外源Ca2+对干旱胁迫下4个牡丹品种抗性生理的调控效应
2024-07-03郁敏米银法
郁敏 米银法


doi:10.15889/j.issn.1002-1302.2024.10.024
摘要:以4个观赏性状优良的牡丹品种银红巧对、洛阳红、八千代春、胡红为试材,研究干旱胁迫下外源CaCl2对其叶内生理指标的调控机制以及不同品种间的响应差异,利用隶属函数值法评价Ca2+对4种牡丹抗旱性调控的总体效果。结果表明,喷施Ca2+后,4个牡丹品种的叶MDA、H2O2含量和 O-2·生成速率显著降低,可溶性蛋白、游离脯氨酸含量和SOD、CAT、POD活性显著提高。表明外施Ca2+能明显改善干旱胁迫下牡丹细胞的渗透势,维护良好的细胞膨压,有效清除ROS中H2O、O-2·对细胞膜的毒害作用,保持较好的细胞膜状态,提高植株的抗逆能力。胁迫期内,4种牡丹对喷施Ca2+的生理调控机制在品种间差异显著。与各自对照相比,在叶H2O2含量方面,银红巧对、胡红在轻度、中度干旱时显著降低,重度干旱时显著升高;洛阳红、八千代春在轻度干旱时显著降低,中度、重度干旱时显著升高。游离脯氨酸含量的最大增幅除八千代春出现在中度干旱外,其余3个品种均出现在重度干旱。外施CaCl2没有改变4种牡丹叶内CAT活性先升后降及POD活性逐渐升高的变化趋势。在叶内SOD活性方面,除轻度干旱时的八千代春与对照无显著差异外,其余均比各自对照显著提高,其中银红巧对、胡红即使在重度干旱时外施Ca2+仍能诱导SOD保持较高的活性。结果表明,Ca2+对银红巧对、胡红的调控作用效果总体优于洛阳红、八千代春。总体而言,Ca2+调控效果由强到弱顺序为胡红>银红巧对>洛阳红>八千代春。本研究结果可为观赏牡丹的田间栽培管理、抗旱品种的选配、砧木的选择与引种等提供实践依据和理论指导。
关键词:牡丹;外源钙;干旱胁迫;生理调节;评价
中图分类号:S685.110.1 文献标志码:A
文章编号:1002-1302(2024)10-0179-07
收稿日期:2023-06-25
基金项目:国家自然科学基金(编号:31872157);国家重点研发计划(编号:2018YFD1000800);河南省科技攻关项目(编号:232102110031)。
作者简介:郁 敏(1982—),女,河南郑州人,硕士,副教授,从事园林植物与观赏园艺方面教研工作。E-mail:jimiapple@126.com。
随着世界范围内碳排放以及温室效应的日益加剧,全球气温升高、气候紊乱、降水分布不均、极端天气频繁,导致干旱问题普遍频发,植物在生长过程中出现组织大量缺水、正常生理代谢紊乱、生物代谢酶失活、光合同化力下降等[1-2],严重时造成植物减产、绝产,甚至死亡。农业干旱问题已成为一项全球公认的普遍自然灾害,科学界也一直致力于寻求多种途径来缓解干旱胁迫造成的危害。许多研究表明,干旱逆境下植物叶面或根施耦联胞外信号与细胞内生理生化反应的重要第2信使物质即外源Ca2+,能够调控植物体内的抗氧化系统,显著提高植物抗氧化酶如过氧化氢酶(CAT)、过氧化物酶(POD)、超氧化物歧化酶(SOD)的活性和渗透调节物质的含量,降低MDA含量和H2O2、 O-2·的生成速率等,来维护植物细胞膜结构的稳定性,进一步缓解干旱胁迫对植物造成的伤害,增强植物对干旱逆境的抵御能力。外源Ca2+能提高甘草[3]、商洛黄芩[4]、白刺花幼苗[5]的抗氧化酶CAT、POD、SOD的活性和木豆种苗[6]的可溶性蛋白、脯氨酸含量;显著降低樟子松苗木[7]、棉花幼苗[8]的MDA含量和H2O2、 O-2·的生成速率,维护细胞膜结构的稳定性,显著增强这些植物的抗旱适应能力;并显著改善干旱胁迫下玉米植株的生长状况[9],提高植株的养分吸收能力。许多研究表明,遭受干旱胁迫时,可依据植物主要生理指标的变化,利用隶属函数值对植物的抗性进行总体评价[10];品种的隶属函数值越大,其抗旱能力越强[11-12]。黄莉娟等研究干旱胁迫下6个不同抗性的毛花雀稗生理生化指标的变化情况,利用隶属函数法综合评价不同品种毛花雀稗植株的耐旱性,发现美国种质具有很高的综合隶属函数值,抗旱性最强;贵州安龙(草坡)种质的综合隶属函数值次之,抗旱性良好;越南种质的综合隶属函数值最低,抗旱性最差;其余种质居中[13]。
在传统名花牡丹(Paeonia suffruticosa Andr.)根部土壤缺水时,有关外源Ca2+对不同牡丹品种生理指标的调节机制的研究尚不多见。本试验以不同抗旱能力的牡丹品种为试材,研究外源Ca2+对根部缺水条件下牡丹叶内生理指标的调控机制和品种间的响应差异,以期为观赏牡丹的田间栽培管理、抗旱品种选配、砧木选择与引种等提供实践依据和理论基础。
1 材料与方法
1.1 试验材料
试验于2020年7月至2021年6月在河南省林业科学研究院进行。以4个牡丹品种银红巧对、洛阳红、八千代春、胡红的3年生牡丹分株苗为试材,试材均来自国家牡丹基因库(洛阳)。
1.2 试验设计
2020年7月初选择长势良好、均匀一致的优良苗,以肥沃的大田土为基质,统一修整后栽种在内径30 cm、高28 cm的花盆内,1盆1株,每种牡丹栽种10株,正常肥水管理。 2021年6月3—13日,将盆栽分为不喷施CaCl2组(对照组)和喷施CaCl2组(处理组)2组,每组设置5盆。对照组喷施蒸馏水,处理组喷施浓度为 0.3 mmol/L 的CaCl2,早晚间隔8 h喷施,对牡丹叶片正反面喷洒2次/d,喷至叶面有水滴落为止,连续喷施10 d。这10 d内,材料同时进行干旱胁迫试验,干旱胁迫期间搭建防雨棚遮雨备用。6月3—13日内按李军等的方法[14]每天傍晚对盆栽进行称重和补水,10 d内使盆栽维持土壤的相对含水量为80%。6月13日后进行控水处理,让所有盆栽自然干旱。测得第3、6、9天时的相对含水量分别为50%~55%(轻度干旱)、35%~40%(中度干旱)、25%~30%(重度干旱),控水至第10天各处理大部分叶片明显萎蔫时,结束室外试验。
1.3 测定项目
分别在试验期内第3、6、9天的10:00进行取样,用于各项指标测定。取样时统一采摘各牡丹植株基部以上第3~5节位的叶片,采样后立即放入冰盒,带回实验室用于指标测定。脯氨酸、丙二醛(MDA)和可溶性蛋白含量按照李合生的方法[15]测定。H2O2、 O-2·含量以及SOD(超氧化物歧化酶)、POD (过氧化物酶)、CAT(过氧化氢酶)活性参考高俊凤的方法[16]测定。每个指标重复测定3次,用Excel 2003进行数据处理。
1.4 干旱胁迫下外源Ca2+对不同牡丹品种抗旱性调节综合效应评价
以测定的8个生理指标为基础,按文献[10-12]中的方法,求得各牡丹品种各生理指标隶属函数值的平均值,根据平均值,综合评价外源Ca2+对干旱胁迫下牡丹的生理调节作用,确定Ca2+对不同品种牡丹干旱胁迫伤害的缓解效果[11-13]。
2 结果与分析
2.1 干旱胁迫下外源Ca2+对牡丹叶MDA、 O-2·、H2O2含量变化的影响
丙二醛(MDA)是植物遭受逆境胁迫细胞膜脂发生过氧化时的产物,MDA含量常作为衡量细胞膜脂过氧化程度和鉴定植物逆境抗性的重要生理指标[17]。由图1-A可知,4个供试牡丹品种叶内MDA含量随土壤干旱程度的增强而逐渐升高。叶片喷施CaCl2可不同程度地降低各牡丹品种的叶内MDA含量。土壤轻度干旱时,处理组胡红、银红巧对分别比对照组显著降低30.5%、19.9%(P<0.05);其余2个品种(洛阳红、八千代春)的对照组和处理组间无显著差异。土壤中度干旱时,处理组4个供试品种叶内MDA含量均比对照组显著降低,其中银红巧对、洛阳红、八千代春、胡红分别比对照组降低19.6%、20.5%、21.6%、25.9%(P<0.05)。土壤重度干旱时,处理组银红巧对、胡红的MDA含量均显著降低,比各自对照组分别降低13.5%、28.1%(P<0.05);洛阳红、八千代春则无显著差异。结果表明,外施CaCl2可明显减轻牡丹叶片细胞膜脂的过氧化程度,缓解干旱胁迫对牡丹植株的伤害。但CaCl2缓解伤害的效果因不同牡丹品种而异,外施CaCl2在3种干旱胁迫程度下对胡红、银红巧对均能显著改善,而对洛阳红、八千代春仅在中度干旱胁迫时效果显著。
植物体遭受逆境胁迫时会产生大量高能态的活性氧自由基,这些自由基彼此间发生关联反应,生成更多活性氧自由基产物,进而加剧逆境的伤害程度。因此,逆境条件下植物体内 O-2·、H2O2的含量高低是衡量活性氧自由基强弱的2个重要生理指标。由图1-B可以看出,4个牡丹品种植株遭受干旱胁迫时, O-2·生成速率随干旱的严重程度而逐渐
升高,均在严重干旱时达到最高。外施CaCl2后,4个牡丹品种在3种不同程度的干旱胁迫下,叶内 O-2·的生成速率均比对照显著降低。3种不同程度干旱胁迫条件下,银红巧对、洛阳红、八千代春、胡红的叶内 O-2·下降幅度范围分别为23.1%~33.3%、17.0%~31.9%、16.7%~30.6%、30.7%~43.1%。外施CaCl2能让胡红 O-2·在整个干旱胁迫过程中的生成速率维持在较低水平,且其生成速率降幅最大,和其余3个品种相比差异显著。总体而言,胡红清除 O-2·的生理应对机制优于其余3个品种。
由图1-C可知,随着干旱程度的加剧,未外施CaCl2的对照组中,洛阳红、八千代春叶内的H2O2含量在前期降低后期升高,银红巧对、胡红叶内H2O2含量则表现为前期不显著变化,但后期显著升高(P<0.05)。外施CaCl2后,轻度、中度干旱胁迫下,银红巧对、胡红的H2O2含量比对照显著降低,重度干旱胁迫下显著升高;轻度干旱胁迫下,洛阳红、八千代春的H2O2含量比对照显著降低,中度、重度干旱胁迫下则显著升高(P<0.05)。4个牡丹品种叶内H2O2的含量表现出显著的品种间差异。轻度干旱胁迫下,外施CaCl2,银红巧对、洛阳红、八千代春、胡红的叶内H2O2含量分别比对照降低43.1%、52.1%、50.7%、44.9%;中度干旱胁迫下,银红巧对、胡红的叶内H2O2含量分别比对照降低18.3%、30.7%。可见,外施CaCl2能显著降低H2O2对4个牡丹品种在轻度干旱胁迫下的伤害以及银红巧对、胡红在中度干旱胁迫下的伤害。总体而言,银红巧对、胡红的叶内H2O2对外施Ca2+的调控机制响应更为迅速积极。
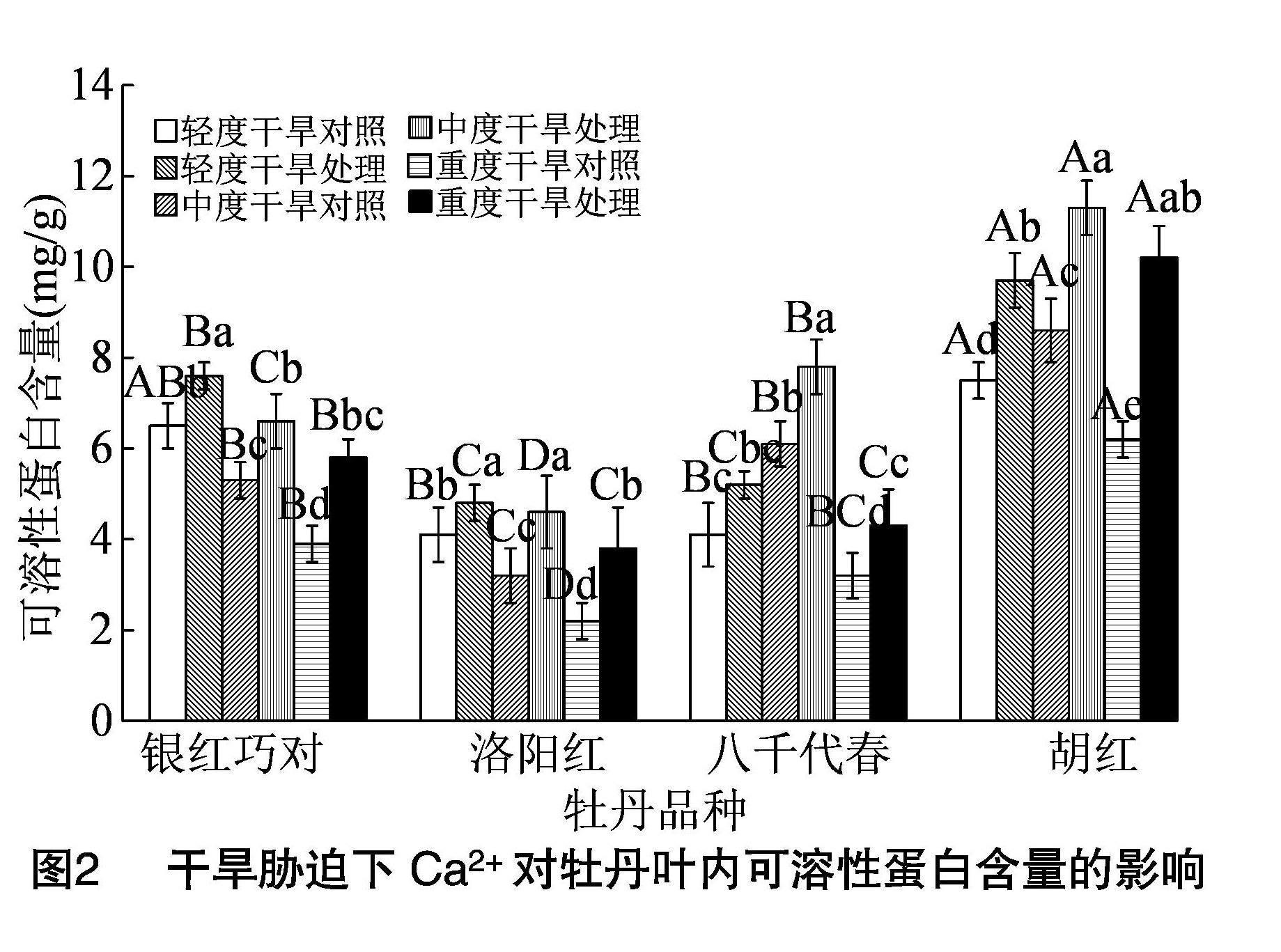
2.2 干旱胁迫下外源Ca2+对牡丹叶可溶性蛋白含量变化的影响
植物遭遇逆境胁迫时,细胞内往往诱发相应的生理调节机制,靠若干渗透调节类物质改变细胞渗透势,减轻胁迫伤害。其中,可溶性糖、游离脯氨酸、可溶性蛋白等渗透调节类物质常作为植物逆境生理研究的重要参考指标,也可作为品种抗性鉴定的主要依据。由图2可知,对照组遭受干旱胁迫时,银红巧对、洛阳红的叶内可溶性蛋白含量随干旱胁迫的加剧而逐渐降低;八千代春、胡红则先升高后降低,品种间差异显著。与各自对照相比,外施CaCl2后4个牡丹品种叶内的可溶性蛋白含量均显著升高,除八千代春在中度干旱胁迫下增幅最大外,其余3个品种均在重度干旱胁迫下增幅最大且最大增幅时银红巧对、洛阳红、八千代春、胡红比各自对照分别增加48.7%、72.7%、34.4%、64.5%,与不施加CaCl2的对照差异显著(P<0.05)。综上,外施CaCl2可显著诱发4个牡丹品种体内可溶性蛋白含量提高,进一步减轻干旱胁迫对细胞造成的伤害。
2.3 干旱胁迫下外源Ca2+对牡丹叶游离脯氨酸含量变化的影响
一般来讲,在遭受干旱、温度异常、高盐等逆境胁迫时,植物细胞质中将会积累大量游离脯氨酸,发挥渗透调节功能。此外,脯氨酸作为植物蛋白的重要组分,在稳定植物大分子结构、降低细胞酸性、缓解氨毒、调控细胞内生物氧化还原反应等方面起着至关重要的作用。因此,逆境胁迫下植物细胞脯氨酸的含量是植物抗逆能力及品种鉴定的重要参考,普遍认为脯氨酸含量高或增幅大的植物品种,其抗性强[6]。由图3可知,随着干旱胁迫程度的加剧,未外施CaCl2的4个牡丹品种对照组叶内脯氨酸含量在前期轻度、中度干旱阶段呈显著升高趋势,后期重度干旱阶段又呈显著降低趋势。这表明牡丹植株在干旱胁迫程度较轻阶段,可依赖脯氨酸的大量积累增强其抵御逆境的能力;但随着干旱胁迫程度的加重及胁迫时间的延长,后期游离脯氨酸含量又明显降低,这与干旱胁迫条件下绣球的研究结论[18]一致。可能是由于重度干旱胁迫对4个牡丹品种已造成较为严重且难以逆转的伤害。由本试验结果可知,外施CaCl2后4个牡丹品种的叶内游离脯氨酸含量均比各自对照显著增加。游离脯氨酸含量的最大增幅,除八千代春出现在中度干旱、洛阳红出现在重度干旱胁迫下之外,其余2个品种(银红巧对、胡红)均出现在轻度干旱胁迫下。可见干旱胁迫过程中,脯氨酸含量的增加幅度表现出显著的品种差异。与各自对照相比,整个干旱胁迫过程中,银红巧对、洛阳红、八千代春、胡红脯氨酸含量的增幅范围分别为24.4%~70.0%、40.6%~43.1%、31.0%~45.2%、58.1%~121.5%。因此在遭遇干旱胁迫时,可通过外施CaCl2的方法来改善牡丹细胞的渗透势,维持较好的细胞膨压,确保干旱逆境下的细胞膜维持优良状态,增强植株的抗逆能力。
2.4 干旱胁迫下外施Ca2+对牡丹叶SOD、CAT、POD活性变化的影响
逆境胁迫下植物体内活性氧ROS(如H2O2、 O-2·)迅速生成,超过承受阈值时,ROS生成与清除平衡机制将被打破,严重失衡时细胞膜结构受损,生理代谢紊乱,植物受到不可逆转的伤害。在自然进化过程中,植物体内形成了有效的抗氧化防御体系,其中以SOD、POD、CAT等为代表的抗氧化酶在清除活性氧方面起着至关重要的作用[19]。因此,这些酶的活性变化也是植物抗逆性研究及品种抗性
评价的重要依据。
由图4-A可知,遭遇干旱胁迫时,未外施CaCl2的对照组在轻度、中度干旱胁迫下,4个牡丹品种叶内的SOD活性均显著提高。洛阳红、八千代春的叶SOD活性随干旱胁迫程度的加剧呈先升后降的趋势。但银红巧对、胡红在轻度、中度干旱时升高,重度干旱时呈平稳趋势,即中度、重度干旱时无显著差异。这说明银红巧对、胡红在干旱胁迫后期仍能保持较高的SOD活性,而SOD作为清除活性氧系统的第1道防线,是 O-2·最重要的抗氧化酶清除剂,它的功能在于能把 O-2·歧化成毒性更低的H2O2,从而保护干旱胁迫下的牡丹植株。可见,银红巧对、胡红清除 O-2·的能力强于其余2个牡丹品种。外施CaCl2后,除轻度干旱胁迫下八千代春的叶内SOD活性与对照无显著差异外,其余3个牡丹品种的SOD活性均比各自对照显著提高;其中银红巧对、胡红即使在重度干旱胁迫下,外施Ca2+仍能诱导其叶内SOD保持较高的活性。这说明外施Ca2+可显著提高4个牡丹品种的叶内SOD活性,有效清除 O-2·产生的毒害作用;相对而言,Ca2+对银红巧对、胡红品种SOD活性的调控效果更显著。与各自对照相比,在整个干旱胁迫阶段外施Ca2+,银红巧对、洛阳红、八千代春、胡红的叶内SOD活性的增幅分别为21.3%~29.4%、22.7%~32.3%、15.6%~21.4%、32.7%~37.8%。
在几种有效的抗氧化酶中,CAT、POD重要的生理功能是协同清除叶绿体内SOD歧化 O-2·后的ROS产物H2O2,把H2O2催化成无毒产物H2O、O2,从而降低因 O-2·、H2O2累积对细胞造成的功能性损伤[15]。在本试验中,未喷施CaCl2的4个牡丹品种的叶内CAT活性随干旱胁迫程度的增加呈先升后降的变化趋势(图4-B),POD活性呈逐渐升高的变化趋势(图4-C)。说明干旱胁迫下,4个牡丹品种的CAT、POD在协同清除H2O2方面发挥了重要作用;但干旱胁迫程度的加据打破了牡丹植株内部活性氧的产生与清除平衡,造成 O-2·、H2O2过量积累,超出了及时清除的能力,因而后期又表现出含量升高的趋势,这与图1-B、图1-C相符。图4-B、图4-C表明,外施CaCl2并没改变整个胁迫期内4种牡丹CAT、POD活性的变化趋势,总体上CAT与未外施Ca2+的对照组(先升后降)的变化趋势一致,POD与与未外施Ca2+的对照组(逐渐升高)的变化趋势一致。但进一步分析表明,无论是CAT还是POD,外施Ca2+均能显著诱发和提高它们的活性,这与干旱胁迫时绣球POD的活性变化结果[18]一致。整个胁迫期外施Ca2+,与各自对照相比,银红巧对、洛阳红、八千代春、胡红的CAT活性增幅分别为11.7%~28.6%、21.5%~50.1%、20.4%~38.1%、22.4%~31.6%,POD活性增幅分别为27.3%~54.9%、4.4%~50.0%、26.3%~47.4%、22.4%~40.3%。可见,外施Ca2+在提高干旱胁迫下牡丹叶内CAT、POD活性,清除H2O2和调节改善牡丹抗旱性能方面作用显著。
2.5 干旱胁迫下外施Ca2+对4个牡丹品种抗性影响的综合效应评价
外源Ca2+对4个牡丹品种受干旱胁迫伤害的缓解效果,是内部调节机制下的诸多生理指标协同互作的综合反映,所以仅用少数生理指标进行评价,会造成较大的偏差或很难决定。因此,本研究选取与抗旱性密切相关的8个重要生理指标,运用隶属函数法对其作用效果进行总体评价。评价时为了兼顾公平,在3种(轻度、中度、重度)干旱胁迫条件下对各品种的隶属函数值的重要程度视为等同,均纳入考评,最终汇总求出总值(表1、表2),并按总分值由大到小进行综合效果强弱排序。经总体评价,Ca2+调控效应缓解效果由强到弱的顺序为胡红>银红巧对>洛阳红>八千代春(表3)。
3 讨论与结论
植物在生长过程中遭遇逆境胁迫严重时,活性氧产生与清除平衡机制失衡,抗氧化酶失活,高能态活性氧ROS大量积累,大量活性氧自由基之间彼此关联、相互促成,发生危害更重的Haber Weiss生化反应,又会产生更多的活性氧自由基产物,致使植物细胞生物膜发生更为严重的过氧化或脱脂化,造成细胞膜磷脂降解,细胞膜透性增大并生成大量产物MDA,因而 O-2·、H2O2、MDA的含量共同反映逆境下植物细胞膜的损伤程度,是衡量植物逆境损伤程度的3个重要指标,同时也是鉴定植物逆境抗性的重要依据[17]。本试验结果表明,4个牡丹品种在干旱胁迫下未喷施CaCl2时,MDA含量随胁迫程度加深逐渐升高, 但银红巧对、胡红的叶MDA含量和相对增幅在4个品种中均表现较低,说明3种不同干旱程度胁迫下,银红巧对、胡红的细胞膜损伤程度较轻,具有相对较强的抗旱性。叶片喷施CaCl2后,4个牡丹品种的叶MDA含量均不同程度地降低,但其对Ca2+的响应程度明显不同;在土壤轻度、重度干旱时,胡红、银红巧对比对照显著降低,而洛阳红、八千代春则无显著变化;可见土壤轻度、重度干旱时胡红、银红巧对的叶MDA对Ca2+的调节作用反应较为迅速,而洛阳红、八千代春则相对迟缓。中度干旱胁迫下,4个牡丹品种的叶内MDA含量均比对照显著降低,表明中度干旱时4种牡丹的叶内MDA对Ca2+的调节响应都很敏感。因而在牡丹遭受干旱时,外施CaCl2可明显减轻胁迫对牡丹叶片细胞膜脂的过氧化程度,降低对牡丹植株的伤害。可能是因为外施Ca2+能够显著提高牡丹叶肉细胞内的游离Ca2+浓度,进而促进较多的Ca2+参与细胞壁的构成,同时Ca2+也是植物细胞膜中磷酸和蛋白质羧基间的联结纽带,较高浓度的Ca2+可通过提高SOD活性、降低膜脂化物MDA含量等途径,来维护牡丹叶肉细胞膜的完整性,保护其细胞膜结构的完整性,这与尹大川等的研究结论[7]一致。但4种牡丹的叶内MDA含量对Ca2+的调控敏感程度各异。外施CaCl2时,胡红、银红巧对的MDA含量在3种胁迫程度下均显著改善;而洛阳红、八千代春仅在中度胁迫时效果显著。
就 O-2·而言,随干旱程度的加深,4种牡丹叶的 O-2·生成速率呈逐渐升高趋势。外施CaCl2后,4种牡丹叶 内的O-2·生成速率在3种干旱胁迫条件下均比对照显著降低,但 O-2·对Ca2+的响应机制在牡丹品种间存在差异。与银红巧对、洛阳红、八千代春相比,外施CaCl2后胡红的 O-2·生成速率在整个胁迫过程中均能维持较低的状态和较大的降幅(降幅为30.7%~43.1%),可见遭遇干旱胁迫时在外施Ca2+的诱导下,胡红具备更好的 O-2·清除应对机制。4种试材在根部缺水时,未喷施CaCl2牡丹的叶内H2O2含量随干旱胁迫程度的加剧表现不同,洛阳红、八千代春在前期降低后期升高,而银红巧对、胡红则前期变化趋势差异不显著,但后期显著升高。这表明4个牡丹品种在干旱胁迫下都受到了不同程度的生理伤害,尤其是重度干旱胁迫对牡丹植株伤害过于严重,超过胁迫阈值,打破了牡丹植株内H2O2产生与清除系统的平衡[11],因而胁迫后期4个牡丹品种的H2O2含量又呈显著升高的趋势。叶片外施CaCl2后,4个牡丹品种叶内H2O2含量的变化在品种间也存在显著差异。其中,银红巧对、胡红的H2O2含量在轻度、中度干旱时比对照显著降低,重度干旱时显著升高;洛阳红、八千代春的H2O2含量在轻度干旱时比对照显著降低,在中度、重度干旱时则显著升高。可见,外施Ca2+均降低了4种牡丹在轻度干旱时的H2O2含量,并降低银红巧对、胡红在中度干旱时的H2O2含量。说明银红巧对、胡红对外施Ca2+调控叶内H2O2的响应机制相对更为积极。
许多研究认为,逆境胁迫下,当有外源Ca2+或Ca2+载体存在时,植物细胞能迅速产生 O-2·、H2O2等活性氧分子,进而启动机体内其他信号,诱发一系列保护性的生理反应[20],从而减轻盐胁迫、干旱胁迫甚至病原菌侵害等对植物体的伤害。因此,可通过外施CaCl2的手段,来提高植物细胞内游离Ca2+浓度并维持一定的高浓度状态;高浓度的Ca2+一方面可促进与CaM结合,另一方面又有利于SOD、POD、CAT、APX等一系列保护酶类的快速表达和可溶性蛋白、可溶性糖、脯氨酸等渗透调节物质生理生化反应的迅速启动,从而大大降低细胞的逆境伤害,形成适应机制[21]。本试验中4种牡丹在遭遇干旱胁迫时,叶肉细胞内迅速产生 O-2·、H2O2等活性氧分子,诱导可溶性蛋白、脯氨酸等渗透调节物质生理生化反应得以迅速启动,SOD、POD、CAT等一系列保护酶类得以快速表达。与对照相比,外施Ca2+显著提高4个供试牡丹品种叶内游离脯氨酸、可溶性蛋白含量,其中游离脯氨酸含量均在中度干旱时最高。胁迫过程中,可溶性渗透调节物质的增加幅度在品种间存在差异;尤其是胡红的脯氨酸含量增加幅度在轻度干旱时就达到了121.5%。可见,遭遇干旱胁迫时可以通过外施Ca2+维护干旱胁迫下牡丹良好的细胞渗透势和细胞膨压,使牡丹叶肉细胞膜维持良好的状态,增强植株抗性。
植物遭遇逆境胁迫时,SOD作为抗氧化酶,是清除活性氧系统的第1道防线,也是 O-2·最重要的清除剂,它的功能在于能把 O-2·歧化成毒性更低的H2O2,从而保护植株免受逆境伤害[3]。本研究结果表明,遭遇干旱胁迫时外施CaCl2,除轻度干旱时的八千代春叶内SOD活性与对照无显著差异外,其余3个牡丹品种的SOD活性均比各自对照显著提高,尤其是银红巧对、胡红,即便在重度干旱时,Ca2+仍能诱导其SOD保持较高的活性状态。说明外施CaCl2可提高4种牡丹叶内SOD活性,有效清除 O-2·产生的毒害作用,且Ca2+对银红巧对、胡红SOD活性的调控作用效果更好。
CAT、POD是协同清除由SOD歧化 O-2·后的高能态活性氧产物H2O2的2种重要生物活性酶,催化反应为2H2O2→2H2O+O2,经此反应后,可将H2O2转化成无毒无害的产物H2O、O2,从而降低细胞损伤[15]。本试验中随干旱胁迫程度的增加,未外施CaCl2的4个牡丹品种的CAT活性呈先升后降、POD活性呈逐渐升高的变化趋势,说明CAT、POD在协同清除H2O2方面发挥了重要作用。只不过随着干旱胁迫程度的加重,内部活性氧自由基的产生与清除动态平衡被破坏,产生的过量 O-2·、H2O2超出了其及时清除的能力,又表现出 O-2·、H2O2含量升高的现象。从整个干旱胁迫过程来看,外施CaCl2基本上不会改变4种牡丹叶内CAT、POD活性的变化趋势,即总体上CAT活性与对照组先升后降的变化趋势一致,POD活性与对照组逐渐升高的变化趋势一致,但外施Ca2+均显著诱导了CAT、POD活性的提高。可见Ca2+在提高干旱胁迫下牡丹叶内SOD、CAT、POD活性和清除 O-2·、H2O2并提高牡丹植株的抗旱性能等方面效果明显。经多性状指标隶属函数值法总体评价后,Ca2+对4种牡丹受干旱胁迫伤害的缓解效果由强到弱的顺序为胡红>银红巧对>洛阳红>八千代春。本研究可以为选择牡丹砧木以及抗性育种提供理论参考。
本研究证明,外施Ca2+可促使4种供试牡丹植株通过降低 O-2·、H2O2含量,提高可溶性蛋白、脯氨酸等渗透调节物质含量以及SOD、POD、CAT活性等方式,来减轻干旱胁迫对4个供试牡丹植株的伤害。但也有研究者认为,Ca2+虽对细胞膜有保护作用,但受其浓度高低的影响,如果细胞质、细胞核中Ca2+浓度较高,存在时间过长不能及时泵回到钙库或者胞外,可能会引起植物“钙中毒”,同时过高浓度的Ca2+也会与磷酸反应生成沉淀,干扰以磷酸为基础的能量代谢[22]。总之,在干旱胁迫条件下,Ca2+对牡丹抗性生理的调控机制是一个内部复杂的多系统协同代谢过程,尚存在诸多问题有待进一步研究。
参考文献:
[1]乌凤章. 11个北高丛越橘品种对干旱胁迫的响应[J]. 经济林研究,2020,38(1):36-45.
[2]郑秋玲,谭 伟,马 宁,等. 钙对高温下巨峰葡萄叶片光合作用和叶绿素荧光的影响[J]. 中国农业科学,2010,43(9):1963-1968.
[3]安 钰,刘 华,李 明,等. 外源钙对干旱胁迫下甘草生理特性的影响[J]. 中国现代中药,2019,21(10):1397-1401.
[4]李小玲,华智锐,何娇娇. 干旱条件下外源钙对商洛黄芩生理特性的影响[J]. 江西农业学报,2019,31(4):31-35.
[5]赵丽丽,王普昶,陈 超,等. 外源钙对干旱胁迫下岩溶山区白刺花幼苗生长、生理特性的影响[J]. 核农学报,2017,31(10):2039-2046.
[6]陈 莹,王普昶,赵丽丽,等. 外源钙对干旱胁迫下木豆种苗生理特性的影响[J]. 草地学报,2014,22(5):1051-1055.
[7]尹大川,祁金玉,邓 勋,等. 施用外源钙对干旱胁迫下樟子松苗木生理特性的影响[J]. 沈阳农业大学学报,2018,49(5):559-565.
[8]王广恩,金卫平,李俊兰,等. 干旱胁迫下外源钙对棉花幼苗抗旱相关生理指标的影响[J]. 华北农学报,2010,25(增刊2):115-118.
[9]李少朋,毕银丽,陈昢圳,等. 干旱胁迫下外源钙对矿区玉米生长的影响[J]. 中国矿业大学学报,2013,42(3):477-482.
[10]汪娅琴,郭小兰,李培培,等. 4个兔眼蓝莓品种对持续干旱的生理响应及其抗旱性评价[J]. 经济林研究,2021,39(3):186-196.
[11]李林瑜,方紫妍,艾克拜尔·毛拉,等. 自然干旱胁迫对两种小檗幼苗生长和生理生化指标的影响[J]. 北方园艺,2020(4):80-86.
[12]种培芳,单立山,苏世平,等. 甘肃旱区5个经济林树种的苗期抗旱性综合评价[J]. 干旱地区农业研究,2017,35(1):198-204,247.
[13]黄莉娟,赵丽丽,唐华江,等. 不同毛花雀稗种质对干旱胁迫的响应及其抗旱性评价[J]. 西南农业学报,2019,32(11):2557-2563.
[14]李 军,孔祥生,李金航,等. 逐渐干旱对牡丹生理指标的影响[J]. 北方园艺,2014(16):50-53.
[15]李合生. 植物生理生化实验原理和技术[M]. 3版. 北京:高等教育出版社,2000.
[16]高俊凤. 植物生理学试验技术[M]. 西安:世界图书出版社,2000:159-198.
[17]王自布,潘木引,冉晓燕. PEG-6000模拟干旱胁迫对菊花的生理响应[J]. 贵州师范学院学报,2019,35(3):18-22.
[18]蔡建国,章 毅,孙欧文,等. 绣球抗旱性综合评价及指标体系构建[J]. 应用生态学报,2018,29(10):3175-3182.
[19]彭民贵,张 继,陈学林,等. 聚乙二醇模拟干旱胁迫下紫斑牡丹的抗旱性研究[J]. 西北农林科技大学学报(自然科学版),2014,42(4):179-186.
[20]陈晓玲,余土元,秦华明,等. 钙对铬胁迫下玉米幼苗生长及生理特性的影响[J]. 玉米科学,2009,17(4):74-78.
[21]龚 明,李 英,曹宗巽. 植物体内的钙信使系统[J]. 植物学通报,1990,25(3):19-29.
[22]彭 淼,钟晓红,张 玲,等. 钙对干旱胁迫下草莓SOD、CAT、PPO活性的影响[J]. 长江大学学报(自然科学版)农学卷,2009,6(1):11-14,110-111.
