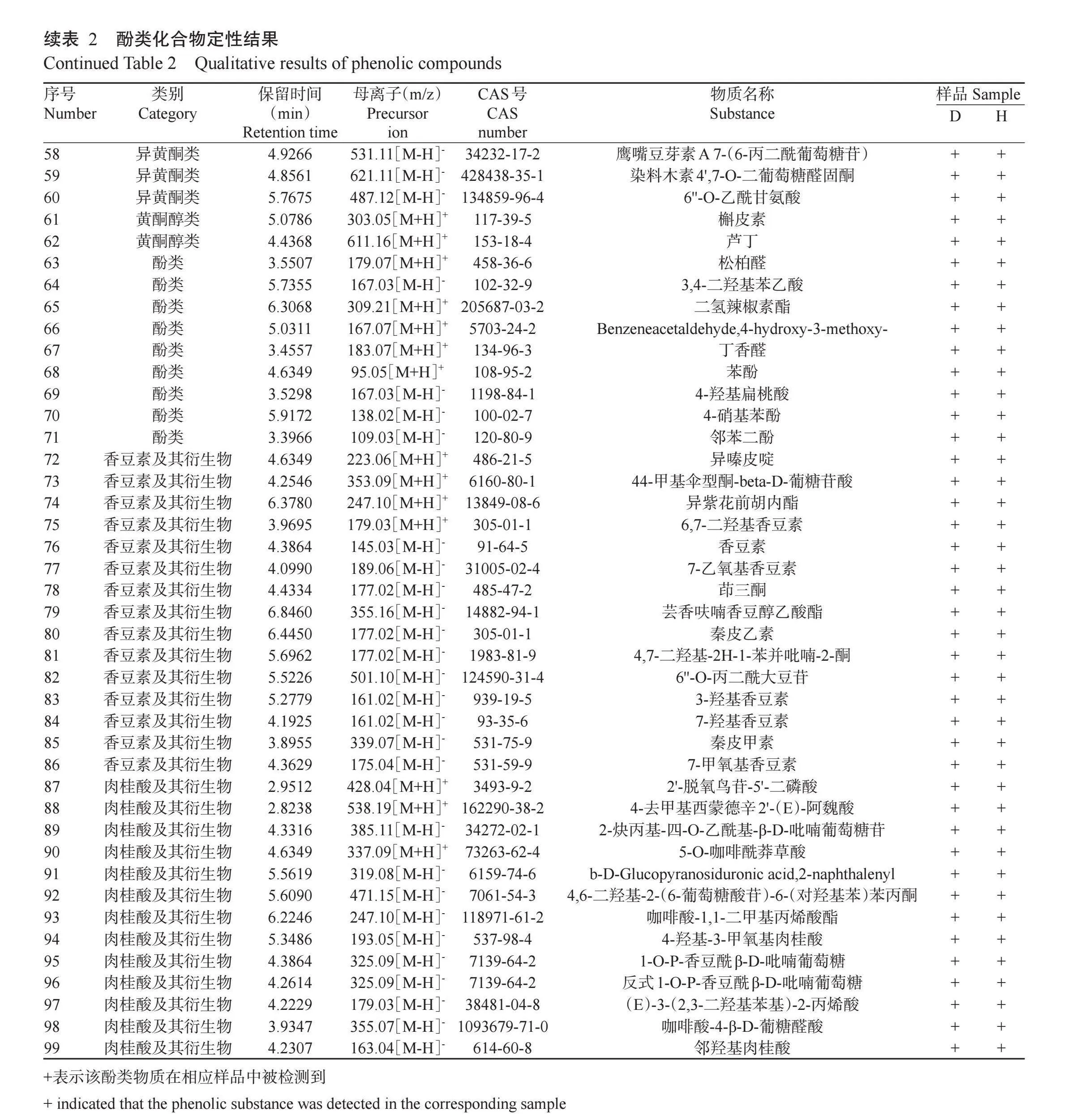
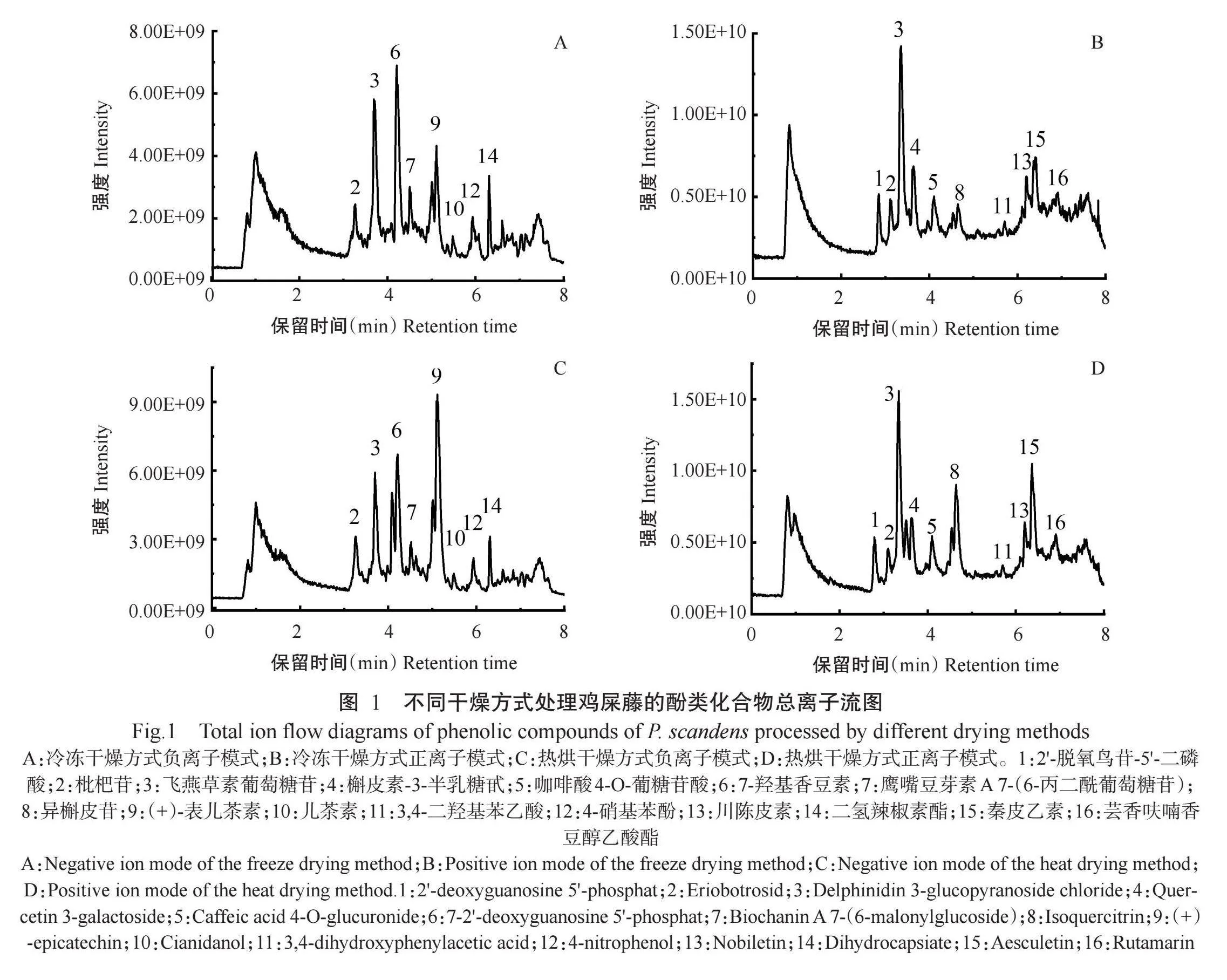
基于超高效液相色谱串联傅里叶变换质谱仪对冷冻干燥和热烘干燥处理的鸡屎藤酚类化合物进行测定,并结合HMDB、Metlin、MassBank及自建数据库对不同干燥方式的鸡屎藤酚类化合物进行定性分析。不同干燥方式处理的鸡屎藤酚类化合物总离子流见图 鉴定到的酚类化合物种类见表2。在正、负离子模式下从冷冻干燥和热烘干燥处理的鸡屎藤中均检测出99种酚类化合物,即不同干燥方式未改变鸡屎藤酚类化合物种类,说明鸡屎藤酚类化合物对温度变化具有一定的稳定性。鸡屎藤酚类化合物中,黄酮类最多,达54种,其次是香豆素及其衍生物15种,肉桂酸及其衍生物13种,酚类9种,异黄酮类6种,黄酮醇类最少,仅2种。
2.3冷冻干燥和热烘干燥处理鸡屎藤酚类化合物差异性分析结果
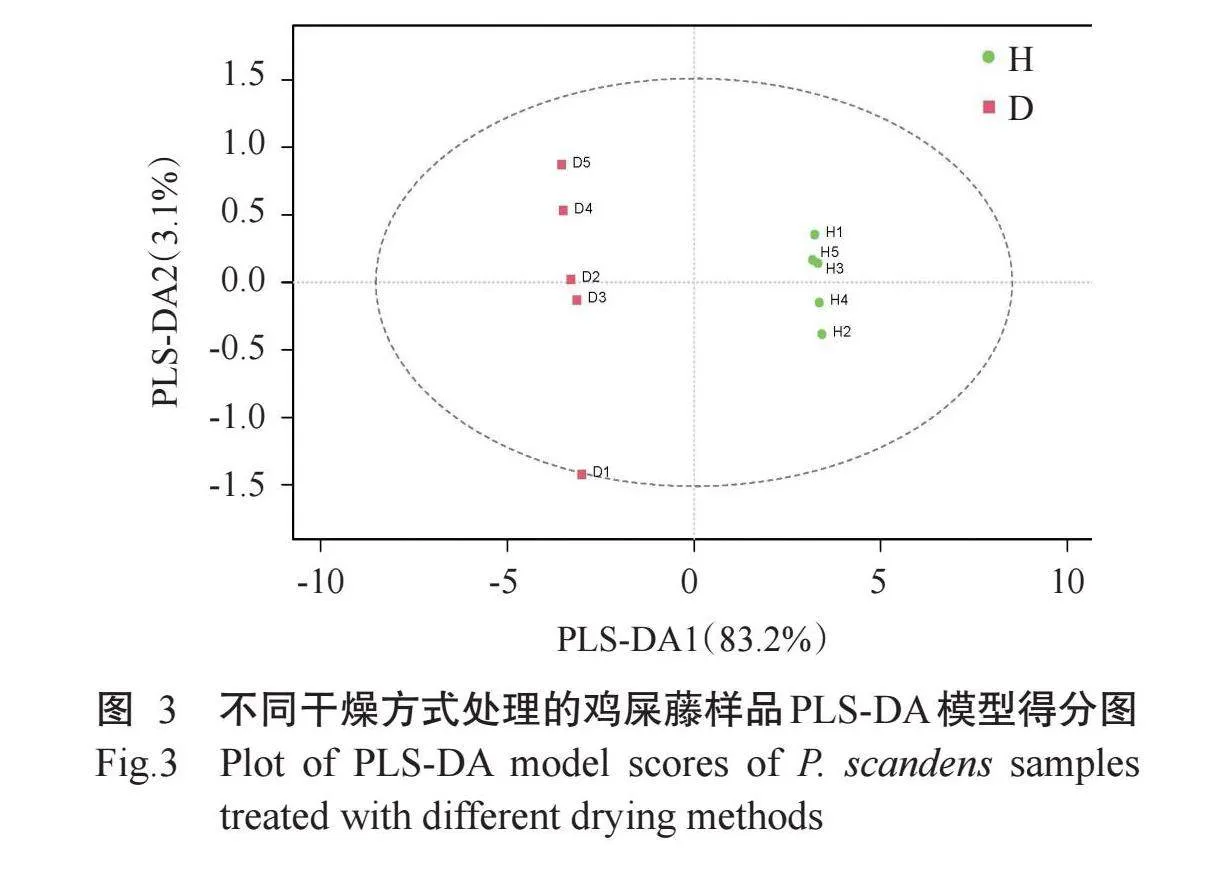
2.3.1主成分分析(PCA)结果对样品进行PCA,判别冷冻干燥和热烘干燥2种方式处理鸡屎藤各组样本之间和组内样本之间的变异度大小。由图2可知,质控样品聚集性较高,说明试验设备运行稳定、数据质量高和试验方法可靠。D组和H组样品聚集程度均较高,D组样品分布于PCA图中左侧,H组样品分布于右侧,第一主成分(PC1)的贡献率为79.07%,第二主成分(PC2)的贡献率为13.05%,二者累计贡献率达92.12%,因此,D组和H组各组内样品间整体区分度较好。D组和H组间样品差异明显,表明不同干燥方式对鸡屎藤酚类化合物具有明显影响。
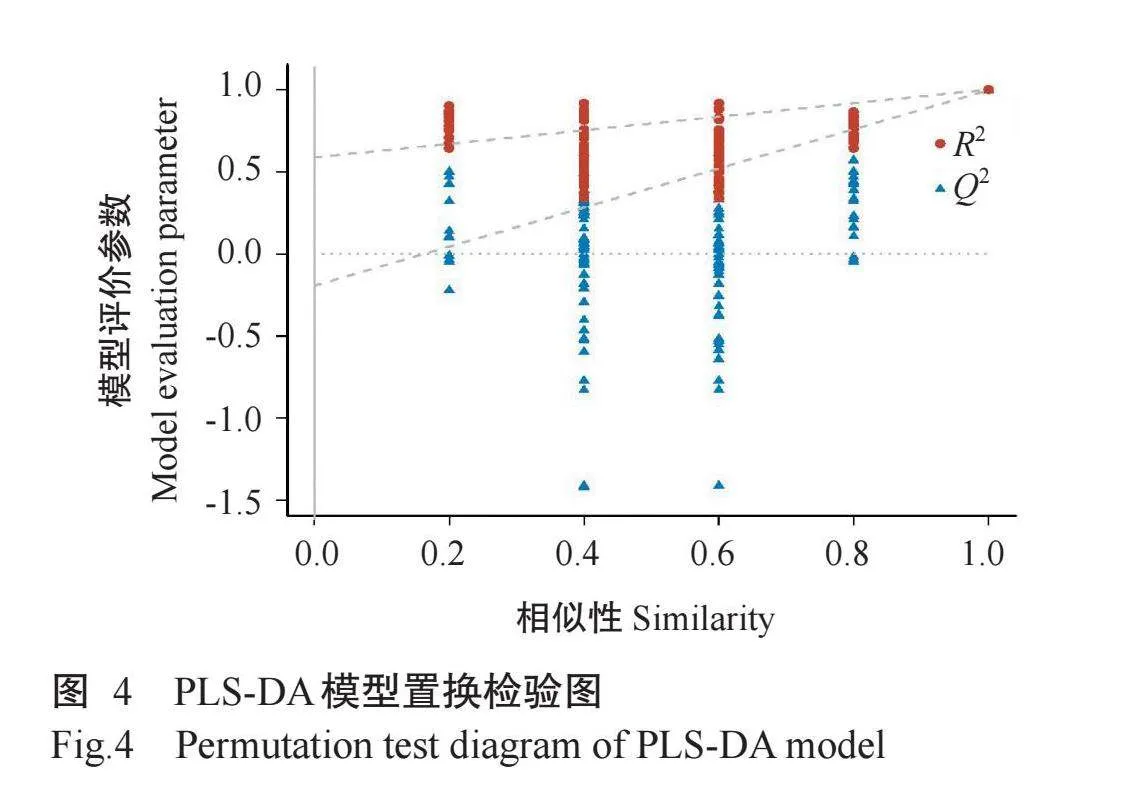
2.3.2 PLS-DA结果PCA法可对主要信息进行有效提取,但无法忽略组内误差和消除与研究目的无关的随机误差,而PLS-DA能对组间观察值进行有效区分,同时能忽略组内误差并消除与研究目的无关的随机误差。R2从0到 表示模型的拟合能力逐渐增强,而Q2代表模型的预测能力,当Q2>0.5时,表示预测能力良好,Q2>0.9时,表示预测能力极好(Jianget al.,2017)。根据PLS-DA模型对99种酚类化合物数据进行分析,由图3可知,冷冻干燥鸡屎藤样品分布在置信区间左侧,热烘干燥鸡屎藤样品分布在置信区间右侧,2组样品的区分效果非常明显。PLS-DA得到2个主成分,第一主成分(PLS-DA1)的贡献率为83.2%,第二主成分(PLS-DA2)的贡献率为3.1%,RX(2)=0.86 RY(2)=1.000,Q2=0.996,表明该模型拟合能力较强,预测能力极好。采用置换检验图对当前PLS-DA模型是否过拟合进行评估,结果如图4所示,Q2左侧所有的点均低于右侧的点,且2条回归线整体趋势向上,说明置换检验合格,该PLS-DA模型不存在过拟合现象,可用于分析样品间的代谢物差异。
2.4冷冻干燥和热烘干燥处理鸡屎藤酚类差异代谢物的层次聚类分析结果
代谢物在生物学中具有结果和功能的相互性与互补性,为直观地观察冷冻干燥和热烘干燥处理下鸡屎藤酚类化合物的含量变化,本研究采用层次聚类分析对性质相近的代谢物归为一类,性质差异较大的代谢物分为不同类(张舒等,2021)。图5为冷冻干燥和热烘干燥处理鸡屎藤后差异代谢物层次聚类分析热力及变量影响投影值(VIP)分析图。热力图的颜色代表代谢物丰度,红色是高表达组分,蓝色是低表达组分,由蓝到白再到红,代表代谢物丰度逐渐升高。图5可分成上下2个区域,上半区域是D组对H组差异代谢物表达量结果,由左侧聚类分析热图可知,D组对H组表达量有10种差异代谢物,结合右侧的VIP可明显看出,差异代谢物紫杉叶醇、异嗪皮啶、枇杷苷、6-羟基木犀草素和3,4-二羟基苯乙酸的条形图长度较长,表明此区域的差异代谢物表达量D组明显高于H组样本。下半区域是H组对D组差异代谢物表达量结果,从左侧的聚类分析热图结合右侧VIP可知,15种差异代谢物表达量均存在极显著差异(r<0.001),差异代谢物条形图长度由长到短顺序依次为二氢辣椒素酯、2'-羟基染料木素7-(6'-丙二酰糖苷)、槲皮素-3-半乳糖甙、5-O-咖啡酰莽草酸、7-乙氧基香豆素、苯酚、咖啡酸-1,1-二甲基丙烯酸酯、鹰嘴豆芽素A 7-(6-丙二酰葡萄糖苷)等,即这些差异代谢物表达量H组明显高于D组样本。通过层次聚类分析可看出,不同干燥方式对鸡屎藤酚类化合物均产生显著影响。
2.5冷冻干燥和热烘干燥处理鸡屎藤酚类差异代谢物的筛选与分析结果
以PLS-DA模型中VIP>1、差异倍数(Fold Change)>1或Fold Change<1且P<0.05为标准,对冷冻干燥和热烘干燥处理鸡屎藤中酚类差异代谢物进行筛选,结果见表3。由表3可知,2种干燥方式处理的鸡屎藤中共有25种酚类化合物发生极显著变化(P<0.01),占全部酚类物质种类的25.25%,其中包括9种黄酮类、5种酚类、5种香豆素及其衍生物、4种肉桂酸及其衍生物、2种异黄酮类。
差异代谢物中,D组样品中有10种酚类代谢物的相对含量高于H组样品,其中二氢辣椒素酯、2'-羟基染料木素7-(6'-丙二酰糖苷)、5-O-咖啡酰莽草酸、槲皮素-3-半乳糖甙、7-乙氧基香豆素、苯酚、鹰嘴豆芽素A 7-(6-丙二酰葡萄糖苷)、咖啡酸-1,1-二甲基丙烯酸酯等化合物含量显著下降,说明温度对酚类物质的稳定性和活性具有一定影响,热处理会导致不稳定的酚类化合物发生降解和含量发生变化(Kumari etal.,2015)。H组样品中有15种酚类代谢物的相对含量高于D组样品,这些物质包含黄酮类、异黄酮类、酚类、香豆素及其衍生物、肉桂酸及其衍生物化合物,在热烘干燥处理过程中其含量得到提升,倍性变化范围在0.7271~0.9624;其中,2'-羟基染料木素7-(6'-丙二酰糖苷)和鹰嘴豆芽素A 7-(6-丙二酰葡萄糖苷)2种异黄酮类化合物的相对含量上升趋势明显,表明热处理能破坏鸡屎藤黄酮类化合物与单糖或多糖部分之间的OH基团(O-糖苷),降解部分葡萄糖苷型类黄酮,使某些类黄酮成为游离态,从而导致某些异黄酮类化合物含量增加(Acosta-Estrada et al.,2014)。此外,适当的热处理结合态酚类化合物,会使结合态酚类提取率较低的化合物半溶出,导致其含量提高(Nayak et al.,2015)。6-羟基木犀草素、6-β-D-吡喃葡萄糖基-4',5-二羟基-3',7-二甲氧基黄酮、异紫花前胡内酯、秦皮乙素、b-D-Glucopyranosiduronic acid,2-naphthalenyl等酚类化合物差异相对小,表明相对于其他酚类代谢物,其热稳定性较高,可能是因为这些化合物中含有稳定的化学结构苯环和碳碳双键等或多糖上的氧离子与羟基形成氢键,在相互作用中形成葡聚糖凝胶,酚类化合物的稳定性得到提高(康子悦等,2021)。结合VIP和Fold Change分析可知,不同干燥方式处理的鸡屎藤中酚类化合物含量有不同程度的升高和下降,表明干燥方式对鸡屎藤酚类物质的含量具有显著影响。
2.6鸡屎藤酚类差异代谢物的KEGG代谢通路富集分析结果
对冷冻干燥和热烘干燥处理鸡屎藤中酚类差异代谢物的代谢通路进行富集分析,由表4和图6可知,2种干燥方式处理鸡屎藤中酚类物质变化的代谢通路匹配到7条,富集程度最高的前5条通路分别为类黄酮生物合成、苯丙烷生物合成、多种次生代谢产物的生物合成1、酪氨酸代谢及二苯乙烯类、二芳基庚烷类和姜辣素生物合成。其中,苯丙烷生物合成和类黄酮生物合成通路共有4种代谢物参与,分别为松柏醛、5-O-咖啡酰莽草酸、b-D-Glucopyrano-siduronic acid,2-naphthalenyl和紫杉叶醇,其中前2种化合物上调,后2种化合物下调。多种次生代谢产物的生物合成1通路包括异紫花前胡内酯和秦皮乙素,在此通路上2种化合物均上调;酪氨酸代谢通路包括苯酚和3,4-二羟基苯乙酸化合物,在此通路上有1种化合物上调,1种化合物下调;二苯乙烯类、二芳基庚烷类和姜辣素生物合成通路包括b-D-Glucopyranosiduronic acid,2-naphthalenyl和5-O-咖啡酰莽草酸2种代谢物,在此通路上有1种化合物上调,1种化合物下调。
2.7不同干燥方式处理鸡屎藤抗氧化活性与酚类差异代谢物的相关分析结果
由图7可知,除2'-羟基染料木素7-(6'-丙二酰糖苷)和鹰嘴豆芽素A 7-(6-丙二酰葡萄糖苷)外,其他酚类差异代谢物与ABTS阳离子自由基清除能力均无显著相关性。25种酚类差异代谢物与DPPH自由基清除能力和铜离子还原能力均表现出显著相关性,有10种酚类差异代谢物与这2个指标呈显著正相关,15种酚类差异代谢物与这2个指标呈显著负相关,其中2'-羟基染料木素7-(6'-丙二酰糖苷)、山柰酚-(2G-芹糖醇罗宾糖苷)、葡糖二苯乙烯、二氢辣椒素酯、7-乙氧基香豆素5种酚类差异代谢物与铜离子还原能力呈极显著负相关(P<0.001),其他酚类差异代谢物均显示不同程度的显著相关性,这些相关酚类差异代谢物所属类别较多,表明鸡屎藤发挥功能活性是多种代谢物协同作用的结果,并非单一作用。
3讨论
本研究采用冷冻干燥和热烘干燥对鸡屎藤进行处理,2种干燥方式处理后鸡屎藤的总黄酮和总酚含量均无显著变化,说明总黄酮和总酚对温度变化具有一定的稳定性。热加工过程虽然加速细胞破裂,但高温能使氧化酶和水解酶钝化,因此,高温加工并不会导致多酚降解,与Dewanto等(2002)研究发现热加工番茄的总酚和总黄酮含量均无显著变化的结果一致。
采用超高效液相色谱串联傅里叶变换质谱仪对热烘干燥和冷冻干燥处理的鸡屎藤中酚类物质进行检测,共检测到酚类化合物99种,主要包括黄酮类、香豆素及其衍生物、肉桂酸及其衍生物和酚类等。黄酮类化合物是鸡屎藤酚类化合物中数量最多的物质,说明黄酮类化合物是鸡屎藤的主要生物活性成分。槲皮素是一种天然存在的多羟基黄酮醇,是黄酮类化合物中活性强的抗氧化剂之一,多以糖基结合和糖苷的形式存在。2种干燥方式均检出槲皮素、槲皮素-3-半乳糖甙、槲皮素3-O-丙二酰葡萄糖苷、槲皮素3,4'-二葡萄糖苷、槲皮素3-O-alpha-L-鼠李糖基-(1->2)-alpha-L-阿拉伯吡喃糖苷、槲皮素3-(6'-丙二酰葡萄糖苷)和槲皮素3-葡萄糖基(1-3)鼠李糖基(1-6)半乳糖苷等;其中,槲皮素-3-半乳糖甙和槲皮素3-葡萄糖基(1-3)鼠李糖基(1-6)半乳糖苷的相对含量有显著变化,与黄建林和张展霞(2005)研究发现低温加热会使槲皮素糖苷发生部分水解,不同加热方式会影响槲皮素糖苷物质转化的结果一致。
以VIP>1、Fold Chang>1或Fold Chang<1且P<0.05为标准,从冷冻干燥和热烘干燥处理的鸡屎藤中筛选出25种极显著差异代谢物。与冷冻干燥方式相比,热烘干燥方式处理的鸡屎藤酚类物质中有10种代谢物出现下调,主要有黄酮类和香豆素及其衍生物;15种代谢物呈上调,主要为黄酮类、酚类和肉桂酸及其衍生物,说明温度对鸡屎藤中的酚类物质具有很大影响。热烘干燥后鸡屎藤表达上调的酚类代谢物明显多于表达下调的酚类代谢物,说明热烘干燥处理能转化鸡屎藤中的非挥发性代谢物,适当的热处理会促进鸡屎藤中代谢物的水解、取代、异构化以及其他热物理和化学反应;该结果与李云嵌等(2023)研究得出热泵干燥后三七叶表达上调的代谢物明显多于表达下调的代谢物结果一致。这些差异极显著的酚类化合物具有不同的功能效果。例如二氢辣椒素酯具有镇痛、治疗神经痛和关节炎、抗癌等生理作用(Hedrich etal.,2016)。异黄酮类2'-羟基染料木素7-(6'-丙二酰糖苷)和鹰嘴豆芽素A 7-(6-丙二酰葡萄糖苷),具有抗氧化和抗炎的作用(Zhang et al.,2010)。7-乙氧基香豆素、异紫花前胡内酯和秦皮乙素等香豆素类化合物,具有抗菌、抗肿瘤、抗病毒、抗炎、抗氧化等药理活性(Afek etal.,2010;聂安政等,2016;Wang et al.,2022b),其中秦皮乙素对黑色素瘤、肝癌、肺癌及胃癌等均有治疗作用(Riveiro etal.,2010;贾绍华等,2012)。3,4-二羟基苯乙酸具有较强的清除自由基活性和抗炎活性(Tang et al.,2016),同时具有防止肝损伤和调节胰岛素功能的作用(Carrasco-Pozo etal.,2015)。在这些酚类化合物的共同作用下,鸡屎藤不仅有独特的风味,还具有多种生理活性功能。由此可见,鸡屎藤具有开发成抗氧化、抗炎和抗病毒等一系列生理活性产品的潜能和前景。
有研究表明,植物多酚前体物质来源于糖代谢的中间产物,且经莽草酸途径、苯丙烷代谢途径和类黄酮代谢途径合成(林春草等,2022)。本研究通过对差异代谢物进行KEGG代谢通路富集分析,共获得7条代谢通路,苯丙烷生物合成和类黄酮生物合成代谢通路富集最丰富,而莽草酸途径未富集到,可能是因为核心物质莽草酸未鉴定分析;同时说明苯丙烷生物合成和类黄酮生物合成这2条代谢通路受温度影响较大,通路中紫杉叶醇和b-D-Glucopyranosiduronic acid,2-naphthalenyl相对含量上调,松柏醛和5-O-咖啡酰莽草酸相对含量下调,说明热处理易使松柏醛和5-O-咖啡酰莽草酸酚类化合物被氧化,易使酚类降解与蛋白质等大分子形成复合物,从而使含量下降,与Harakot等(2014)研究发现热处理使游离酚含量呈下降趋势的结果一致。Pourcel等(2005)研究表明,拟南芥种膜与菜豆的颜色由苯丙烷类化合物的氧化决定。因此推测苯丙烷生物合成代谢通路与色素形成有密切关系。
目前已有一些学者对鸡屎藤的酚类物质进行研究,但尚未见关于不同干燥方式对鸡屎藤酚类物质影响的报道。本研究采用非靶向代谢组学,分析不同干燥方式对鸡屎藤酚类代谢物的影响,为鸡屎藤的开发应用提供理论参考。下一步可针对干燥过程中代谢物的变化,运用细胞和动物模型对其生理活性进行更多的探讨。
4结论
不同干燥方式处理鸡屎藤未改变其酚类化合物种类,但对酚类化合物的含量和抗氧化活性有明显影响。鸡屎藤发挥功能活性不是单一代谢物作用,而是多种代谢物协同作用的结果。
参考文献(References):
高天元,雷雨恬,唐国琳,连艳,马羚,蒋桂华.2020.药用鸡矢藤药材化学成分的UPLC-Q-TOF-MS分析[J].中国实验方剂学杂志,26(17):134-141.[Gao T Y,Lei Y T,Tang G L,Lian Y,Ma L,Jiang G H.2020.UPLC-Q-TOF-MS analysis of chemical constituents of Paederiae herba for folk use[J].Chinese Journal of Experimental Traditional Medical Formulae,26(17):134-141.]doi:10.13422/j.cnki.syfjx.20201812.
胡海清,韩贺东,林燕,陈练红,王晓玲.2013.广西鸡屎藤的化学成分研究[J].中国中药杂志,38(16):2657-2660.[Hu H Q,Han H D,Lin Y,Chen L H,Wang X L.2013.Chemical constituents of Paederiapertomentosa[J].China Journal of Chinese Materia Medica,38(16):2657-2660.]doi:10.4268/cjcmm20131623.
黄国凯.2017.鸡矢藤的质量标准研究[D].广州:广州中医药大学.[Huang G K.2017.Study on the quality standard of Paederia scandens(Lour.)Merr.[D].Guangzhou:Guang-zhou University of Chinese Medicine.]
黄建林,张展霞.2005.微波加热对以槲皮素为甙元的黄酮甙的稳定性的影响[J].分析测试学报,24(2):15-18.[Huang J L,Zhang Z X.2005.Influence of microwave heating on the stability of quercetin glycosides[J].Journal of Instru-mental Analysis,24(2):15-18.]doi:10.3969/j.issn.1004-4957.2005.02.004.
贾绍华,刘冰洁,张道勇,季宇彬.2012.秦皮乙素诱导人胃癌SGC-7901细胞凋亡机制的研究[J].黑龙江医药,25(3):365-368.[Jia S H,Liu B J,Zhang D Y,Ji Y B.2012.Study on the mechanism of esculetin-induced apoptosis in SGC-7901 human gastric cancer cells[J].Heilongjiang Me-dicine Journal,25(3):365-368.]doi:10.14035/j.cnki.hljyy.2012.03.076.
康子悦,沈蒙,葛云飞,王娟,全志刚,肖金玲,王维浩,曹龙奎.2021.基于植物广泛靶向代谢组学技术探究小米粥中酚类化合物组成及其抗氧化性[J].食品科学,42(4):206-214.[Kang Z Y,Shen M,Ge Y F,Wang J,Quan Z G,Xiao J L,Wang W H,Cao L K.2021.Analysis of phenolic composition in millet porridge using widely-targeted metabolomics and evaluation of antioxidant activity[J].Food Science,42(4):206-214.]doi:10.7506/spkx 1002-6630-20190813-145.
李云嵌,何霞红,吴光顺,满金花,张雪春,王振兴.2023.干燥方式对三七叶主要活性成分、体外抗氧化、α-葡萄糖苷酶抑制活性、挥发性成分和代谢物的影响[J].食品科学,44(21):98-113.[Li Y Q,He X H,Wu G S,Man J H,Zhang X C,Wang Z X.2023.Effect of drying method on the major active components,in vitro antioxidant activity,α-glucosidase inhibitory activity,volatile components,and metabolites of Panax notoginseng leaves[J].Food Science,44(21):98-113.]doi:10.7506/spkx 1002-6630-20230217-160.
林春草,陈大伟,戴均贵.2022.黄酮类化合物合成生物学研究进展[J].药学学报,57(5):1322-1335.[Lin C C,Chen D W,Dai J G.2022.Advances of synthetic biology offla-vonoids[J].Acta Pharmaceutica Sinica,57(5):1322-1335.]doi:10.16438/j.0513-4870.2022-0008.
刘乐,王文可,卓静,唐娜,杨晴晴,李佳怡,刘飞红,王梦鸽,袁金红.2022.扫描电子显微镜叔丁醇冷冻干燥快速制样方法的探索[J].电子显微学报,41(1):98-104.[Liu L,Wang W K,Zhuo J,Tang N,Yang Q Q,Li J Y,Liu F H,Wang M G,Yuan J H.2022.Studies on scanning electron microscope method for rapid leaf sample preparation with tert-butanol[J].Journal of Chinese Electron Microscopy Society,41(1):98-104.]doi:10.3969/j.issn.1000-6281.2022.01.016.
刘若菡,刘红燕,张永清.2022.基于场发射扫描电镜的4种唇形科植物腺毛形态观察研究[J].电子显微学报,41(1):72-79.[Liu R H,Liu H Y,Zhang Y Q.2022.Obser-vation on the morphology of glandular trichomes in fourspecies of Lamiaceae based on field emission scanning electron microscope[J].Journal of Chinese Electron Mic-roscopy Society,41(1):72-79.]doi:10.3969/j.issn.1000-6281.2022.01.012.
聂安政,林志健,张冰.2016.秦皮化学成分和药理作用研究进展[J].中草药,47(18):3332-3341.[Nie A Z,Lin Z J,Zhang B.2016.Advance in studies on chemical consti-tuents of Fraxini Cortex and their pharmacological effects[J].Chinese Traditional and Herbal Drugs,47(18):3332-3341.]doi:10.7501/j.issn.0253-2670.2016.18.029.
秦丹丹,张生万,郭萌,郭彩霞,李美萍.2018.干燥方式对无花果酚类物质及其抗氧化活性的影响[J].食品科学,39(9):102-107.[Qin D D,Zhang S W,Guo M,Guo C X,Li M P.2018.Effect of drying methods on polyphenol compo-sition and antioxidant activities of figs(Ficus carica L.)[J].Food Science,39(9):102-107.]doi:10.7506/spkx1002-6630-201809016.
沈风娇,任倩倩,董琦,朱丽,张建芳,杨婧,张冉,梁红柱,赵建成,石硕.2017.一种适合野外使用的被子植物分子标本干燥方式[J].植物生态学报,41(7):787-794.[Shen F J,Ren Q Q,Dong Q,Zhu L,Zhang J F,Yang J,Zhang R,Liang H Z,Zhao J C,Shi S.2017.A new angiosperms molecular specimen treatment method for field use[J].Chi-nese Journal of Plant Ecology,41(7):787-794.]doi:10.17521/cjpe.2016.0322.
唐超玲,韩冬苗,孙万莹,黄兹宝,王瑞琪,张小波.2023.南药鸡矢藤的现代药学研究进展[J].海南医学院学报,29(20):1521-1525.[Tang C L,Han D M,Sun W Y,Huang Z B,Wang R Q,Zhang X B.2023.Research progress of Paederia scandens(Lour.)Merr in modern pharmacy[J].Journal of Hainan Medical University,29(20):1521-1525.]doi:10.13210/j.cnki.jhmu.20220914.002.
王露.2018.番石榴叶活性多酚组分快速鉴别及发酵释放与转化机制[D].广州:华南理工大学.[Wang L.2018.Quick identification and the mechanism in the release and trans-formation by fermentation of active phenolics components in guava leaves[D].Guangzhou:South China University of Technology.]
王晓宇.2008.葡萄酒抗氧化活性及其检测方法的研究[D].杨凌:西北农林科技大学.[Wang X Y.2008.Study onthe antioxidant activity and methods of detection in wine[D].Yangling:Northwest A&F University.]
王星星,王重娟,李仲昆.2021.鸡矢藤的化学成分及药理活性研究进展[J].世界中医药,16(5):826-830.[Wang X X,Wang Z J,Li Z K.2021.Research progress on chemical composition,pharmacological activity of Paederia scan-dens[J].World Chinese Medicine,16(5):826-830.]doi:10.3969/j.issn.1673-7202.2021.05.025.
吴雅璐.2019.不同干燥方法对花椒叶色泽、挥发性物质及抗氧化性的影响[D].太原:山西师范大学.[Wu Y L.2019.The effects of different drying methods on the color,vola-tile matter and antioxidant activity of Zanthoxylum bungea-num leaves[D].Taiyuan:Shanxi Normal University.]doi:10.27287/d.cnki.gsxsu.2019.000364.
邢颖,张月,徐怀德,周素梅.2020.不同干燥方法对生姜叶活性成分和抗氧化活性的影响[J].食品工业科技,41(18):75-80.[Xing Y,Zhang Y,Xu H D,Zhou S M.2020.Effects of different drying methods on active components and antioxidant activities of ginger leaves[J].Science and Technology of Food Industry,41(18):75-80.]doi:10.13386/j.issn 1002-0306.2020.18.012.
徐寸发,丁林云,周昊,汪佳蕾,杨晨晔,王宁,杨丹.2023.基于扫描电子显微镜制样的不同干燥方法对棉花和水稻幼苗叶片制备的差异分析[J].电子显微学报,42(3):358-364.[Xu C F,Ding L Y,Zhou H,Wang J L,Yang C Y,Wang N,Yang D.2023.Analysis of different drying methods for the preparation of Gossypium hirsutum and Oryza sativa seedling leaves based on scanning electron microscope[J].Journal of Chinese Electron Microscopy Society,42(3):358-364.]doi:10.3969/j.issn.1000-6281.2023.03.011.
徐秀苹,孟淑春,冯旻.2017.一种快速、简易的扫描电镜植物样品干燥新方法[J].中国细胞生物学学报,39(2):203-206.[Xu X P,Meng S C,Feng M.2017.New method on scanning electron microscopy(SEM)samples drying[J].Chinese Journal of Cell Biology,39(2):203-206.]doi:10.11844/cjcb.2017.02.0262.
张红建,刘帅光,马泽威,王青松,田燕,郑联合.2023.自然发酵对青金桔酚类物质及其抑制消化相关酶活性的影响[J/OL].食品工业科技.https://link.cnki.net/urlid/11.1759.TS.20231027.1134.008.[Zhang H J,Liu S G,Ma Z W,Wang Q S,Tian Y,Zheng L H.2023.Analysis of phenolicsubstances in Citrus microcarpa with nature fermentation and evaluation of its inhibitory activites on digestion-related enzymes[J/OL].Science and Technology of FoodIndustry.https://link.cnki.net/urlid/11.1759.TS.20231027.1134.008.]
张舒,王长远,冯玉超,盛亚男,富天昕,张艺玮,姜颖俊,于淼,张丽媛.2021.气相色谱—质谱联用代谢组学技术分析不同产地稻米代谢物[J].食品科学,42(8):206-213.[Zhang S,Wang C Y,Feng Y C,Sheng Y N,Fu T X,Zhang Y W,Jiang Y J,Yu M,Zhang L Y.2021.Analysis of metabolites in rice produced in different regions by GC-MS-based metabonomics[J].Food Science,42(8):206-213.]doi:10.7506/spkx 1002-6630-20200409-128.
Acosta-Estrada B A,Gutiérrez-Uribe J A,Serna-Saldivar S O.2014.Bound phenolics in foods,a review[J].Food Chemi-stry,152:46-55.doi:10.1016/j.foodchem.2013.11.093.
Afek U,Orenstein J,Aharoni N.2010.The involvement of marmesin and its interaction with GA3 and psoralens in parsley decay resistance[J].Canadian Journal of Plant Pa-thology,24(1):61-64.doi:10.1080/07060660109506972.
Akther F,Alim M A,Nasrin N A,Khan M,Gomes D N,Suhan M,Islam M,Begum R.2023.Effects of different drying methods on the proximate composition,antioxidant activi-ty,and phytochemical content of Hibiscus sabdariffa L.Calyx[J].Food Chemistry Advances,3:100553.doi:10.1016/j.focha.2023.100553.
Antal T,Chong C H,Law C L,Sikolya L.2014.Effects of freeze drying on retention of essential oils,changesinglan-dular trichomes of lemon balm leaves[J].International Food Research Journal,21(1):387-394.
Carrasco-Pozo C,Gotteland M,Castillo R L,Chen C.2015.3,4-dihydroxyphenylacetic acid,a microbiota-derived meta-bolite of quercetin,protects against pancreaticβ-cells dys-function induced by high cholesterol[J].Experimental Cell Research,334(2):270-282.doi:10.1016/j.yexcr.2015.03.021.
Dewanto V,Wu X Z,Adom K K,Liu R H.2002.Thermal pro-cessing enhances the nutritional value of tomatoes by increasing total antioxidant activity[J].Journal of Agricul-tural and Food Chemistry,50(10):3010-3014.doi:10.1021/jf0115589.
Duarte L,Gasaly N,Poblete-Aro C,Uribe D,Echeverria F,Gotteland M,Garcia-Diaz D F.2021.Polyphenols and their anti-obesity role mediated by the gut microbiota:A comprehensive review[J].Reviews in Endocrine&Meta-bolic Disorders,22(2):367-388.doi:10.1007/s 11154-020-09622-0.
Harakot B,Suriharn B,Tangwongchai R,Scott M P,Lertrat K.2014.Anthocyanin,phenolics and antioxidant activity changes in purple waxy corn as affected by traditional cooking[J].Food Chemistry,164:510-517.doi:10.1016/j.foodchem.2014.05.069.
Hedrich W D,Hassan H E,Wang H B.2016.Insights into CYP2B6-mediated drug-drug interactions[J].Acta Pharma-ceuticaSinica B,6(5):413-425.doi:10.1016/j.apsb.2016.07.016.
Jiang G T,Kang H Y,Yu Y Q.2017.Cross-platform metabolo-mics investigating the intracellular metabolic alterations of HaCaT cells exposed tophenanthrene[J].Journal of Chro-matography B,1060:15-21.doi:10.1016/j.jchromb.2017.05.023.
Kumari S,Krishnan V,Sachdev A.2015.Impact of soaking and germination durations on antioxidants and anti-nutrients of black and yellow soybean(Glycine max L.)varieties[J].Journal of Plant Biochemistry&Biotechno-logy,24:355-358.doi:10.1007/s 13562-014-0282-6.
Liu M,Wang L,Huang B J,Lu Q,,Liu R.2022a.3,4-dihy-droxyphenylacetic acid ameliorates gut barrier dysfunction via regulation of MAPK-MLCK pathway in type 2 diabe-tes mice[J].Life Sciences,305:120742.doi:10.1016/j.lfs.2022.120742.
Liu Y X,Zhe W,Zhang R F,Peng Z T,Wang Y X,Gao H Q,Guo Z Q,Xiao J.2022b.Ultrasonic-assisted extraction of polyphenolic compounds from Paederia scandens(Lour.)Merr.Using deep eutectic solvent:Optimization,identifica-tion,and comparison with traditional methods[J].Ultra-sonics Sonochemistry,86:106005.doi:10.1016/j.ultsonch.2022.106005.
Liu Y Y,Hu X,Li E H,Fang Y J,Xue H,Zhang J C,Jha R,Wang R M.2024.Bioaffinity ultrafiltration combined with UPLC-ESI-QTrap-MS/MS for screening of xanthine oxi-dase inhibitors from PaederiafoetidaL.leaves[J].Ara-bian Journal of Chemistry,17(4):105706.doi:10.1016/j.arabjc.2024.105706.
Nayak B,Liu R H,Tang J.2015.Effect of processing on pheno-lic antioxidants of fruits,vegetables,and grains—A review[J].Critical Reviews in Food Science and Nutrition,55(7):887-919.doi:10.1080/10408398.2011.654142.
Oliver S,Vittorio O,Cirillo G,Boyer C.2016.Enhancing thetherapeutic effects of polyphenols with macromolecules[J].Polymer Chemistry,7(8):1529-1544.doi:10.1039/c5py01912e.
Pourcel L,Routaboul J M,Kerhoas L,Caboche M,Lepiniec L,Debeaujon I.2005.TRANSPARENT TESTA10 encodes a laccase-like enzyme involved in oxidative polymerization of flavonoids in Arabidopsis seed coat[J].The Plant Cell,17(11):2966-2980.doi:10.1105/tpc.105.035154.
Rambaran T F,Nordstrm A.2021.Medical and pharmacoki-netic effects of nanopolyphenols:A systematic review of clinical trials[J].Food Frontiers,2(2):140-152.doi:10.1002/fft2.72.
Riveiro M E,De Kimpe N,Moglioni A,Vázquez R,Monczor F,Shayo C,Davio C.2010.Coumarins:Old compounds with novel promising therapeutic perspectives[J].Current Medicinal Chemistry,17(13):1325-1338.doi:10.2174/092986710790936284.
Tang Y,Nakashima S,Saiki S,Naomi A,Abe N,Kuwazuru S,Zhu B W,Ashida H,Murata Y,Nakamura Y.2016.3,4-dihydroxyphenylacetic acid is a predominant biologically-active catabolite of quercetin glycosides[J].Food Research International,89:716-723.doi:10.1016/j.foodres.2016.09.034.
Wang R M,Yao L L,Lin X,Hu X P,Wang L.2022a.Exploring the potential mechanism of Rhodomyrtus tomentosa(Ait.)Hassk fruit phenolic rich extract on ameliorating nonalco-holic fatty liver disease by integration of transcriptomics and metabolomics profiling[J].Food Research Internatio-nal,151:110824.doi:10.1016/j.foodres.2021.110824.
Wang S K,Chen TX,Wang W,Xu LL,Zhang Y Q,Jin Z,Liu Y B,TangY Z.2022b.Aesculetin exhibited anti-inflammatory activities through inhibiting NF-KB and MAPKs pathway in vitro and in vivo[J].Journal of Ethnopharmacology,296:115489.doi:10.1016/j.jep.2022.115489.
Wu Z J,Wang J H,Fang D M,Zhang G L.2013.Analysis of iri-doid glucosides from Paederia scandens using HPLC-ESI-MS/MS[J].Journal of Chromatography B,923-924:54-64.doi:10.1016/j.jchromb.2013.01.032.
Zhang J H,Du F P,Peng B,Lu R H,Gao H X,Zhou Z Q.2010.Structure,electronic properties,and radical scaven-ging mechanisms of daidzein,genistein,formononetin,and biochanin A:A density functional study[J].Journal of Mo-lecular Structure:Theochem,955(1-3):1-6.doi:10.1016/j.theochem.2010.04.036.
Zhang Z Y,Peng L,Chen Y R,Zhang K,Chen J H,Li Z,Li W L.2022.An integration of UPLC-Q-TOF/MS and chemo-metrics analysis for the holistic quality evaluation of diffe-rent geographical Paederia scandens[J].Rapid Communi-cations in Mass Spectrometry,36(13):e9317.doi:10.1002/rcm.9317.
(责任编辑罗丽)
