草鱼敏感的负趋音筛选
2024-05-20邬玉娇李晓兵尼玛旦增刘国勇许家炜柯森繁李卫东李冬晴石小涛
杨 吉, 邬玉娇, 李晓兵, 尼玛旦增, 达 瓦, 张 占,刘国勇, 许家炜, 柯森繁, 李卫东, 李冬晴, 石小涛*
(1. 三峡大学水利与环境学院,湖北 宜昌 443002;2. 三峡大学,湖北省鱼类过坝技术国际科技合作基地,湖北 宜昌 443002;3. 中水北方勘测设计研究有限责任公司,天津 300222;4. 西藏自治区水利电力规划勘测设计研究院,西藏 拉萨 850000;5. 河海大学水利水电学院,江苏 南京 210024)
水利工程(水电站、农田水利工程等)可通过增加水的调度进行发电、作物灌溉以增加社会经济效益,但其对水生态系统[1-2]和淡水鱼类[3]产生了不利影响,大坝的修建会阻碍洄游性鱼类的上溯,引水结构和抽水活动更会造成幼鱼或成鱼的损伤和死亡[4]。为解决该问题,可采取部署物理屏障等措施防止鱼类进入危险区域。虽然物理屏障可以防止一些鱼类被夹带[5],但其成本高昂[6],难以安装和维护[7],因此,作为不对鱼类造成伤害的非结构性措施,声音辅助诱趋鱼措施被广泛应用于水利工程[8]。
水环境中充斥的生物和非生物的声音对鱼类的生存和繁殖至关重要[9-10],其在鱼类之间的交流、交配行为、猎物和捕食者的发现、定向和迁徙及栖息地选择中发挥重要功能[11]。已有研究表明,鱼类对单频音和宽频音具有不同的行为反应。在声压为83 dB (dB 为声压级单位)时,蓝鳍金枪鱼(Thunnus thynnus)对400~500 Hz 的单频音最为敏感[12];270 Hz 振动的板材声对虹鳟(Oncorhynchus mykiss)具有吸引作用[13]。大西洋鲑(Salmo salar)幼鱼对10 Hz 的声音存在逃避行为[14-15]。鲢(Hypophthalmichthys molitrix)通常对宽频音表现出负趋音反应[16]。在渡船声的影响下,黑带娇丽鱼(Amatitlania nigrofasciata)的躲避行为增加,欧洲鲦(Hemiculter leucisculus)和三刺鱼(Gasterosteus aculeatus)都有明显的逃避行为[17-18]。这些现象表明鱼类能够对各种声音做出反应且可以区分不同大小或频率的声音,这种检测声音并确定声源方向的能力被称为声源定位[19]。在其他感官无法有效运作的条件下,声源定位可以为鱼类寻找猎物、避开捕食者和危险水域提供重要的空间线索[20]。因此,找出不同鱼类敏感的负趋音作为声学屏障以减少鱼类可能受到的危害至关重要。
目前,国外已建过鱼设施大部分是以鲑科(Salmonidae)鱼类为主要过鱼对象[21],而中国过鱼设施的主要过鱼对象大部分是鲤科(Cyprinidae)鱼类[22],通过聚焦研究鲤科鱼类可解决我国大量过鱼设施对鱼类基础数据的共性需求[23]。本研究结合前人的经验,采用单频音和宽频音两种形式对草鱼进行实验,通过草鱼幼鱼应对水下不同声音的行为反应,判断并筛选其敏感的负趋音,为过鱼设施中辅助诱驱鱼手段及水利结构中避免鱼类的夹带提供参考依据。
1 材料与方法
1.1 实验装置与材料
实验水槽参照Vetter 等[24]的方法,采用尺寸为9.0 m×0.6 m×1.0 m 的狭长混凝土水槽,水槽底部及侧面贴上边长为0.6 m 的白色瓷砖,以便观察实验鱼在水槽中的位置。在水下喇叭的前方距离水槽两端1.2 m 的位置放置拦网,防止实验鱼游入喇叭后方,实验区域实长6.6 m (图1)。实验过程中,槽内水温为20~25 °C,水深保持在0.5 m。水槽上方架设两台摄像机(红外网络摄像头,焦距4 mm,帧率24 Hz,海康威视),每次实验开始前,进行视频录制,以记录实验鱼在声音播放时的行为反应。

图1 实验水槽1.水槽,2.隔网,3.水下扬声器,4.摄像头,5.支撑杆。Fig. 1 Experimental flume 1. flume, 2. netting, 3. underwater loudspeaker, 4. camera, 5. supporting rod.
实验材料为草鱼(Ctenopharyngodon idella),属鲤形目(Cypriniformes)鲤科雅罗鱼亚科(Leuciscinae)草鱼属(Ctenopharyngodon),鲤形目鱼类除了具有耳、鱼鳔、侧线等器官外,还具有韦伯氏器[25],使得鲤形目鱼类的听觉系统更为完善,听力更加敏感、范围更广,且草鱼为中国主要淡水鱼类养殖对象,分布于中国各大水系,故使用草鱼作为实验用鱼。草鱼取自当阳市草鱼养殖场,体长为7~15 cm。实验前,将购得的草鱼放在直径为2 m 的钢化玻璃缸中,进行48 h 的饥饿暂养,全天不间断充氧,选择体表无伤,活性良好的鱼进行实验。
本实验所有操作严格遵守实验动物福利伦理和动物实验安全审查规范,并按照水产动物实验伦理审查委员会制定的规章制度执行。
1.2 实验方法
实验地点位于湖北宜昌,于4—5 月进行,采用单频音和宽频音这两种不同的音频对草鱼进行趋音性实验,以获得更准确、可使结论推导范围更广的结果。
实验中单频音实验共用鱼30 尾,宽频音实验共用鱼30 尾。每次实验观察单尾鱼的行为反应,单频音和宽频音实验各重复30 组,共60 组。每组实验10 min,在每次实验开始前记录实验鱼在水槽中自由游泳10 min 的视频作为空白对照。
鱼类趋音行为的定义 正趋音反应:当播放声音开始15 s 内游向声源,并且从远离声源的一端在30 s 内鱼游过水槽中线(5 m 处),记为1次反应。负趋音反应:当播放声音开始15 s 内游离声源,并且从靠近声源的一端在30 s 内鱼游过水槽中线(5 m 处),记为1 次反应。中性反应:既不符合 “正趋音反应” ,又不符合 “负趋音反应” 。连续反应:如果鱼有 “负趋音反应” ,就继续进行声音实验。当鱼游过中线时,一端停止播放声音,另外一端开始播放声音。这样来回交替更换声源位置,记录鱼来回游动次数。如果鱼有2 次或更多次趋音反应即有 “连续反应” 。
游泳速度的定义 实验鱼在1 次经过水槽中线后并在30 s 内再次经过水槽中线,这种躲避声音的行为记为一次反应。在这2 次经过水槽中线的时间内实验鱼的平均速度为反应速度;平均反应速度为所有反应速度的平均值;实验鱼的最大反应速度为实验鱼的反应速度最大值;总平均速度为在实验时间10 min 内实验鱼的平均速度。
声音的采集 单频音的合成:使用Cool editor pro 软 件 合 成500、1 000、1 500、2 000、2 500 和3 000 Hz 的单频音。宽频音的录制:扬子鳄(Alligator sinensis) 吼叫声在安徽鳄鱼繁殖研究中心录制,用竹竿绑住水听器放于离扬子鳄1.5 m的位置,待其吼叫时进行录制,再选取其中有效的声音部分用Cool editor pro 软件进行处理,得到一段4.8 s 的音频。
单频音实验 实验开始前让鱼在实验水槽中适应2 h 左右。开始实验时,观察鱼的位置,当鱼靠近一端的水下喇叭时播放一段30 s 的单频音(500、1 000、1 500、2 000、2 500 和3 000 Hz),若实验鱼在15 s 内游离声源并在30 s 内游过水槽中线(5 m),则记为1 次反应,并关掉正在播放的水下喇叭,打开另一端的水下喇叭继续播放声音,若鱼继续反应,则如此反复交替更换声源,直到实验结束,单次实验时间为10 min。
为尽量减少之前实验音对鱼的影响,待实验鱼休息15 min 后,再播放另一种单频音,直到6种单频音都播放完毕。单频音实验共用草鱼30 尾。待实验完成后,将实验鱼保存好,待进行下一步宽频音实验。
宽频音实验 实验以扬子鳄吼叫声作为声音刺激源,使用进行过单频音实验的鱼作为实验鱼,让鱼休息15 min 后开始播放扬子鳄吼叫声,若实验鱼有反应则来回更换声源,直到实验结束,单次实验时间为10 min。
在实验开始前,先让实验鱼在水槽中适应2 h左右。开始实验时,观察鱼的位置,当鱼靠近一端的水下喇叭时播放扬子鳄吼叫声,若鱼有反应,则继续交替放音,直到实验结束。宽频音实验共用草鱼30 尾。6 种单频音加上空白对照组和1 种宽频音,共8 类实验(n=8)。
1.3 数据分析
水下喇叭附近声场的表征 将声级计与水听计相连,测量水下喇叭播放声音后的声音强度。测量时,单频音及扬子鳄吼叫声的声强值都取5 s 内的平均值,然后用Origin 8.1 软件画出测出的声场图,且只测量实验区域的声场,即两个拦网之间的区域(长6.6 m,宽0.6 m)。将水槽划分为0.1 m×0.4 m 的网格,在靠近水槽边壁的位置划分为0.05 m×0.40 m 的网格进行测量,共测量153 个点,测量水深为22 cm。测量所有播放的单频音及宽频音,用Origin 8.1 软件画出测量的声场图。
声音的反映 通过测量均方根电压(rms)并使用Cool Edit Pro 2.1 软件转换为以dB 为单位的相对声压水平,计算每个频率的相对声压级(SPL)。采用2 048 点快速傅里叶变换(汉宁窗)和采样率为44.1 kHz 的方法计算了声波的频率和功率谱,用Origin 8.1 软件画出声强频率图。
数据分析方法 使用跟踪软件Logger Pro 3.8.5.1 软件对录像进行分析。Logger Pro 打点的时间间隔为1 s,每个视频共打点600 个,以确定每个时刻实验鱼的位置。以每条鱼作为观察对象,对游泳速度和消耗时间进行量化,并统计鱼的反应次数、游过水槽中线的总次数、反应速度、最大反应速度、总平均速度及首次游过水槽中线所需时间,使用单因素方差分析(One Way-ANOVA)分析鱼的反应次数和游泳速度等数据。
2 结果
2.1 声音在水槽中的分布
对播放的6 种单频音和扬子鳄吼叫声进行声场图的绘制,图2-a~f 为播放500~3 000 Hz 的单频音时所绘制的声场图。
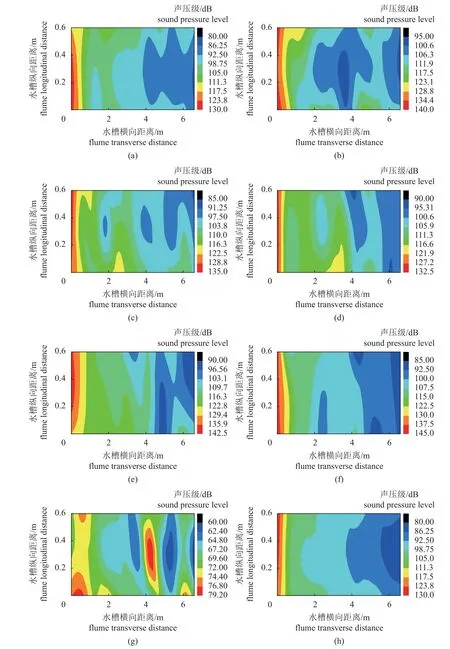
图2 水槽中的声场分布图(a) 500 Hz,(b) 1 000 Hz,(c) 1 500 Hz,(d) 2 000 Hz,(e) 2 500 Hz,(f) 3 000 Hz,(g) 背景声场,(h) 扬子鳄吼叫声,图6 同。Fig. 2 Sound field distribution in flume(a) 5 00 Hz, (b) 1 000 Hz, (c) 1 500 Hz, (d) 2 000 Hz, (e) 2 500 Hz, (f) 3 000 Hz, (g) background sound field, (h) A. sinensis roar, the same as Fig.6.
播放单频音时,最大声强在水槽中喇叭附近,为130~145 dB;最低声强在水槽末端,为80~95 dB。在整个水槽中声强有一定的衰减,最大衰减幅度为60 dB,最小衰减幅度为42.5 dB。图g 为水槽中的背景噪音,最大约为76.8 dB,最小约为60 dB。图h 为播放扬子鳄吼叫声时所绘制的声场图,播放时,水槽中的最大声强为130 dB,最小声强为80 dB,在水槽中的衰减幅度约为50 dB。
2.2 部分声音的声强和频率
图3 中除了反映1 500 Hz 单频音和扬子鳄吼叫声外,还对水下喇叭播放出的扬子鳄吼叫声进行了描述。从图中可以看出,空气中的扬子鳄吼叫声与水下录制的扬子鳄吼叫声频率无太大差别,其峰值集中在100~500 Hz。
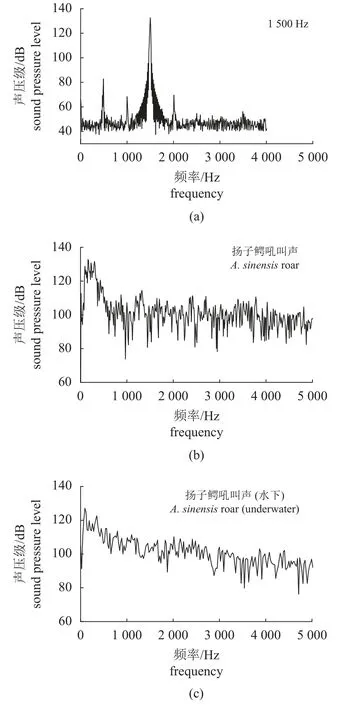
图3 几种典型的声强频率图(a) 1 500 Hz;(b) 扬子鳄吼叫声;(c) 扬子鳄吼叫声(水下)。Fig. 3 Several typical sound intensity frequency diagrams(a) 1 500 Hz; (b) A. sinensis roar; (c) A. sinensis roar (underwater).
2.3 草鱼的反应
实验鱼的反应次数如图4 所示。从图中可以看出,在空白对照组中,实验鱼的反应次数为(0.9±0.3)次,而在播放单频音时,实验鱼的反应次数为(0.4~1.7)次。其中在播放500 Hz 时,实验鱼的平均反应次数最大,为(1.7±0.6)次,而播放1 500 Hz 的单频音时,实验鱼的反应次数最少,为(0.4±0.3)次。与单频音和空白对照组相比,在播放扬子鳄吼叫声时,实验鱼的平均反应次数显著增加,高达(5.0±0.9)次(F=8.158,P<0.000 1)。
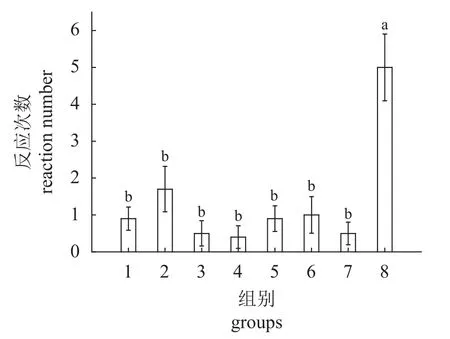
图4 不同声音下草鱼的反应次数(1) 空白对照;(2) 500 Hz;(3) 1 000 Hz;(4) 1 500 Hz;(5) 2 000 Hz;(6) 2 500 Hz;(7) 3 000 Hz;(8) 扬子鳄吼叫声。 图中不同字母表示有显著差异(P < 0.05),图5、图7、图8 同。Fig. 4 Times of response of C. idella under different sounds(1) blank control; (2) 500 Hz; (3) 1 000 Hz; (4) 1 500 Hz; (5) 2 000 Hz;(6) 2 500 Hz; (7) 3 000 Hz; (8) A. sinensis roar. There were significant differences between different letters in the figure (P < 0.05 ), Fig. 5, Fig. 7,Fig. 8 are the same.
实验鱼游过水槽中线的总次数如图5 所示。在实验时间10 min 内,实验鱼反应的总次数为实验鱼游过水槽中线次数的总和。该图中,空白对照组反映了实验鱼的活跃程度。从图中可以看出,在进行1 500、3 000 Hz 的单频音时,实验鱼在水槽中来回游动的次数略低于空白对照组,但是无显著差异。其他单频音实验时,实验鱼来回游动的次数无明显增加。该现象证明单频音对实验鱼在水槽中的行为表现无显著影响。但是在播放扬子鳄吼叫声时,实验鱼在水槽中来回游动的次数明显增加(F=14.319,P<0.000 1)。该现象表明宽频音显著影响了实验鱼的行为。

图5 不同声音下草鱼游过水槽中线的总次数Fig. 5 Total number of C. idella swimming through the middle line of the flume under different sounds
通过对比图4 和图5,在播放宽频音时,实验鱼的反应次数和反应总次数都显著高于空白对照组和单频音实验组。虽然在播放宽频音时实验鱼的反应总次数(均值为10.9±0.9)并不完全为反应次数(均值为5.0±0.9),但是由于宽频音的作用,引起了实验鱼逃避声源的反应,故相较于单频音,宽频音更具有驱鱼效果。
2.4 草鱼的代表性反应
在实验中,8 尾实验鱼的代表性反应如图6所示。在不播放声音时,实验鱼在水槽中自由游动或待在拦网附近。与空白对照组相比,在进行单频音实验时,实验鱼在水槽中的游泳行为无明显不同,但是在进行宽频音实验时,在播放声音后,实验鱼就立刻表现出了强烈的逃离声源的行为反应。并在贴网一段时间后继续,在水槽中线附近来回游动。

图6 草鱼分别对单频音及宽频音的代表性反应曲线图中 “×” 代表实验鱼的负趋音反应。Fig. 6 Representative response curves of C. idella to single tone and broadband tone' × ' represents negative phonotaxis of the experimental fish.
2.5 实验鱼的游泳速度
在播放扬子鳄吼叫声时,实验鱼的平均反应速度均值为0.20 m/s,而空白对照组为0.14 m/s;在播放500~3 000 Hz 的单频音时,实验鱼的平均反应速度为(0.11~0.17 m/s)。可以看出在播放单频音时,实验鱼的平均反应速度与空白对照组差异不显著,而在播放扬子鳄吼叫声时,实验鱼的平均反应速度虽高于单频音与空白对照组,却无显著差异(F=1.156,P=0.348)。在不同声音下实验鱼的最大反应速度范围为(0.12~0.29 m/s),其中在播放扬子鳄吼叫声时,实验鱼的最大反应速度最大。在播放3 000 Hz 的单频音时实验鱼的最大反应速度最小,这与实验鱼的平均反应速度结果一致。
虽然在播放扬子鳄吼叫声时,实验鱼的最大反应速度略高于空白对照组与单频音实验组,但是无显著差异(F=1.232,P=0.308)。从图7 中可以看出,在播放扬子鳄吼叫声时,实验鱼的总平均速度为(0.10±0.01 m/s),与空白对照组(0.06±0.01 m/s)、500 Hz (0.06±0.01 m/s)、1 000 Hz (0.05±0.01 m/s)及2 000 Hz (0.05±0.02 m/s)的单频音实验组相比无显著差异,但与1 500 Hz (0.03±0.01 m/s)、2 500 Hz (0.030±0.005 m/s)及3 000 Hz (0.04±0.01 m/s)的单频音实验组相比具有显著差异(F=6.372,P<0.000 1)。

图7 草鱼的总平均速度Fig. 7 Total average speed of C. idella
通过比较实验鱼对不同声音的平均反应速度、最大反应速度及总平均速度可知,在空白对照组中或在播放声音后,实验鱼能在30 s 内2 次经过水槽中线,所产生的反应速度相差不大。
2.6 实验鱼首次游过水槽中线所需时间
实验开始时,首次播放声音后,实验鱼游过水槽中线所需时间如图8 所示。从图中可以看出,播放扬子鳄吼叫声时,实验鱼首次游过水槽中线时间最短,虽与空白对照组、500、1 000 及2 500 Hz单频音实验组无显著差异。但与1 500、2 000 及3 000 Hz 单 频 音 实 验 组 差 异 显 著(F=6.446,P<0.000 1)。在进行3 000 Hz 的单频音实验时,实验鱼首次游过水槽中线所需时间最长,其次是1 500及2 500 Hz 的单频音实验。
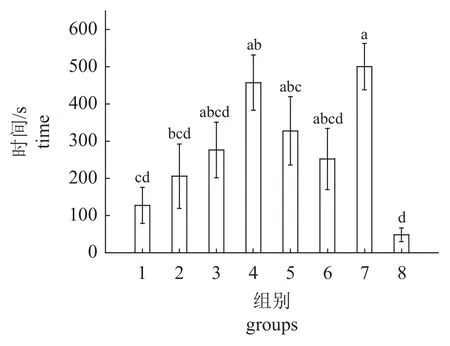
图8 草鱼首次游过水槽中线所需时间Fig. 8 Time required for C. idella to swim across the middle line of the flume for the first time
3 讨论
鱼类对声音的趋性按其行为响应特征可分为正趋音性和负趋音性,正趋音性即声源对鱼类产生吸引效果,鱼类会因正趋音的存在靠近声源;而负趋音相反,声源对鱼类产生驱离效果,鱼类会因负趋音的存在远离声源。
水温变化会对鱼类的生理、分布和生存产生影响,升高的水温会提高新陈代谢速率,并迫使鱼类迁移到较冷的水域[26]。鱼类作为变温动物,依靠行为策略来调节体温。不同种类的鱼通过热导航,根据所经历的环境温度来调整行为策略,从而找到它们适宜的最佳温度[27]。草鱼对水温适应性较强,适宜温度在20~32 °C,覆盖了4—5 月的昼夜温差变化。本实验过程中水温维持在20~25 °C,在草鱼适宜温度范围内,可忽略水温对草鱼行为反应的影响。
实验通过对比草鱼在不同声音下的表现,找出了草鱼敏感的负趋音,即扬子鳄吼叫声。在前人的研究中,Vetter 等[24]以鳙(Aristichys nobilis)为对象进行鱼类趋音性研究,并通过交替改变声源位置的方法,观察鳙对单频音(500~2 000 Hz)和宽频音(0.06~10.00 kHz)的行为反应,结果表明鳙对船声(宽频音)更为敏感,表现出负趋音性,而对单频音的反应不强烈。刘国勇等[28]使用水下扬声器播放扬子鳄嘶嘶声(0.05~5.00 kHz)录音的宽频音,评估裸腹叶须鱼(Ptychobarbus kaznakovi)的反应次数和横向游泳速度,研究结果表明,扬子鳄嘶嘶声是驱除裸腹叶须鱼的有效威慑声音。这与实验的结果一致,该结果表明部分鲤科鱼类对宽频音的反应更为强烈。
鱼类对单频音的反应远不如对宽频音的反应强烈,产生此现象的原因可能是听力阈值的存在[29]。鱼类的可听频率维持在固定区间内,不同鱼类的听力阈值有所不同。声强超过鱼类的听力范围会严重损伤鱼类的听觉系统,造成不可逆的伤害[30]。实验的用音都在草鱼的听力范围内,不会对其造成损伤。有研究表明,大多数鱼类能听到的声音频率范围为50~1 000 Hz,少数鱼类能听到大于3 kHz 频率的声音,仅有极少数鱼类能够听 到 大 于100 kHz 频 率 的 声 音[31-32]。Ladich 等[33]对51 科111 种鱼类的基线听力能力进行了描述和比较,发现多数鲤科鱼类对300~3 000 Hz 频率比较敏感。Vetter 等[24]的一项研究表明,一些鲤科鱼类的听力阈值可能会高于3 000 Hz,其中鲢和鳙可以检测到0.1~5.0 kHz 的宽范围频率,它们的最低平均听力阈值为500 Hz[34]。本研究的扬子鳄吼叫声宽频音频段范围为0~5 000 Hz,在100~500 Hz 达到波峰,并在5 000 Hz 内维持在较高水平,这与Vetter 等[24]的研究结果一致。
另外,实验选取扬子鳄吼叫声作为宽频音,而草鱼对此表现出负趋音性,其可能的原因是因捕食胁迫的存在。在遭遇捕食者时,由于视觉、声音、气味等对鱼类感官的影响,鱼类会表现出反捕食行为以应对当前的威胁[35-37]。有研究表明,在实际捕食胁迫下,草鱼会在具备较高游泳性能的基础上缩短反应时滞以保证其能有效躲避捕食者,会基于自身游泳运动能力及当前环境中的捕食风险水平优化其逃逸反应[37]。朱存良[38]提出鱼类会对其同类的负伤声和逃避声、外敌的摄食声和游泳声以及渔船、渔具发出的异常声等呈负趋音性,鳄鱼作为肉食性动物,因此草鱼幼鱼会对扬子鳄吼叫声产生逃离反应。
在实际环境中,水下声音复杂多变,在远岸的水体中,由于风浪和底质的影响,会有声场波动,鱼类会因为这种短暂声场变化而产生反应,但不会对鱼类的生理产生影响。在近岸的水体中,由于人类活动的影响,会产生非自然噪声,例如近岸处的城市噪声,码头旁的船舶噪声等,这类声音频率过大且长期存在,会给生活在周边的鱼类产生不可逆的影响,比如反应迟钝、听觉系统损伤等[39]。为解决该问题,可参考本研究结论,在岸边布置扬声器,使用不同鱼类的敏感负趋音控制指定鱼种的离岸距离,保护鱼类。
研究表明,可利用宽频音建立声学屏障[40],防止鱼类受到引水结构的夹带等意外伤害,为鱼类保护提供新思路。当然,研究存在一定的不足。由于实验场地的限制,水槽的宽度有限,在播放声音后,声音在水槽中有反射,导致声场分布杂乱,对实验鱼的行为会有一定的影响[24],如果在池塘或天然河流等地进行实验,会消除这部分的误差。另外,在此次实验中,只记录了实验鱼反复越过水槽中线的次数,在实际环境中,鱼类的逃窜路径十分复杂,逃窜路线的复杂程度是摆脱危险的关键,如果能全方位地观察鱼类的游泳形态(加速度、转弯率等)[41],研究将更深入。
(作者声明本文无实际或潜在的利益冲突)
