南亚热带生境下6种紫珠属植物的光响应模型拟合分析
2024-07-22邓丽张旻冯世秀


摘要 利用直角双曲线模型、非直角双曲线模型、直角双曲线修正模型和指数模型,对南亚热带生境下的6种紫珠属(Callicarpa)植物进行光合参数分析和光响应曲线拟合。通过对比4种模型计算光饱和点(LSP)、光补偿点(LCP)、最大净光合速率(Pnmax)、暗呼吸速率(Rd)和表观量子效率(AQE)以及决定系数(R2)、均方根误差(RMSE)、平均绝对误差(MAE)等参数,筛选出6种紫珠属植物最适光响应模型。结果表明:直角双曲线修正模型是6种紫珠属植物进行光合研究的最适用光响应模型,杜虹花对强光和弱光的利用能力均最高,朝鲜紫珠的光合潜能最大。综上,不同紫珠属植物对光的利用能力既表现出种间差异性,又表现出种间相似性,4种光响应模型在紫珠属植物的光合参数计算上存在较大差异,但在模型的拟合优度和拟合精度上存在一致性。
关键词 紫珠属植物;光响应模型;光合特性;南亚热带
中图分类号 S686 文献标识码 A 文章编号 0517-6611(2024)13-0101-06
doi:10.3969/j.issn.0517-6611.2024.13.025
开放科学(资源服务)标识码(OSID):
Fitting Analysis of Light Response Models for Six Species of Callicarpa L.Plants in South Asian Tropical Habitats
DENG Li,ZHANG Min,FENG Shi-xiu
(Xianhu Botanical Garden of Chinese Academy of Sciences/Shenzhen Key Laboratory of South Subtropical Plant Diversity,Shenzhen,Guangdong 518004)
Abstract In this study,four light models including rectangular hyperbola model,non-rectangular hyperbolic model,modified rectangular hyperbolic model and exponential model were jointly adopted to evaluate the photosynthetic parameters and light response fitting curves of six Callicarpa Linn.species in southern subtropical regions.By comparison of the light saturation point (LSP),light compensation point (LCP),maximum net photosynthetic rate (Pnmax),dark respiration rate (Rd) and apparent quantum efficiency (AQE),as well as the coefficient of determination (R2),root mean square error (RMSE) and mean absolute error (MAE) derived from the four models,the most suitable light response models for these six plants were obtained.The results demonstrated that the modified rectangular hyperbolic model performed the best fitting ability for the photosynthetic study of the six Callicarpa species,and the highest light utilization ability under both strong and weak light belonged to C.formosana,and the highest photosynthetic efficiency was found in C.japonica var.Luxurians.In summary,these six Callicarpa species showed both interspecific variability and interspecific similarity in light utilization capacity,and the four light response models differed greatly in the calculation of the photosynthetic potential,however,there was consistency in the model’s goodness-of-fit and fitting accuracy.
Key words Callicarpa Linn.;Light response model;Photosynthetic characteristics;Southern subtropics
基金项目 深圳市城市管理和综合执法局科研项目(202206)。
作者简介 邓丽(1992—),女,湖北麻城人,工程师,硕士,从事风景园林规划设计、城市规划与区域设计研究。*通信作者,教授级高级工程师,博士,从事资源植物与开发利用研究。
收稿日期 2023-07-14
植物的光合作用是植物进行有机物合成的重要生物过程,通过固定光能和CO2等非生物能量及原料,合成有机物并转换化学能,以供植物生长发育[1]。不同植物对光和CO2的响应和利用效率存在差异,研究植物的光响应曲线有助于确定植物的最大净光合速率(Pnmax)、光饱和点(LSP)、光补偿点(LCP)及暗呼吸速率(Rd)等光合参数[2]。借助光响应模型能快速有效拟合光响应曲线,目前常用的光响应模型有直角双曲线模型[3]、非直角双曲线模型[4]、指数模型[5]和直角双曲线修正模型[6]。以不同光响应模型,分别对同一组植物光合参数实测值进行分析计算,得出的拟合数值往往与实测值存在不同程度的差异,筛选出差异程度最小、拟合优度最高的光响应模型有助于更精确地了解植物的光合特性,为植物的应用配置形式提供参考。
紫珠属(Callicarpa L.)植物广泛分布于华中、华南等地区山间和林地。该属裸花紫珠(C.nudiflora)、广东紫珠(C.kwangtungensis)、杜虹花(C.formosana)等作为传统中药材应用历史悠久;老鸦糊(C.giraldii)、日本紫珠(C.japonica)等作为新型园林观果植物应用素材已逐步呈现在园林景观和大型花卉市场;裸花紫珠、大叶紫珠(C.macrophylla)等叶片硕大,叶面及叶背密被短毛或绒毛,能有效吸附扬尘等空气污染物,在城市园林应用中能发挥极佳的生态效益。当前针对紫珠属植物的研究集中于化学成分分析及提取物的药理药效研究[7-9],针对其生理特性的分析较少,且多在控制光照或水肥等非生物因素的条件下进行单个物种的光合特性研究。如,苏金等[10]通过控制荫蔽条件,研究了紫珠(C.bodinieri)叶片的叶绿素荧光和光合特征在不同光强下的变化;杨建伟等[11]通过控水法研究了田间持水量对小紫珠(C.dichotoma)光合生理的影响;李晨晨等[12]通过遮阴处理研究了裸花紫珠幼苗期的光合特性与药用成分含量之间的关系。上述研究均仅采用单一的光响应模型进行数据分析与处理,也并未对紫珠属种间植物的光合特性进行对比。
笔者选取在华南地区高温、湿热环境下生长发育良好的大叶紫珠、裸花紫珠、疏齿紫珠(C.remotiserrulata)、朝鲜紫珠(C.japonica var.luxurians)、老鸦糊以及杜虹花为研究对象,进行光合作用测量,并利用直角双曲线模型、非直角双曲线模型、直角双曲线修正模型和指数模型进行光合参数计算和光响应曲线拟合,对比4种模型的预测能力与准确程度,从而筛选出每种植物最适的光响应分析模型,进而为紫珠属植物的栽植繁育与园林应用提供参考依据。
1 材料与方法
1.1 试验地概况
试验地点为深圳市仙湖植物园(22°34′N,114°10′E),地处深圳第一高峰梧桐山西北山麓,毗邻深圳水库,受局部地区微气候影响,仙湖植物园内空气湿度较大,雨热充沛。深圳市属南亚热带季风气候,年平均气温高达23.0 ℃,年降雨量1 935.8 mm,年日照时长1 837.6 h。
1.2 试验材料
深圳市仙湖植物园内种植有紫珠属植物10余种,该试验选择植物园内自然生长发育较好的6个种进行光合作用测量。每个种选取6株2年生健壮植株,测取从顶端向下第3对成熟健康叶片的光合参数。
1.3 试验方法
选择2021年11月12—29日晴朗无风的天气开展试验,试验期间外部气温处于20~26 ℃。试验分为2个时段进行,分别为9:00—11:30和14:30—16:00。使用Li-6400XT便携式光合作用测量仪进行数据测量,开机后预热15~20 min,配置6400-02BLED红蓝光源后设置样品室CO2浓度为400 μmol/mol,设置光强梯度为2 000、1 500、1 200、1 000、800、700、600、500、400、200、100、50、20、10、0 μmol/(m2·s)。设定改变光强后每次等待时间不低于120 s,不高于200 s。每次测量开始前使用仪器自动测量程序进行CO2注入系统校准,并保存校准结果,确保进气系统稳定不漏气。每片叶片在测定完15个光强梯度数据后查看光响应曲线散点图,确保试验数据有效,然后依次进行后续叶片测定。
1.4 光响应模型
采用直角双曲线模型、非直角双曲线模型、指数模型和直角双曲线修正模型分别对6种紫珠属植物进行光响应曲线拟合和数据测算。
1.4.1 直角双曲线模型。直角双曲线模型[3]方程式为
Pn=α×PAR×Pnmaxα×PAR+Pnmax-Rd
式中:Pn为净光合速率,μmol/(m2·s);α为初始量子效率;PAR为光合有效辐射强度,μmol/(m2·s);Pnmax为最大净光合速率,μmol/(m2·s);Rd为暗呼吸速率,μmol/(m2·s)。
1.4.2 非直角双曲线模型。非直角双曲线模型的表达式为
Pn=
α×PAR+Pnmax-(α×PAR+Pnmax)2-4×k×α×PAR×Pnmax2×k-Rd
式中:k为反应非直角双曲线弯曲程度的曲角参数,取值范围为0≤k≤1;其他参数意义同上。
1.4.3 指数模型。指数模型的表达式为
Pn=Pnmax×(1-e-α×PARPnmax)-Rd
式中,e=2.718。
1.4.4 直角双曲线修正模型。直角双曲线修正模型的表达式为
Pn=α×PAR×(1-β×PAR)1+γ×PAR-Rd
式中,β,γ为修正系数。
1.5 模型评价
为验证各光响应模型的拟合优度,采用决定系数(R2)、均方根误差(RMSE)和平均绝对误差(MAE)进行对比,其中R2由光合计算模型计算得出,数值越高表明模型拟合程度越好,RMSE和MAE通过IBM SPSS Statistics计算得出,数值越低,则表明拟合程度越好[13]。
1.6 数据处理与分析
利用IBM SPSS Statistics软件对试验测取数据进行处理,计算每个种6组试验数据的Photo值的算术平均值,并以此记为实测值数据,根据实测值数据的走势预估各项光合参数,其中表观量子效率(AQE)和R2无法预估。将每个种6组试验数据分别代入4种光响应模型,计算同种6组数据在不同模型下的光合参数,并计算同种6组数据在相同模型下拟合值的算术平均值,以此算术平均值记为该种在该模型下的拟合值。直角双曲线模型和非直角双曲线模型下LSP需通过计算得出,计算公式为:LSP=(Pnmax+Rd)/AQE。指数模型下LSP采用光合速率为0.9Pnmax时所对应的光强数值[14]。
2 结果与分析
2.1 4种光响应模型对6种紫珠属植物的光响应曲线拟合效果评价
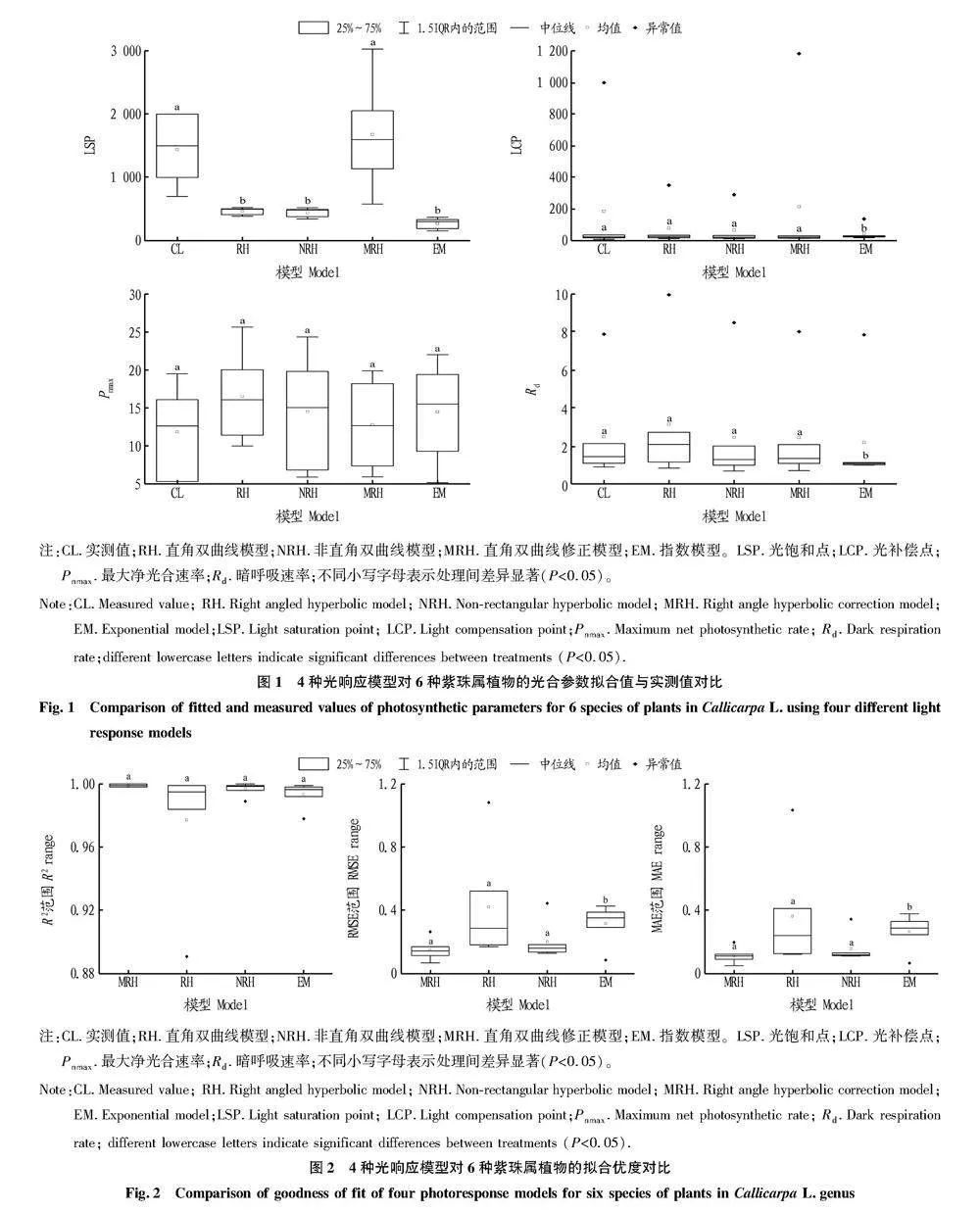
不同光响应模型对不同紫珠属植物光响应曲线拟合的R2均较高,除直角双曲线模型下疏齿紫珠R2低于0.900之外,其他模型下R2均高于0.970。不同光响应模型下同种植物的AQE、LSP、LCP、Pnmax和Rd模拟结果存在差异,且均与实测值之间存在不同程度的偏离(表1)。
在LSP模型模拟中,6种紫珠属植物MRH模型的模拟值与实测值CL差异最小,且在进行样本相关性检验时P<0.01,存在0.01水平显著性。而RH、NRH和EM模型的模拟值均显著小于CL(P<0.05)(图1)。在LCP模型模拟中,NRH和MRH模型的模拟值与CL之间存在显著正相关关
系,且P<0.01,模型拟合效果较好。RH和EM模型的模拟
值与CL之间不存在相关关系,P>0.05,表明这2种模型对LCP的模拟效果较差。
在Pnmax的模拟中,RH、NRH、MRH和EM模型的模拟值均于CL之间存在显著正相关,且相关系数均大于0.98,P<0.01,4种模型的拟合效果均较好。根据表1和图1可知,MRH和EM模型的Pnmax模拟值较其他2种模型的模拟值更接近于CL。
在Rd的模拟中,CL与NRH和MRH模型之间均存在显著正相关关系,均呈现出0.01水平的显著性。而CL与RH和EM模型之间相关系数较低,P>0.05,不存在相关关系。由图1可知,在Rd的模拟中MRH模型的预测值较NRH模型的预测值更接近CL,表明MRH模型的拟合效果更佳。
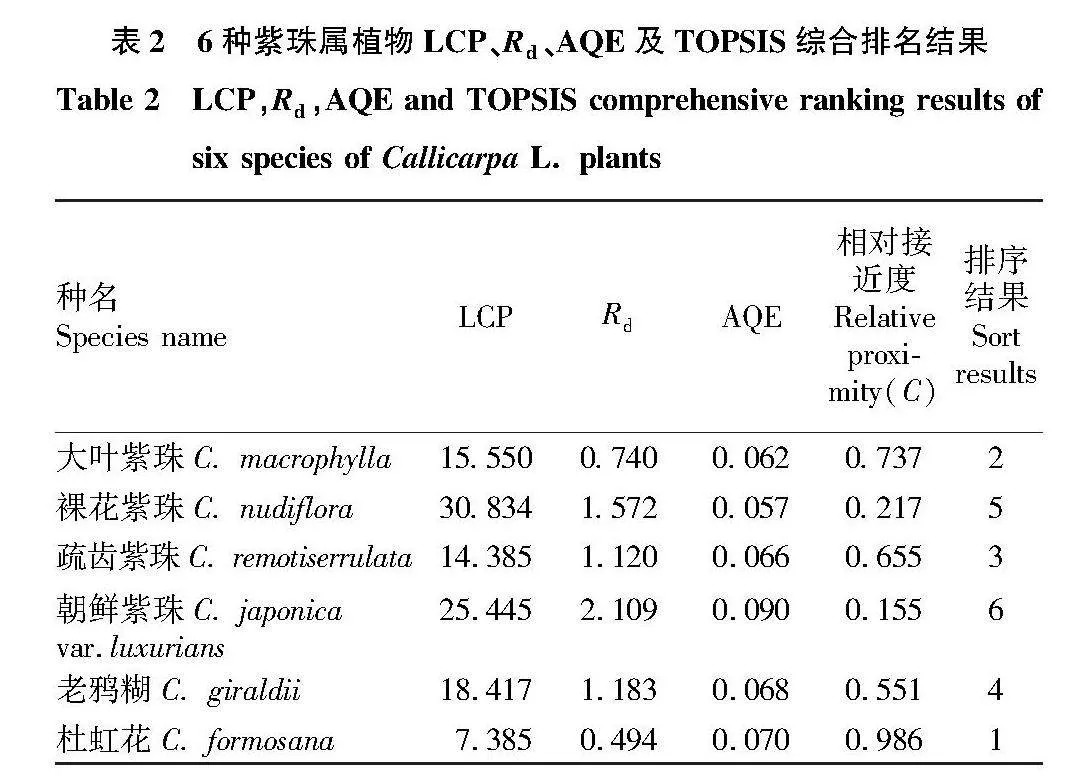
2.2 4种光响应模型对6种紫珠属植物光响应曲线的拟合优度评价
为筛选出适用于评价6种紫珠属植物的光响应模型,对4种模型的拟合优度进行评估,通过对比R2、RMSE和MAE的数值大小,以定量数据对模型拟合优度进行检验。由图2可见,通过对比R2,使用相关系数研究4种模型之间的相关关系强弱程度,可知RH、NRH、EM模型与MRH模型之间不存在显著相关关系(P>0.05)。通过对比4种模型的RMSE和MAE,发现MRH模型和NRH模型的RMSE和MAE小于其他2种模型。结合R2和RMSE以及MAE数值进行分析,MRH模型最适合6种紫珠属植物进行光响应曲线模拟,其次为NRH模型。
2.3 6种紫珠属植物的光合参数
2.3.1 光饱和点(LSP)。
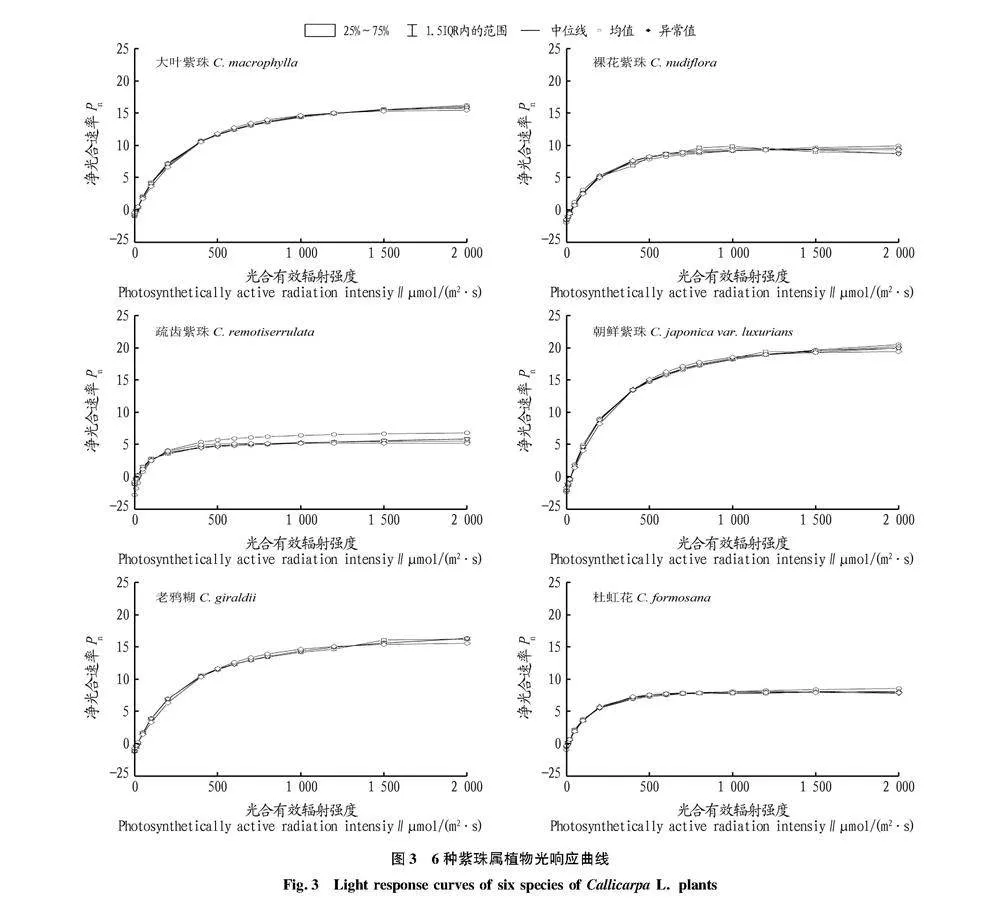
6种紫珠属植物的LSP为580.000~3 026.650 μmol/(m2·s),其中疏齿紫珠的LSP最低,为580.000 μmol/(m2·s),裸花紫珠和杜虹花的LSP接近1 200.000 μmol/(m2·s),大叶紫珠接近1 600.00 μmol/(m2·s),朝鲜紫珠和老鸦糊均在2 000.000 μmol/(m2·s)以上,表明疏齿紫珠为6种紫珠属植物中最不耐强光照的植物,朝鲜紫珠和老鸦糊在强光照环境中适应性较强(表1、图3)。
2.3.2 光补偿点(LCP)。
6种紫珠属植物的LCP为7.385~30.834 μmol/(m2·s),杜虹花LCP最低,疏齿紫珠、大叶紫珠和老鸦糊次之,朝鲜紫珠和裸花紫珠LCP较高。这表明杜虹花在光照强度较低的环境下仍能有效积累光合作用产物,而朝鲜紫珠和裸花紫珠则需要在光照强度大于25 μmol/(m2·s)时才能有效积累光合作用有机物(表1、图3)。
2.3.3 最大净光合速率(Pnmax)。
Pnmax[2]反映单位面积植物叶片利用光能进行光合作用的最大能力。疏
齿紫珠Pnmax最低,为5.961 μmol/(m2·s),杜虹花和裸花紫珠的Pnmax分别为8.060、9.494 μmol/(m2·s),大叶紫珠、朝鲜紫珠和老鸦糊的Pnmax分别15.861、19.884、18.194 μmol/(m2·s)(表1、图3)。
2.3.4 暗呼吸速率(Rd)。
Rd反映植物在弱光照条件下只进行呼吸作用而不进行光合作用时消耗的氧气和有机物的
能力,该项值越低,则表明植物在弱光照条件下适应能力越强[13]。
杜虹花和大叶紫珠的Rd均低于1.000 μmol/(m2·s),疏齿紫珠、老鸦糊和裸花紫珠的Rd为1.000~2.000 μmol/(m2·s),朝鲜紫珠Rd最高,为2.109 μmol/(m2·s)(表1、图3)。
2.3.5 表观量子效率(AQE)。AQE是植物在弱光照条件下进行光合作用能力的重要指征,是指在弱光照条件下[≤200 μmol/(m2·s)]以直线方程来拟合光响应曲线的初始斜率[15]。6种紫珠属植物的最适光响应模型MRH计算出的AQE数值在0.057~0.090,AQE为高优指标,朝鲜紫珠的AQE最高,杜虹花、老鸦糊和疏齿紫珠次之,裸花紫珠的AQE值最低(表1、图3)。
2.4 利用TOPSIS综合评价法评价6种紫珠属植物的弱光利用能力
为综合评价6种紫珠属植物对弱光的利用能力,利用TOPSIS评价法对指示植物利用弱光能力的3项参数LCP、Rd和AQE进行综合评价,其中LCP和Rd数值为低优数值,需先进行逆向化处理,确保3项评价参数同为正向指标。利用IBM SPSS Statistics进行数据逆向化处理后,通过TOPSIS法首先找出评价指标的正负理想解,接着计算出6种紫珠属植物分别与正负理想解的距离值,最终计算得出6种紫珠属植物与最优方案的相对接近程度(C值),该值越大,说明越接近最优方案。
由表2可知,通过TOPSIS评价法计算后,杜虹花利用弱光的能力最强,其次是大叶紫珠、疏齿紫珠和老鸦糊,裸花紫珠与朝鲜紫珠利用弱光进行光合作用的能力弱。
3 讨论与结论
3.1 讨论
3.1.1 不同光响应模型对南亚热带生境下6种紫珠属植物
光合参数的拟合效果。不同植物利用光的能力不同,适用的
最优光响应模型也存在差异,当前国内外学者进行光合作用研究时普遍使用直角双曲线模型、非直角双曲线模型、直角双曲线修正模型和指数模型。其中,直角双曲线修正模型由Ye等[6]提出,因其能直接计算出LSP数据,并且较其他模型而言叶子飘模型的预测值往往更接近实测值[16-18],还可以精确模拟出植物遭受光抑制时的光响应参数变化,故在国内研究中更多采用该模型进行光合参数计算与光响应曲线拟合。刘阳阳等[19]对新疆地区7个品种的山葡萄的最适光响应模型进行评价,发现直角双曲线修正模型能更有效地模拟光抑制情况下山葡萄的光合参数,且拟合值与实测值符合程度更高。蔡霞等[20]对结球甘蓝的光合模型进行了评价与筛选,发现叶子飘模型的拟合数值更接近于田间实测值,可以更好地分析结球甘蓝生长过程中的光需求特性。徐斌等[21]对广东含笑的光响应特征进行研究,发现相对于其他模型,叶子飘模型拟合的光响应曲线效果最佳。
该研究发现,最适宜用来分析紫珠属光合参数和光响应曲线的模型是直角双曲线修正模型,通过该模型计算得出的4项主要光合参数与实测值之间的误差较其他3种模型产生的误差更小,拟合的光响应曲线更接近实测值生成的曲线,模型拟合优度也是4种模型中最好的一种。非直角双曲线模型的拟合优度仅次于直角双曲线修正模型,但在LSP的模拟上预测值与实际值之间的误差大于直角双曲线模型的模拟误差。直角双曲线模型在LSP的模拟数值上较直角双曲线修正模型的模拟误差大,较非直角双曲线模型和指数模型的模拟误差小。在6种紫珠属植物的光合参数模拟中指数模型是4种模型中模拟准确度最低,模拟误差值最大的模型。
3.1.2 6种紫珠属植物的光合特性。
南亚热带地区具有强光照,全年日晒时间长,长夏短冬,夏季午间光照强度极高的特点。准确地掌握紫珠属植物的光合参数,有助于为该属植物的园林和生产应用评估科学合理的光照条件。
LSP反映了植物在强光照环境下的适应能力,该项指标关乎植物在何种光照强度下会发生光抑制的生理现象[22-23]。6种紫珠属植物中老鸦糊和朝鲜紫珠的LSP在实际测量中均未能有效测得,在光强梯度接近2 000 μmol/(m2·s)时Photo值仍处于不断上升的趋势,而通过MRH模型计算得出的LSP均超过2 000 μmol/(m2·s),其中老鸦糊的LSP超过3 000 mol/(m2·s)。观察试验地点自然种植的老鸦糊和朝鲜紫珠,发现在炎热的夏日2种植物仍处于缓慢生长状态,在全光照的条件下并未发生日晒灼伤现象,可知老鸦糊和朝鲜紫珠耐强光能力极强。大叶紫珠、裸花紫珠和杜虹花的耐强光能力也处于较高水平,LSP分别约为1 500、1 200 及1 100 μmol/(m2·s)。疏齿紫珠是6种紫珠属植物中最不耐强光照的植物,LSP约为600 μmol/(m2·s)。在实地观察中也发现,在夏季烈日直射下部分植株叶片出现黑色斑块,该种更适宜种植于林下遮阴地带。
Pnmax反映植物叶片在同时进行光合作用和呼吸作用的情况下积累物质的最大能力,该项值越高表明植物叶片的光合潜能越大[24]。与LSP一样,6种紫珠属植物的Pnmax表现出相似的品种差异特性,朝鲜紫珠和老鸦糊的Pnmax处于第一梯队水平,光合潜能最大,其次是大叶紫珠,裸花紫珠和杜虹花再次之,疏齿紫珠的Pnmax处于最低水平。
AQE、LCP和Rd反映植物利用弱光的能力[25]。其中,AQE为植物在LCP附近时忽略光响应曲线的非线性关系,计算得到的量子效率,即为AQE[15]。AQE的数值可能受有效辐射强度、统计的数据点个数[26]、植物种类[27-28]、土壤基质[29]等因素影响而存在差异,该数值越高表明植物的光能转化效率越高[30],对弱光的利用能力也越高[31]。LCP反映植物对光照的要求,该值越低表明植物对弱光的利用能力越强[32-33]。Rd反映植物在光强为0时的呼吸速率,该值越小表明植物在进行CO2交换时消耗的Pn越小[34],在弱光照条件下占优势。综合AQE、LCP和Rd 3项光合参数,发现杜虹花的LCP和Rd值为6种紫珠属植物中最低水平,AQE值仅次于朝鲜紫珠,这表明杜虹花单位面积的叶片耐阴能力强,呼吸作用消耗有机物速率低,光能利用效率高,在弱光照条件下生存能力强。疏齿紫珠、大叶紫珠和老鸦糊的LCP数值处于相似水平,AQE差异较小,大叶紫珠的Rd暗呼吸速率比其他2种植物略低,这3种植物对弱光的利用能力基本类似。朝鲜紫珠和裸花紫珠LCP值较高,Rd也处于最高水平,虽然朝鲜紫珠表现较高的AQE值,但通过TOPSIS法综合分析后这2种植物利用弱光的能力仍不及其他4种植物,这2种植物在园林和生产应用中应注意营造充足的光照条件,确保植物正常生长发育。
3.2 结论
综上所述,对南亚热带6种紫珠属植物进行光合作用研究时,最适用的光响应模型为直角双曲线修正模型,在模型的拟合优度、拟合效果和拟合精度上均为4种模型中最佳模型。对6种紫珠属植物的光能利用能力进行综合评价,6种紫珠属植物的光合潜能、耐阴性、耐强光能力存在物种间的差异性,朝鲜紫珠的光合潜能最大,杜虹花的耐阴性最强,老鸦糊的耐强光能力最佳。对6种紫珠属植物的光耐性与光合潜能进行综合排名,表现为老鸦糊>朝鲜紫珠>大叶紫珠>杜虹花>裸花紫珠>疏齿紫珠。
参考文献
[1]
叶子飘.光合作用对光和CO2响应模型的研究进展[J].植物生态学报,2010,34(6):727-740.
[2] 叶子飘,于强.光合作用光响应模型的比较[J].植物生态学报,2008,32(6):1356-1361.
[3] BALY E C C.The kinetics of photosynthesis[J].Proceedings of the royal society B:Biological sciences,1935,117(804):218-239.
[4] Thornley J H M.Mathematical models in plant physiology,A quantitative approach to problems in plant and crop physiology[M].London : Academic Press,1976: 86-110.
[5] BASSMAN J H,ZWIER J C.Gas exchange characteristics of Populus trichocarpa,Populus deltoides and Populus trichocarpa x P.deltoides clones[J].Tree physiology,1991,8(2):145-159.
[6] YE Z P,YU Q,KANG H J.Evaluation of photosynthetic electron flow using simultaneous measurements of gas exchange and chlorophyll fluorescence under photorespiratory conditions[J].Photosynthetica,2012,50(3):472-476.
[7] 占丽丽,叶先文,张敏,等.紫珠属植物化学成分及药理活性研究进展[J].江西中医药,2020,51(8):66-73.
[8] 黄梅,陈振夏,于福来,等.裸花紫珠主要化学成分的分布及其动态积累研究[J].中草药,2020,51(5):1308-1315.
[9] 孙宜春,黄春跃,李慧馨,等.基于苯乙醇苷类成分的紫珠属药材鉴别[J].中国医药工业杂志,2021,52(9):1230-1236.
[10] 苏金,方炎明,张强,等.遮阴对紫珠光合特性的影响[J].东北林业大学学报,2019,47(11):47-51.
[11] 杨建伟,赵丹,孙桂芳,等.干旱胁迫对小紫珠光合生理的影响[J].西北植物学报,2018,38(4):733-740.
[12] 李晨晨,周再知,周树平,等.遮荫对裸花紫珠苗期光合特性及总黄酮含量的影响[J].热带亚热带植物学报,2017,25(6):569-578.
[13] 李雪琴,卢艺苗,黄爱梅,等.亚热带10种蕨类植物光响应模型拟合及光合特性研究[J].生态学报,2022,42(8):3333-3344.
[14] 黄红英,窦新永,孙蓓育,等.两种不同生态型麻疯树夏季光合特性的比较[J].生态学报,2009,29(6):2861-2867.
[15] 叶子飘.光响应模型在超级杂交稻组合-Ⅱ优明86中的应用[J].生态学杂志,2007,26(8):1323-1326.
[16] 潘越,郭靖,韩政伟,等.不同配方叶面肥处理下苹果光响应模型选择及综合评价[J].西南农业学报,2021,34(4):806-813.
[17] 何昕孺,王琴平,秦垦,等.枸杞不同品系的光合特性及光响应模型拟合研究[J].中国农学通报,2020,36(28):57-62.
[18] 吴旭,唐亚坤,陈晨,等.黄土丘陵区沙棘、油松和刺槐光合生理特性及其环境适应性[J].生态学报,2019,39(21):8111-8125.
[19] 刘阳阳,潘越,王世伟,等.不同山葡萄品种光响应模型拟合及综合评价[J].中国农业科技导报,2022,24(2):104-114.
[20] 蔡霞,张德军,郭惊涛,等.结球甘蓝的光合模型筛选与光响应特征比较[J].种子,2021,40(11):117-121,136.
[21] 徐斌,朱报著,潘文,等.广东含笑的光响应特性及其最适模型研究[J].林业科学研究,2017,30(4):604-609.
[22] 呼和牧仁,周梅,翟洪波,等.影响树木光合作用因素的研究进展[J].内蒙古农业大学学报(自然科学版),2009,30(2):287-291.
[23] 姜海凤.三种落叶松光合生理生态学特性比较研究[D].哈尔滨:东北林业大学,2003.
[24] 许大全.光合作用效率[M].上海:上海科学技术出版社,2002.
[25] 曾小平,赵平,蔡锡安,等.25种南亚热带植物耐阴性的初步研究[J].北京林业大学学报,2006,28(4):88-95.
[26] 郎莹,张光灿,张征坤,等.不同土壤水分下山杏光合作用光响应过程及其模拟[J].生态学报,2011,31(16):4499-4508.
[27] 胡文海,胡雪华,曾建军,等.干旱胁迫对2个辣椒品种光合特性的影响[J].华中农业大学学报,2008,27(6):776-781.
[28] 张昆,万勇善,刘风珍,等.花生幼苗光合特性对弱光的响应[J].应用生态学报,2009,20(12):2989-2995.
[29] 陈建,张光灿,张淑勇,等军.辽东楤木光合和蒸腾作用对光照和土壤水分的响应过程[J].应用生态学报,2008,19(6):1185-1190.
[30] 董志新,韩清芳,贾志宽,等.不同苜蓿(Medicago sativa L.)品种光合速率对光和CO2浓度的响应特征[J].生态学报,2007,27(6):2272-2278.
[31] RICHARDSON A D,BERLYN G P.Spectral reflectance and photosynthetic properties of Betula papyrifera (Betulaceae) leaves along an elevational gradient on Mt.Mansfield,Vermont,USA[J].American journal of botany,2002,89(1):88-94.
[32] SENEVIRATHNA A M W K,STIRLING C M,RODRIGO V H L,et al.Growth,photosynthetic performance and shade adaptation of rubber (Hevea brasiliensis) grown in natural shade[J].Tree physiology,2003,23(10):705-712.
[33] 郭亮,杨国顺,白描,等.避雨栽培条件下不同葡萄品种光合特性的研究[J].湖南农业科学,2015(2):10-13.
[34] 娄玉穗,尚泓泉,吕中伟,等.基于光合特性的8个葡萄品种耐弱光、抗高温特点比较[J].果树学报,2021,38(9):1491-1502.
